
鄂尔多斯高原灌丛化草地恢复机制解析
2022-06-09 07:38郑元润来利明蔡文涛杨柳柳鑫
山西大学学报(自然科学版) 2022年3期
郑元润,来利明,蔡文涛,杨柳,柳鑫
(中国科学院 植物研究所,北京 100093)
0 引言
草地灌丛化是全球干旱、半干旱区草地退化的普遍现象[1-2],全球超过20亿人口受到草地灌丛化的影响[3],因而受到科学界的高度重视。由于灌丛可以阻截风沙带来的细粒物质,经过多年的风蚀与风积作用,原来的草地会形成许多高低、大小不等的以灌木为中心的“肥岛(fertilizer island)”[4-5]。沙基质的灌丛化草地进一步退化后则向流动-半流动沙地演化,最终甚至成为沙漠[6]。人类干扰是灌木入侵的主要驱动因素[7-8],全球变化加快了灌木入侵的发生[9]。草本与灌木植物对水分与营养物质利用方式的不同[10],以及土壤水分可利用性的变化[7]是决定灌木入侵的重要生态因子。草地灌丛化改变了植物群落的物种组成,生态系统的功能亦发生显著变化,尤其是土壤质地与土壤水分平衡过程变化更为明显[7-13]。
草地灌丛化的一些生态学过程已在美国西南部退化草地中得到较好研究,如Chihuahuan沙地[14-16]、Mojave沙地[17]及 Sonoran 沙地[18]等。草地灌丛化驱动因素的作用机制已得到较好解释,提出了控制草地灌丛化发展的初步方法,如Abernathy去除灌木与加播草本植物的控制实验[19],以及火烧在控制草地灌丛化中的作用等[20]。研究表明仅仅停止放牧,严重灌丛化后的草地难以自然恢复,去除灌木能够促进灌丛化草地恢复,去除灌木与加播草本植物相结合效果更佳[19],即使如此,草地仅得到局部恢复,总体上仍以灌木占优势[21]。目前已有多种假说解释草地灌丛化的机制。自下而上假说与水分及养分等资源效应有关,自上而下假说与火及动物采食有关[22-23]。生态位假说是自下而上的假说之一,认为水分是干旱、半干旱区植物群落动态的主要驱动因子,灌木和草本植物表现出不同的水分生态位,深根系灌木主要利用深层土壤水分,浅根系草本植物主要利用浅层土壤水分[22-24]。双层假说认为深层土壤水分有利于灌木植物生长,浅层土壤水分有利于草本植物生长[22]。资源库假说认为浅层土壤水分利于所有植物生长,深层土壤水分仅利于灌木植物生长[2]。由于群落生态过程的复杂性,目前仍缺乏统一的理论解释草地灌丛化过程,灌丛化草地能否恢复仍是争议性较大的问题。
我国干旱、半干旱区草地亦发生了灌丛化,如在锡林郭勒草原[25]、贺兰山西坡草地[26]、鄂尔多斯高原等[6],对草地灌丛化生态过程研究逐步深入。郑敬刚等对灌丛化草地土壤异质性的研究发现灌丛化导致土壤水分、养分空间分布发生改变,导致草地结构和功能的变化[26]。韩砚君等采用卫星遥感数据对锡林河流域近30年的植被变化进行了研究,结果表明草地灌丛化过程中群落结构发生了明显变化,草地灌丛化趋势明显加剧,景观格局变化显著,破碎化程度增加[27]。彭海英等认为灌丛化影响草地生态系统中水分的再分配和利用格局[28]。熊小刚和韩兴国发现小叶锦鸡儿(Caragana mi⁃crophylla)灌丛扩张改变了土壤碳、氮资源的空间分布格局,影响了生态系统的稳定性[25]。尽管目前已从自然与人为因素综合作用角度对草地灌丛化的成因进行了分析,对草地灌丛化的过程和机制有了较为深入的认识,但这些研究结果多基于对不同区域的分散研究,针对同一生态系统的长期、多维度的综合分析较少,制约了对草地灌丛化发生过程和机制的系统认识,进而导致灌丛化草地能否恢复,在哪种条件下可以恢复的全球性认识不一致。因而,选择代表性灌丛化草地,开展长期系统研究十分必要。
鄂尔多斯高原位于内蒙古自治区南部,西、北、东三面为黄河环抱,为干旱、半干旱区的典型过渡地带,生态环境脆弱敏感。鄂尔多斯高原属于古老沉积构造盆地,东部为准格尔黄土丘陵,西部为桌子山低山缓坡和鄂尔多斯高地,北部为库布其沙漠,南部为沙漠、滩地、湖泊相间的毛乌素沙地,中部为波状起伏的高原,地表风沙广布,东部为干草原栗钙土,西部为荒漠草原棕钙土[29]。历史上鄂尔多斯高原是水草丰美的草原,地带性植被为暖温带长芒草(Stipa bungeana)草原,但目前鄂尔多斯高原的优势植物为半灌木油蒿(Artemisia ordosica),发生严重草地灌丛化。由于鄂尔多斯高原为我国的荒漠化中心之一[30],亦为黄河粗泥沙的主要来源;同时,鄂尔多斯高原也是我国重要的能源基地和华北生态安全的重要屏障。因此,研究鄂尔多斯高原草地灌丛化过程,分析草地灌丛化的驱动因素,解决灌丛化草地能否恢复的争论性问题,对深化和发展草地灌丛化理论,促进灌丛化草地恢复和可持续利用,保障我国能源和生态安全具有重要意义。
本文为研究团队20多年来关于草地灌丛化研究的系统总结,针对鄂尔多斯高原草地灌丛化的主要驱动因素—过度放牧,我们分析了禁牧后灌丛化草地的植被演替,从放牧干扰角度揭示了灌丛化草地的恢复过程;研究了天然灌草植物群落过渡过程中土壤和植物群落的变化特征,从群落自然分布的角度阐明了灌丛化草地的恢复过程。在此基础上,从灌草植物抗氧化酶和渗透调节物质及灌草植物水分利用来源角度阐明了灌丛化草地的恢复机制,提升了对干旱、半干旱区草地灌丛化过程和机制的科学认识,对我国乃至全球类似区域灌丛化草地的管理提供了重要的理论和实验支持。
1 研究区概况
研究地点位于内蒙古自治区伊金霍洛旗(39°21′N,109°51′E),该旗位于鄂尔多斯高原东南部,毛乌素沙地边缘,东与准格尔旗相邻,西与乌审旗接壤,南与陕西省榆林市神木县交界。本区属于暖温带半干旱气候,年均降水345.2 mm,年均温6.7°C。生长季为4月至10月。地带性植被为暖温带长芒草草原,目前仅零星分布在梁地的上部,半灌木油蒿群落是风沙土上发育最普遍的植被类型。除分布在梁地上部的地带性栗钙土和滩地上的沙地草甸土外,绝大部分地区土壤为风沙土[6]。
2 研究方法
为理解停止放牧等人为干扰对油蒿灌丛草地动态的影响,阐明表层土壤机械组成、不同土层土壤水分、根系生物量、地上生物量及物种组成的变化规律,澄清油蒿灌丛草地在停止人为干扰后能否恢复的问题,于1992年在伊金霍洛旗分布的油蒿群落设立6个50 m×50 m的样地,选择其中3个样地进行围封。之后分别于1992、1996、1999、2002、2005、2008、2011在每个样地内设立1个2 m×2 m的样方,调查植物种数,个体数量,地上部分生物量。采用烘干法测定 5、10、20、30、40、60、80、100 cm 土壤水分。采用比重计法测定0~10 cm土壤机械组成。采用挖掘法每隔10 cm测定分层根系生物量[6]。
在伊金霍洛旗的较小范围内存在长芒草群落向油蒿群落逐渐过渡的自然分布区,形成长芒草群落、长芒草+糙隐子草(Cleistogenes squarrosa)群落、长芒草+油蒿群落、油蒿+糙隐子草群落、油蒿+达乌里胡枝子(Lespedeza davurica)群落、油蒿群落等逐渐过渡的群落,这为研究草地灌丛化的过程提供了天然场所。为从自然分布的由长芒草向油蒿过渡系列群落的角度阐明灌丛化草地发生或恢复过程,分析植物群落特征、土壤物理性质变化与草地灌丛化的关系,于2015年8月植物生长旺季,在长芒草群落向油蒿群落过渡区域的6个典型群落中,设置6个样地,每个样地包括3个重复。在每个重复样地中设置样方,样方大小为灌木5 m×5 m,草本植物1 m×1 m。记录每一个样方的经纬度、海拔、群落种类组成、植株高度、个体数量等指标。采用刈割法收割地上植株,测定每种植物的生物量[31]。采用根钻法测定植物根系生物量[32]。用环刀法在每个样方内采集不同土层土样,取样深度为0~50 cm。采用激光粒度仪测定土壤机械组成,采用烘干法测定土壤水分和容重[32]。
由过度放牧造成的不同程度的灌丛化草地系列,包括流动沙地沙蓬(Agriophyllum squarro⁃sum)群落、半固定沙地圆头蒿(Artemisia sphaerocephala)群落、固定沙地油蒿群落、固定沙地长芒草群落,可作为灌丛化草地恢复系列。为理解不同灌丛化阶段优势植物生理生化特征在植物适应相应环境,进而影响植物群落演替的机制,以及不同灌丛化阶段主要灌草植物水分利用来源分异与土壤水分可利用性关系及其对灌丛草地动态的驱动作用,于2017年7月,在每个系列的灌丛化草地设置3个重复样地。在每个重复样地中,设置一个20 m×20 m的样方,采集植物叶片,测定叶片抗氧化酶活性(超氧化物歧化酶、过氧化物酶、过氧化氢酶)、丙二醛和渗透调节物质含量(脯氨酸、可溶性糖和可溶性蛋白)[33]。于2019年5月、7月和9月降水过后2 d,在每个样方内同时收集植物、土壤和地下水同位素样品。在流动沙地沙蓬群落和固定沙地油蒿群落样地附近收集井水作为地下水。所有同位素样品收集、封装后带回实验室进行同位素测定。在每个样方内采用环刀法采集不同层次土壤样品,带回实验室测定土壤水分和容重。采用挖掘法测定植物根系生物量[34]。
采用重复度量方差分析方法分析灌丛化草地围封处理、围封时间和(或)土层深度等因子对土壤根系生物量、地上生物量、土壤水分、土壤机械组成的影响;采用方差分析和Tukey多重比较方法分析群落类型、土层深度等因子对地上生物量、根系生物量、土壤水分、土壤机械组成的影响;采用方差分析方法分析灌丛化阶段、植物种类对主要灌草植物生理生化参数的影响;采用重复度量方差分析方法研究土层深度、季节、灌丛化阶段对土壤水分和氧同位素的影响。
3 结果与分析
3.1 油蒿灌丛化草地恢复过程
3.1.1 停止放牧后油蒿灌丛化草地恢复过程证据
围封去除放牧干扰后(1992年围封),经过长期演替,以油蒿为主的灌丛化草地向以长芒草为主的草地恢复的趋势明显。不同土层土壤水分、表层土壤机械组成、不同土层根系生物量、植物群落地上部分生物量长期观察数据显示:围封后油蒿群落的地上生物量由最高的231.7 g/m2下降至9.3 g/m2,基本处于消亡状态,而长芒草生物量则由零上升至137.6 g/m2;0~10 cm根系生物量显著增加[6],10 cm~20 cm根系生物量显著减少,表明针茅的根系在增加,油蒿的根系在减少(P<0.05)(图1);围封后0~10 cm土壤含水量呈显著上升趋势,10 cm以下土壤含水量呈显著下降趋势,尤其是10 cm~20 cm油蒿根系的主要分布区,土壤水分下降更为明显(P<0.05)(图 2);0~10 cm 土层中,粒径<0.01 mm的细粒物质在围封后显著增加。未围封对照区油蒿灌丛化草地上述指标未发生显著变化(P>0.05)(图1和图2),表明灌丛化草地的恢复主要是由于停止人为干扰所致,气候变化的作用不明显[6]。由于围封基本停止了对土壤和植物群落的干扰,同时由于大气降尘及油蒿阻挡风沙的作用,土壤表层细粒物质逐渐增多,进而形成地表结皮。加之半干旱区降水较少,降雨后水分向土壤深层的入渗减少,土壤表层水分得到改善,而深层土壤水分减少。这种变化不利于主要利用深层土壤水分的深根系植物油蒿,而利于主要利用浅层土壤水分的浅根系植物长芒草的群落建成;反之,如人为干扰继续存在,细粒物质不易在土壤表层积累,难以形成地表结皮,有限的降水向深层渗透,且由于沙土表层毛细管作用被破坏,深层水分不能向土壤表层输送,浅根系的长芒草难以定居和建成[6]。这一结果表明灌草植物水分利用差异及土壤结构变化可能是草地灌丛化发生和恢复的关键因素。当油蒿灌丛化草地逐步恢复为长芒草草地时,由于草本植物对表层土壤水分的消耗增大,表层土壤含水量降低,且由于沙土显著中断了土壤毛细管作用,深层土壤水分不易蒸发而逐渐增加。如过度干扰再次发生,这种土壤水分变化会为油蒿的入侵创造条件。由此可见,目前鄂尔多斯高原以油蒿为主的半灌木植物群落处于放牧干扰和气候相互作用下的动态平衡阶段,如停止放牧干扰,以油蒿为主的灌丛化草地可以恢复为以长芒草为主的草地[6]。

图1 油蒿灌丛化草地围封(F)和未围封样地(UF)不同土层根系生物量。图中直线为回归方程线Fig.1 Root biomass in different soil layers in the fenced(F)and unfenced(UF)plots ofArtemisia ordosica shrub encroached grassland.Lines are results of regression analyses
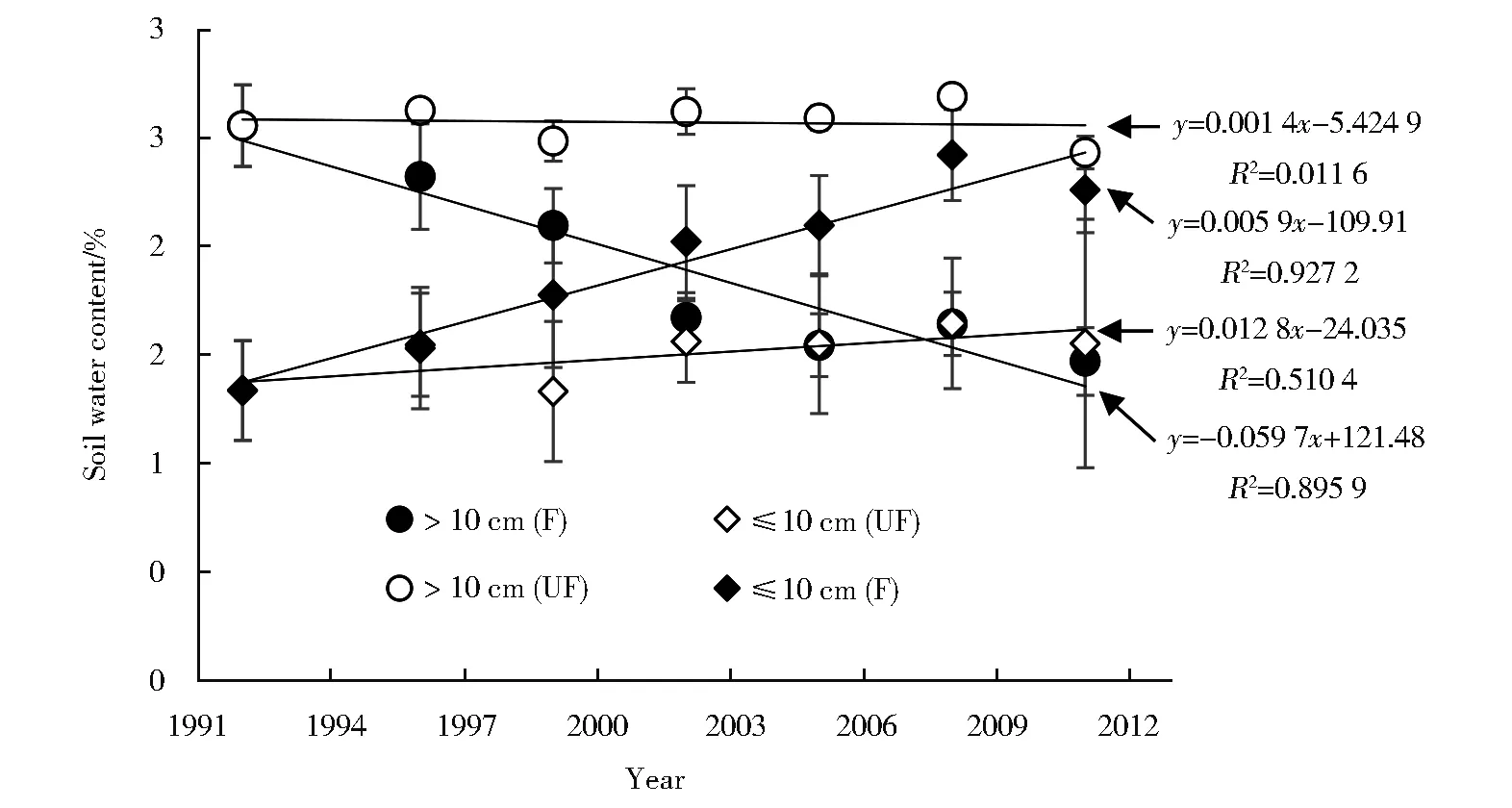
图2 油蒿灌丛化草地围封(F)和未围封样地(UF)不同土层含水量。图中直线为回归方程线Fig.2 Water content in different soil layers in the fenced(F)and unfenced(UF)plots ofArtemisia ordosica shrub encroached grassland.Lines are results of regression analyses
3.1.2 自然分布油蒿群落与长芒草过渡过程证据
从油蒿群落向长芒草群落过渡的过程中,物种数量呈现先增加后减少的趋势,中间阶段物种组成最丰富。随着演替的进行长芒草逐渐增加,油蒿从第五阶段显著减少到最后完消失[31]。从油蒿到长芒草群落的演替过程中,根系分布越来越浅,第六阶段长芒草群落的根系深度达到30 m,第一阶段油蒿群落根系深度可达80 cm~90 cm;土壤由第一阶段的砂土过渡到后5个阶段的砂质壤土,土壤质地类型发生显著变化,第一阶段油蒿群落土壤黏粒含量仅为第六阶段长芒草群落土壤黏粒含量的50%;0~50 cm土层土壤黏粒和粉粒含量总体呈上升趋势,砂粒含量总体呈下降趋势;0~50 cm土层,土壤含水量呈上升趋势,0~10 cm土层土壤含水量逐渐增加[30]。上述结果表明植物土壤相互作用对理解草地灌丛化过程十分关键。土壤表层黏粒物质增加使得降雨分配土壤表层化,有利于浅根系的长芒草群落构建。进一步分析油蒿群落向长芒草群落过渡的土壤结构数据,发现当0~10 cm表层土壤的砂粒含量超过80%时,以长芒草占优势的群落会逐渐演替为以油蒿为主的灌丛化草地,这一数值可作为草地灌丛化发生和恢复的土壤结构阈值,对于制定灌丛化草地恢复对策具有重要意义[32]。
3.2 优势植物抗氧化酶和渗透调节物质调节机制
由于半干旱区日照时数高,地表温度较高,沙土表层干燥;同时,半干旱区降雨较少,沙土渗透性强,降雨易向深层土壤入渗,加之沙土表面土壤毛细管作用弱,深层土壤水分不易蒸发,中深层土壤水分较好,有利于深根系植物生长;黏粒含量较高的土壤水分特性则相反。相应地分布于不同灌丛化阶段的灌草植物为适应土壤水分状况,在生理生态上会表现出不同的适应方式。抗氧化酶和渗透调节物质在植物适应土壤水分变化方面具有较强的指示意义,可用于评估植物对土壤水分的适应状况。不同灌丛化阶段不同土层的土壤水分状况差异较大,可采用抗氧化酶和渗透调节物质分析灌草植物在不同灌丛化阶段替代的机制。
在由过度放牧造成的不同程度的灌丛化草地系列中,流动沙地沙蓬群落中沙蓬的抗氧化酶活性和丙二醛含量显著高于圆头蒿(P<0.05,下同)。半固定沙地油蒿群落中油蒿的超氧化物歧化酶和过氧化氢酶活性高于圆头蒿,而过氧化物酶活性和丙二醛含量小于圆头蒿。固定沙地油蒿群落和固定沙地长芒草群落中长芒草的抗氧化酶活性和丙二醛含量显著高于油蒿。流动沙地沙蓬群落沙蓬的渗透调节物质含量显著低于圆头蒿。半固定沙地油蒿群落中油蒿的渗透调节物质含量显著低于圆头蒿。固定沙地油蒿群落和固定沙地长芒草群落中长芒草可溶性糖含量最高,油蒿最低。由流动沙地沙蓬群落、半固定沙地油蒿群落、固定沙地油蒿群落到固定沙地长芒草群落,群落物种平均抗氧化酶活性先减小再增加,丙二醛含量增加,脯氨酸含量先增加后减小,可溶性蛋白含量先增加后减小。草本植物的酶活性高于灌木。草本植物超氧化物歧化酶、过氧化物酶和过氧化氢酶的平均含量分别是灌木的1.44、6.06和3.26倍。灌木脯氨酸含量是草本植物的2.26倍,草本可溶性糖含量是灌木的1.29倍(图3),灌木可溶性蛋白的含量是草本植物的1.68倍。采用抗氧化酶和渗透调节物质参数,通过主成分分析建立的植物抗旱性指数显示,4种优势植物(沙蓬、圆头蒿、油蒿和长芒草)在干旱胁迫时通过调节抗氧化酶活性和渗透调节物质含量增强抗旱性。从流动沙地到固定沙地,优势植物抗旱性由强到弱为:长芒草>油蒿>沙蓬>圆头蒿,草本植物的抗旱性高于灌木[33]。这也是长芒草能够更好适应半干旱区表层土壤水分变化,逐步代替油蒿的原因之一。
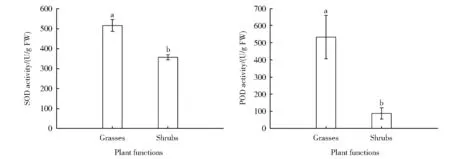
图3 不同功能型植物抗氧化酶活性和渗透调节物质含量(Mean±SE)。不同字母表示差异显著(P<0.05)(修改自Yang等,2019)。SOD、POD和FW分别代表过氧化物歧化酶、过氧化物酶和鲜重Fig.3 Responses of enzyme and osmotic adjustment compounds(Mean±SE)in two plant functions.The different low case letters indicate significant different(P<0.05)(Revised from Yang et al.2019).SOD,POD and FW represent superoxide dismutase,perox‐idase and fresh weight,respectively
3.3 灌草植物水分利用来源分异驱动机制
双层假说和资源库假说均认为灌草植物水分利用来源的分异是驱动草地灌丛化过程的重要因素[2,22]。长期灌丛化草地围封结果也显示油蒿和长芒草水分利用来源分异可能是影响灌草化草地过程的关键因素。但上述认识均缺乏直接证据支持,稳定同位素方法为证实上述假说提供了可能。过度放牧造成的不同程度的灌丛化草地系列中主要灌草植物同位素水分利用来源分析研究结果验证了上述假说与油蒿和长芒草水分利用来源的分异。
流动沙地沙蓬群落沙蓬7月和9月主要利用浅层土壤水(0~20 cm);圆头蒿5月和9月主要利用浅层土壤水,7月主要利用中层土壤水(20 cm~50 cm)。半固定沙地油蒿群落油蒿和圆头蒿9月利用更多的浅层土壤水,7月主要水分来源为中层土壤水。固定沙地油蒿群落油蒿5月和9月主要水分来源为浅层土壤水(28.5%±3.4%和(43.4%±3.0%),7月主要利用深层土壤水(50.2%±16.9%);长芒草在整个生长季均主要利用浅层土壤水((85.8%±5.6%)。固定沙地长芒草群落油蒿5月和9月主要利用中层土壤水(43.6%±8.0%和41.4%±8.5%),油蒿7月主要利用深层土壤水(58.8%±7.8%);长芒草在整个生长季均主要利用浅层土壤水(90.6%±3.6%)。可见浅根系草本植物沙蓬和长芒草在生长季以浅层土壤水分(0~20 cm)为主要水分利用来源;深根系半灌木植物圆头蒿和油蒿会根据土壤水分条件变化调整对各潜在水源的利用比例,水分利用模式灵活,但在固定沙地长芒草群落主要利用中深层土壤水[33]。这表明随灌丛化草地的恢复,半灌木植物与草本植物水分利用来源分异愈加明显,驱动着灌丛化草地的恢复。
3.4 油蒿灌丛化草地管理对策
鄂尔多斯高原草地灌丛化发展时间较长,与国外草地灌丛化形成鲜明区别,如美国西南部草地开发与退化始于19世纪80−90年代,只有100多年的时间,而我国部分草地灌丛化时间达数百年。因此,基于我国典型灌丛化草地的研究有益于更加准确地理解草地灌丛化过程,能够更加有效地指导灌丛化草地的管理。我们的研究结果清楚显示了灌草植物与土壤相互作用是驱动草地灌丛化发生的关键因素。受气候驱动作用和植物群落演化过程中灌草植物与土壤相互作用的共同影响,当停止放牧等人类干扰后,灌丛化草地截留大气降尘,土壤表层结构改善,表层土壤水分增加,草本植物得到恢复。由于灌木逐步退出,深层土壤水分利用减少,随草本植物群落的建成,表层土壤水分减少,深层土壤水分增加,为再次可能的灌丛化创造了条件。当放牧等人类活动加强,草本植物群落逐步受到破坏,得益于增加的深层土壤水分,灌木会再次逐渐入侵。由此可见,适度利用,降低放牧强度是维持灌草植物与土壤平衡关系,进而抑制灌木入侵,维持草地生态系统健康的关键;对于已发生灌丛化的草地,大幅度降低人为干扰,逐步改善表层土壤是灌丛化草地恢复的关键。其次,我们的研究表明位于半干旱区的鄂尔多斯高原及类似地区,在显著降低人类干扰后,灌丛化草地可以恢复为地带性草地,这一结论与国际上严重灌丛化草地不能恢复为地带性草地的结论不同。此外,由于地带性草原中长芒草等草本植物饲用价值高于以油蒿为主的灌丛化草地,应根据灌丛化草地的进程,采取适当措施,促进油蒿灌丛化草地的恢复,这也为灌丛化草地的定向管理提供了科学依据。
4 结论
不同于国际上严重灌丛化草地不能恢复的结论,停止放牧干扰后,鄂尔多斯高原油蒿灌丛化草地自然恢复的趋势十分明显。停止干扰后表层土壤细粒物质增加,半干旱区有限降雨向深层土壤入渗减少,表层土壤含水量增加,深层土壤含水量减少,这种变化有利于浅根系长芒草发育,不利于深根系油蒿发育,符合草地灌丛化过程的“双层”假说。灌丛化草地恢复的土壤质地标准是表层土壤砂粒含量小于80%,黏粒增加,土壤结构逐渐改善。氢氧同位素结果进一步证实灌草植物水分利用来源分异,加之土壤质地变化导致的分层土壤水分变化,共同驱动了油蒿灌丛草地的逐步恢复。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
农业工程学报(2022年10期)2022-08-22
云南大学学报(自然科学版)(2022年4期)2022-08-03
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
江苏农业科学(2020年15期)2020-09-22
安徽农学通报(2020年6期)2020-04-17
语文世界(小学版)(2020年11期)2020-01-08
孩子(2019年8期)2019-08-26
大灰狼(2016年9期)2016-10-13
