
运用RAPD技术分析玉溪市烟草黑胫病菌遗传多样性
2022-06-06 13:54田阳阳王军伟曹洋川杨进飞朱学伟杨毓玲刘迎龙
湖南农业科学 2022年1期
田阳阳,王军伟,胡 阳,曹洋川,杨进飞,朱学伟,杨毓玲,刘迎龙
[1.红塔烟草(集团)有限责任公司,云南 玉溪 653100;2.云南农业大学植物保护学院,云南 昆明 650201;3.玉溪市烟草公司新平县分公司,云南 玉溪 653100]
中国是世界上烟叶生产第一大国,烟草行业在经济生产中具有独特地位,是国家经济的支柱产业[1]。烟草的种植范围极广,从北纬55°到南纬40°之间的南北各地区均有栽培[2]。然而,由烟草疫霉(Phytophthora nicotianaeBreda de Haan)引起的烟草黑胫病是一种毁灭性的土传病害,自1896 年在印度尼西亚的爪哇首次发现后在各地广泛传播,发病率高、分布范围广,造成严重的经济损失[3-5]。烟草黑胫病菌有无性和有性阶段,在侵染植物的过程中,会形成孢子囊、游动孢子、卵孢子和厚垣孢子等多种形态。研究发现,种群大小、基因交流、变异、生殖方式和环境选择作用等诸多因素均能影响微生物群体遗传结构的演化[6]。因此,不同菌株在生物学性状及分子水平上表现出特征性差异,展现出丰富的遗传多样性,但同时也使病害越发严重,难以控制[7]。
近年来,云南省烟叶生产发展迅速,已成为当地农民增收致富的重要途径。该省玉溪市是红塔烟草(集团)有限责任公司的核心采购区域之一,其产量和品质在集团卷烟配方中占有重要位置。然而,随着种植面积扩大,烟区地块难以实现轮作,推广品种抗病性较差,例如KRK26 和红花大金元等品种易感染黑胫病。随着该类感病品种的推广,黑胫病发生面积和严重程度加大。因此,对烟草黑胫病的防治工作一直是烟草生产的重点。病菌群体的地理分布特征既能反映群体的结构动态,也能反映病菌群体的演化规律[8]。为了探究玉溪市烟区烟草黑胫病菌的遗传差异及群体演化规律,研究以玉溪市新平县与澄江市烟区为取样点,通过RAPD 标记技术,从DNA 分子水平探讨云南省玉溪市烟草黑胫病菌群体的遗传多样性,以期为玉溪市烟草黑胫病综合防治提供一定的理论依据与研究基础。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 供试的164 个烟草疫霉菌株分离自云南省玉溪市植烟区土壤及烟草黑胫病标本,在燕麦培养基上活化培养备用,菌株来源及分布见表1。
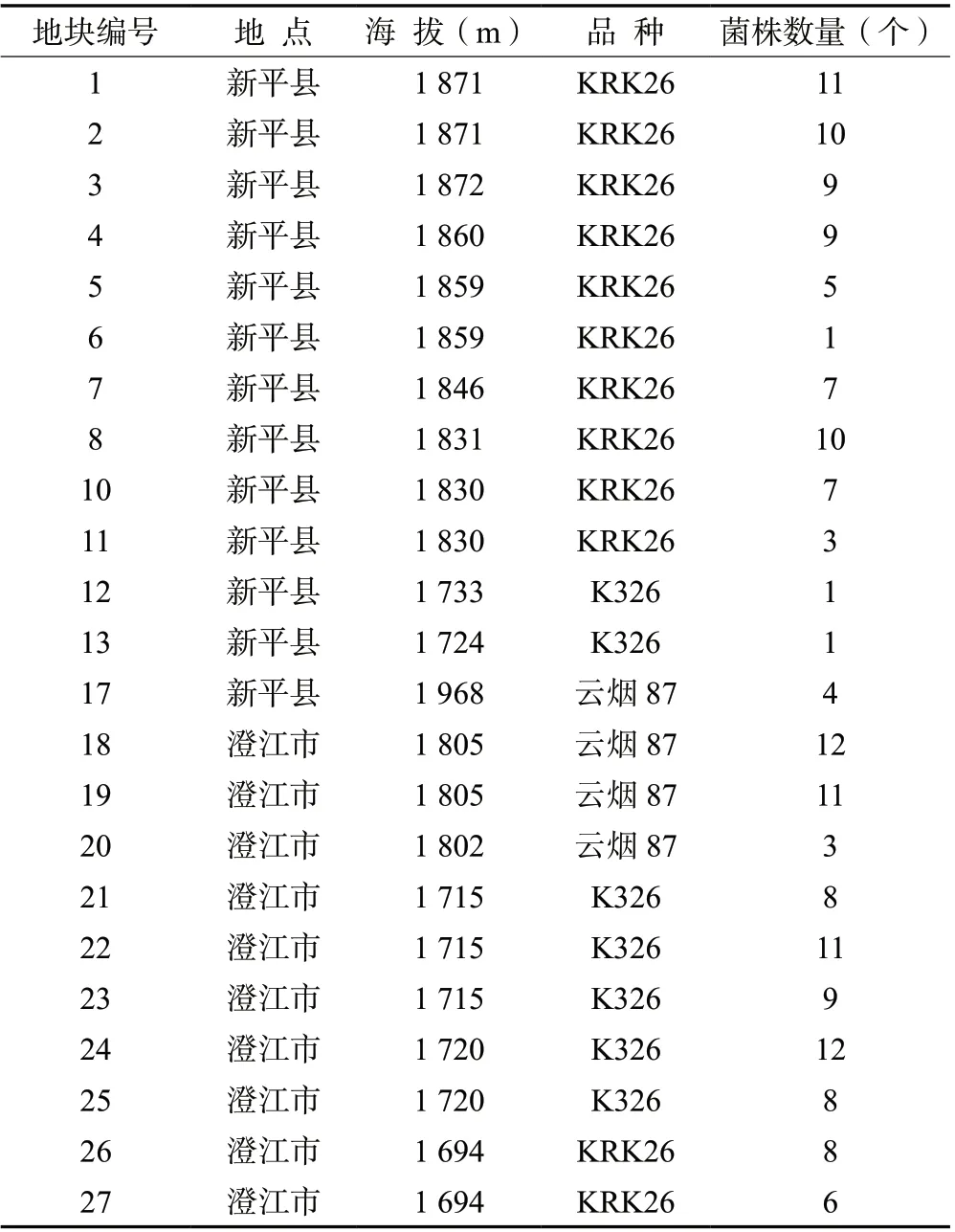
表1 烟草疫霉的地理来源及菌株数量
1.1.2 主要试剂和仪器 RAPD 随机引物由上海捷瑞生物工程有限公司合成;其他试剂如PCR 相关试剂、5 000 DNA Marker、Agarose、CTAB、氯 仿、异戊醇、乙醇、异丙醇等均购自Sigma 公司。主要仪器有:PCR 扩增仪(杭州朗基科学仪器有限公司)、电泳仪(DYY6C,购自北京六一厂);紫外凝胶成像系统(Biotop,220V,上海山富科学仪器有限公司);核酸蛋白定量仪(Nano Drop 2000,美国赛默飞世尔科技公司)。
1.2 试验方法
1.2.1 DNA 的提取 采用CTAB 法提取烟草疫霉DNA[9],用核酸蛋白定量仪(Nano Drop 2000)测定其OD260与OD280的比值。质量较好的DNA 纯度较高,蛋白质含量较少,其OD260/OD280值介于1.8~2.0 之间[10]。将接近于该比值的DNA 样品置于-20℃冰柜内保存备用。
1.2.2 引物筛选 通过查阅相关文献[11-13],从已报道的烟草疫霉RAPD 随机引物和上海捷瑞生物工程有限公司合成的2 000 个寡聚核苷酸随机引物中筛选出扩增谱带多且清楚的6 条随机引物,对供试菌株全基因组DNA 进行PCR 扩增和RAPD 分析。
1.2.3 扩增程序及反应体系 (1)反应体系(20 μL):10×buffer 2 μL,1 U Taq 酶 0.2 μL,0.2 mmol/L dNTP 混合液 1.6 μL,0.5 μmol/L 的RAPD 引物 1 μL,菌丝DNA 模板0.5 μL,最后加ddH2O 补至20 μL,阴性对照为ddH2O。(2)反应条件:95℃预变性2 min;94℃变性1 min,38℃退火1 min,72℃延伸2 min,40 个循环;72℃延伸10 min[12]。取10 μL PCR 产物与3 μL 含核酸染料(花青素)的Loading Buffer 混合均匀后用1.5%琼脂糖凝胶电泳检测,电压设置为50 V,持续2~3 h,结果在紫外凝胶成像系统下观察并采集图像。
1.2.4 扩增产物的分析方法 电泳图谱中的每一条带均为一个分子标记,根据各分子标记的迁移率及其有无转化为二进制数据,具体原则为:在重复扩增中稳定出现的带无论强弱均记为1,无稳定的带记为0。相似系数的计算和聚类分析均由NTSYS 软件完成,并采用UPGMA(Unweighted pair group method with arithmetic mean)构建遗传系统树状图谱[12]。
1.2.5 遗传差异分析 根据遗传系统树状图谱以不同相似水平将供试菌株划分为不同的遗传相似组群,按地理来源、海拔高度、烟草品种等相关因素统计各组群的菌株数量,绘制图表并分析供试菌株的遗传差异。
2 结果与分析
2.1 RAPD 引物筛选结果及多态性分析
根据扩增结果,筛选出6 条多态性较好的引物作为RAPD 扩增引物,详见表2。根据图1 可知,6 条随机引物共扩增出了53 条DNA 条带,平均每条引物扩增出8.83 条DNA 条带,所扩增出的DNA 条带大部分为多态性谱带,多态率为71.70%。不同引物对供试菌株扩增出的DNA 谱带结果也不相同,条带总数在6~10 条。以上结果表明,云南省玉溪市烟草黑胫病病菌存在较丰富的遗传多样性。
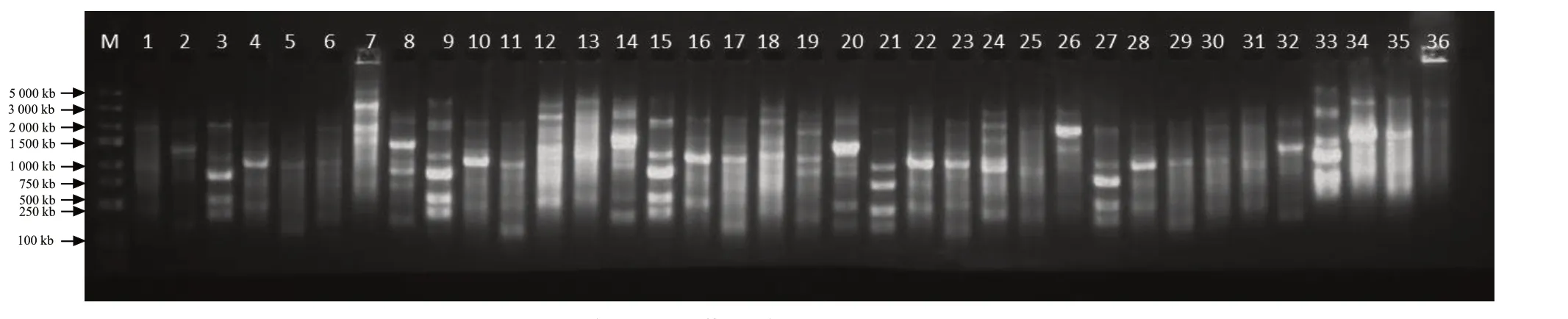
图1 部分供试菌株的RAPD 扩增图谱

表2 6 条RAPD 供试引物扩增谱带数量
2.2 不同烟草疫霉菌株的聚类分析
利用NTSYS 软件以UPGMA 对164 个菌株全基因组DNA 扩增结果进行聚类分析,在0.95 相似水平上每一支各取一株菌株构建遗传系统树状图谱,如图2 所示。根据不同相似水平可划分为不同的遗传相似组,例如在0.74 相似水平上可将164 个菌株划分为4 个组群;在0.84 相似水平上可将其划分为6 个组群;在0.94 相似水平上可将其划分为12 个组群。其中,在0.84 相似水平上划分的6 个组群中,Ⅰ组群的数量最多,为146 株,占89.02%,其他5 组总和仅占10.98%(表3)。
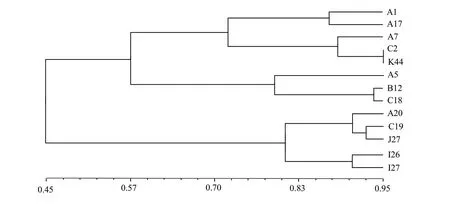
图2 云南省玉溪市烟草疫霉菌株RAPD 聚类分析结果
2.3 烟草疫霉与地理来源、海拔高度、烟草品种关系的遗传差异分析
根据0.84 相似水平将供试的164 个菌株划分为Ⅰ~Ⅵ个组群(表3),按地理来源、海拔高度、烟草品种等相关因素统计不同组群的菌株数量,如图3 所示,大多数烟草疫霉菌株聚集在Ⅰ组群;少量则分散于Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ各组群内。根据地理位置分析,各组群在玉溪市新平县与澄江市境内大致均匀分布;但从新平县和澄江市不同海拔高度(1 694~1 970 m)来看,Ⅰ组群内有较多菌株分布于低海拔地区,其余较高海拔地区无较明显差异;对该地区品种分布与烟草疫霉遗传差异分析可知,烟草品种KRK26 种植区有较多数量的Ⅰ组群烟草疫霉出现,而另外2 个主栽品种则相对较少,但云烟87 也多于K326。

表3 供试菌株在0.84 相似水平上的组群分布及数量

图3 菌株与地理来源、海拔高度、烟草品种关系的遗传差异
3 结论与讨论
3.1 结 论
通过6 条具有多态性的RAPD 引物,随机扩增164 个烟草黑胫病菌菌株的DNA,对供试菌株进行种群遗传差异分析和基因型划分研究,结果显示,在0.84 相似水平上,这些菌株可分为6 个组,其中146个菌株聚集在Ⅰ组,占89.02%,其他5 组总和仅占10.98%。这表明新平县和澄江市的烟草疫霉遗传上具有高度同源性。分析该地区烟草疫霉遗传分化差异发现,其群体基因型划分与地理来源没有直接关系,但与海拔高度、烟草品种分布存在一定关联。该结论对玉溪烟区烟草黑胫病的防控具有重要的指导意义。
3.2 讨 论
RAPD 技术自问世以来,因其成本较低,运用范围广泛而得到快速发展。到目前为止已在物种多态性检测、品种及亲缘关系鉴定、遗传图谱等多个方面得到应用。如钱旎[7]和刘芳等[14]用RAPD 与McRAPD标记技术,分析重庆和河南地区烟草黑胫病菌的生理小种和遗传多样性;刘晓侠等[15]通过RAPD 标记和集团分离分析法(BSA)筛选,找到了与烟草抗黑胫病基因Bs1(t)连锁的RAPD 标记OPB023000 和OPM112500,为该基因的进一步研究利用以及烟草抗病品种的选育积累了基础;Qamer 等[16]基于RAPD标记技术对东南亚地区的蜜蜂、美洲蜜蜂群体遗传差异进行分析,结果表明,它们之间的遗传变异水平较低,但也有较小程度的迁移存在,这一结论为保护该地区特有蜂种的生物多样性提供了指导。
该研究运用RAPD 技术对玉溪烟区的164 株烟草疫霉进行种群遗传差异分析,结果表明,玉溪新平县和澄江市烟区中的烟草疫霉在遗传上具有高度同源性,分化差异较小,遗传变异并不明显,这与付静等[17]的研究结果相似。然而,同一地区也存在少量遗传差异较明显的菌株。通过分析这些差异较明显的菌株与地理来源、品种等因素的关联发现,遗传差异较大的供试菌株,其群体基因型划分与地理来源没有直接的关系,但与海拔高度、烟草品种分布有一定的关联,这与前人的一些研究结果相类似[7,11]。
导致少数烟草疫霉分化差异较大的原因目前尚不清楚,但根据该研究结果并联系生产实际可知,通过种苗或其他工具使内部病菌频繁交流可能是导致少数病菌分化差异较大的原因之一。在玉溪烟区的3 个主栽品种中,种植KRK26 这一品种的区域内有较多数量的Ⅰ型烟草疫霉,而云烟87 和K326 品种中Ⅰ型烟草疫霉相对较少。这可能与品种之间的抗病性差异相关,该结果也说明不同烟草疫霉与其互作的烟草寄主之间存在一定联系,这为农业生产提供了警示,即烟草品种不能单一化种植。若品种单一,当感病寄主遇到强致病性菌株时往往无法抵御,造成大面积的病害流行,导致大量减产甚至绝收。此外,通过基因型分类与生理小种鉴定相结合的检测技术快速鉴定植烟区不同的烟草疫霉生理小种,对后续的品种布局和种植规划具有重要的现实指导意义[14]。而气象因素与烟草疫霉种群遗传差异之间是否存在关联有待进一步深入研究。
猜你喜欢
草业科学(2022年9期)2022-10-21
新疆农业科学(2022年6期)2022-07-13
中国农学通报(2022年12期)2022-06-01
河南农业·综合版(2022年2期)2022-03-18
河南农业(2022年2期)2022-03-14
作物学报(2022年4期)2022-02-10
河南农业·综合版(2021年7期)2021-08-23
云南画报(2021年7期)2021-08-06
农产品市场周刊(2020年6期)2020-05-06
中学生物学(2019年7期)2019-10-17
