
脂代谢相关基因ABHD5在泛癌中的表达模式及其与预后和免疫浸润的相关性分析
2022-06-02 01:03陈宇涛许淼泓曹霁昭卢嘉茵
中山大学学报(医学科学版) 2022年3期
陈宇涛,杨 洋,夏 淦,许淼泓,曹霁昭,卢嘉茵,邹 焱,杨 霞,3
(1.中山大学中山医学院生物化学与分子生物学教研室,广东广州 510080;2.广州市妇女儿童医疗中心,广东广州 510623;3.广东省基因操作和生物大分子产物工程技术研究中心,广东广州 510080)
代谢基因的异常激活或失活是肿瘤的标志之一,代谢基因可以通过代谢或非代谢途径对肿瘤的发生发展产生重要影响。ABHD5 基因编码CGI-58 蛋白,CGI-58 是一种细胞内脂解激活剂,作为脂肪甘油三酯脂肪酶ATGL(又称为PNPLA2)的辅因子,参与催化脂肪分解。ATGL 需要ABHD5 才能实现完整的甘油三酯(triglyceride,TG)水解酶活性[1]。人ABHD5 突变导致伴鱼鳞病的中性脂肪沉积症(neutral lipid storage disease with ichthyosis,NLSDI),又称为Chanarin-Dorfman 综合征[2]。已有文献报道,ABHD5 在结直肠癌(colorectal carcinoma,CRC)中是一种新的肿瘤抑制因子,CRC 细胞通过抑制ABHD5 介导的细胞内脂肪分解将代谢模式转变为有氧糖酵解,并通过抑制ABHD5 进而抑制AMPKα-p53 通路,诱导癌细胞发生上皮间质转化,因而ABHD5 具有抑制CRC 发生和转移的作用[3]。此外,ABHD5通过与BECN1直接相互作用促进自噬,减少DNA 损伤和基因组不稳定性,抑制CRC 肿瘤发生[4]。ABHD5 还在抑制c-Met激活和CRC干性中发挥关键作用[5]。然而,也有研究表明ABHD5 在子宫内膜癌中上调且高水平的ABHD5 表达与较短的总生存期显著相关[6],提示ABHD5 在不同肿瘤中的表达和作用存在差异。目前还没有关于ABHD5 与不同癌症类型之间关联的泛癌研究,因此探讨ABHD5 在不同肿瘤发生发展及预后中的作用具有重要意义。本研究综合利用各类数据库分析泛癌组织中ABHD5 mRNA 和蛋白表达水平的差异,初步探讨其对肿瘤预后的预测价值及其是否参与调节肿瘤相关的免疫反应,为进一步明确潜在的肿瘤免疫治疗靶点提供理论依据。
1 材料与方法
1.1 TCGA 联合GTEx 数据库、TIMER2.0 数据库及UALCAN数据库分析泛癌ABHD5基因表达量
GTEx(Genotype-tissue expression)数据库收录了样本基因在不同组织中的差异表达情况,TCGA数据库收录了基因在不同肿瘤组织中的测序数据[7-8]。在UCSC xene 网站(https://xenabrowser.net/datapages/)提 取GTEx 及TCGA 中TPM 格式的RNA-seq 数据,log2 转化后进行样本间的表达比较。使用TIMER2.0(http://timer.cistrome.org/)数据库比较不同肿瘤样本和正常组织中基因表达量的差异。UALCAN 数据库(http://ualcan.path.uab.edu/)可以访问临床蛋白质组学肿瘤分析协会联盟(CPTAC)数据集的蛋白质表达数据[9],本研究通过CPTAC 数据集比较ABHD5在不同肿瘤样本和正常组织中的蛋白表达水平。
1.2 TCGA 数据库分析ABHD5 与肿瘤临床病理分期的相关性
从GDC 网站(https://portal.gdc.cancer.gov/)获取TCGA数据库中FPKM(fregments per kilobase per million)格式的RNAseq 数据和临床数据(临床变量为T stage,N stage,M stage,Pathologic stage),使用R 语言将FPKM 格式的RNAseq数据转换成TPM(transcripts per million reads)格式,进行log2转化并进行可视化和统计分析。
1.3 Kaplan-Meier Plotter、Oncolnc 及R2 数据库分析ABHD5与预后指标的相关性
Kaplan-Meier Plotter 在线分析网站囊括GEO、EGA 和TCGA数据库中的不同肿瘤类型基因表达与临床预后信息。为了评估ABHD5 的临床预后价值,在Kaplan-Meier Plotter数据库中将患者样本按照基因表达的中位数分为高表达与低表达2 组,分析ABHD5 的表达与肿瘤总生存期(overall survival,OS)、无复发生存期(relapse free survival,RFS)、首次进展(first progression,FP)、进展后生存期(post progression Survival,PPS)的相关性,计算log-RANKP值和95%置信区间(Cl)的风险比率(HRs)。将ABHD5输入到oncolnc网站(http://www.Oncolnc.org/)进行生存分析,计算log-RANKP值。R2 网站(https://hgserver1.amc.nl/cgi-bin/r2/main.cgi)提供大量转录组数据集,可以从基因差异表达,相关通路,生存分析等多种层面开展肿瘤信息学基础分析,将ABHD5 输入到R2 网站,分析其在神经母细胞瘤中与肿瘤总生存期(OS)及无事件生存期(Event free survival,EFS)的相关性。
1.4 TCGA 数据库及TIMER2.0 分析ABHD5 表达与免疫细胞浸润及免疫检查点基因表达的相关性
从GDC 网站(https://portal.gdc.cancer.gov/)获取TCGA数据库中FPKM 格式的RNAseq 数据,使用R 语言转换成TPM 格式,并进行log2转化,利用24 种免疫细胞的markers[10]分析ABHD5与免疫细胞相关性,并进行可视化和统计分析。TIMER2.0(http://timer.cistrome.org/)是系统分析多种癌症类型免疫细胞浸润的在线资源数据库,应用反卷积方法作为统计方法,利用基因表达谱推断肿瘤组织中免疫细胞的浸润丰度[11]。用TIMER 2.0在线工具探索ABHD5表达与各个免疫细胞浸润的关系,并选取在各个数据集中结论一致的肿瘤,用相关性曲线进行可视化。并用TIMER 2.0在线工具探索ABHD5 表达与多种免疫检查点基因表达的相关性。
1.5 ABHD5在肿瘤中相关基因的功能富集分析
String 数据平台(https://string-db.org/)是收集了蛋白质-蛋白质相互作用信息的数据库,其相互作用包括物理性的直接作用和功能性的间接作用。使用STRING 数据库获得50 个经实验评估的ABHD5 结合蛋白,并使用GEPIA 在线数据库(http://gepia.cancer-pku.cn/)[12]分析100个ABHD5共表达基因,其中六个代表性靶基因与ABHD5 表达的相关性用散点图和热图表示。联合以上两个数据库的150 个ABHD5 相关分子,通过基因本体论数据库(gene ontology,GO)及京都基因与基因组数据库(kyoto encyclopedia of gene and genomes,KEGG)进行富集分析。GO分析通过生物过程(BP)、细胞组分(CC)以及分子功能(MF)三个层面进行,KEGG 是一种基于人体的通路数据库,GO 和KEGG 分析均通过R语言包实现。
1.6 免疫印迹法检测人正常细胞及肿瘤细胞中ABHD5的蛋白表达
人神经母细胞瘤细胞株SH-SY5Y、SK-N-BE(2)、人正常胰腺导管上皮细胞株HPDE6-C7、人胰腺癌细胞株PANC1 购买自ATCC 细胞库。兔抗人ABHD5 抗体(12201-1-AP)、鼠抗人GAPDH 抗体(60004-1)购自Proteintech 公司。实验步骤按SDS 蛋白裂解液裂解细胞、蛋白定量、电泳、电转、质量分数7%脱脂奶粉封闭、TBST 洗、孵育一抗、TBST洗、孵育二抗、TBST洗、ECL化学发光液显影等步骤进行,用Bio-rad化学发光成像仪拍照。
1.7 免疫组织化学法检测神经母细胞瘤组织中ABHD5的表达
本研究所用到的神经母细胞瘤临床组织标本均来自广州市妇女儿童医疗中心,其中节细胞神经瘤(ganglioneuroma,GN)、节细胞神经母细胞瘤(ganglioneuroblastoma,GNB)、神经母细胞瘤(neuroblastoma,NB)的标本例数分别为35例、34例和71例。所有提供组织标本患者均签署知情同意书,并经广州市妇女儿童医疗中心人类伦理委员会批准。临床组织标本交由上海芯超生物科技有限公司制作成组织芯片(tissue microarray,TMA)用于后续的免疫组化检测,排除脱片及临床信息不全的组织标本,其余标本用于结果分析。鼠抗人ABHD5抗体(sc-376931)购自Santa cruz公司,免疫组化具体方法及结果判断参照课题组前期论文[13]。
1.8 统计学分析与可视化
比较ABHD5 在膀胱尿路上皮癌(BLCA)不同临床分期中的表达差异,四组间比较时,采用Kruskal WallisH秩和检验,差异有统计学意义时采用Dunnett 法进行两两比较,采用Bonferroni 法校正显著性水平;两组间比较时,数据总体服从正态分布,采用t检验进行两组间比较;采用R 软件包ggplot2(3.3.3 版本)进行可视化。采用Kaplan-Meier 生存曲线评估ABHD5 低表达组和高表达组患者的生存时间,采用log-rank 检验进行两组间比较。在绘制免疫浸润棒棒糖图时,使用R软件包GSVA(1.34.0版本)分析ABHD5表达与免疫细胞浸润的相关性,并采用Spearman 秩相关分析。免疫印迹(western blotting,WB)、免疫组化定量分析中,两组间比较时,数据总体服从正态分布,则采用t检验进行两组间比较,否则采用Wilcoxon 秩和检验进行两组间比较,三组间比较时,数据总体服从正态分布,则采用单因素方差分析,否则采用Kruskal WallisH秩和检验。以P<0.05 作为所有统计分析中差异有统计学意义的阈值。
2 结果
2.1 泛癌中ABHD5 的mRNA 和蛋白水平的差异表达
为了明确泛癌中ABHD5 的mRNA 和蛋白表达,用GTEx 联合TCGA 的RNA-Seq 数据及TIMER2.0 数据库分析肿瘤和临近正常组织中ABHD5 mRNA 的差异表达模式,结果显示,ABHD5 mRNA 表达量在肾上腺皮质癌(adrenocortical carcinoma,ACC)、乳腺浸润癌(breast invasive carcinoma,BRCA)、弥漫性大B细胞淋巴瘤(lymphoid neoplasm diffuse large B-cell lymphoma,DLBC)、头颈鳞状细胞癌(head and neck squamous cell carcinoma,HNSC)、肾嫌色细胞癌(kidney chromophobe,KICH)、肾透明细胞癌(kidney renal clear cell carcinoma,KIRC)、急性髓细胞样白血病(acute myeloid leukemia,LAML)、肺腺癌(lung adenocarcinoma,LUAD)、肺鳞癌(lung squamous cell carcinoma,LUSC)、睾丸癌(testicular germ cell tumors,TGCT)、甲状腺癌(thyroid carcinoma,THCA)、胸腺癌(thymoma,THYM)中显著下调,在宫颈鳞癌和腺癌(cervical squamous cell carcinoma and endocervical adenocarcinoma,CESC)、多形成性胶质细胞瘤(glioblastoma multiforme,GBM)、脑低级别胶质瘤(brain lower grade glioma,LGG)、子宫内膜癌(uterine corpus endometrial carcinoma,UCEC)中显著上调(图1)。用CPTAC 数据库分析肿瘤和临近正常组织中ABHD5 的蛋白水平,结果显示,与正常组织相比,ABHD5 蛋白表达在乳腺癌、结肠癌、头颈鳞癌、肝细胞癌、肺腺癌、卵巢癌、胰腺癌的原发灶组织中显著降低,但是在肾透明细胞癌、多形性胶质母细胞瘤的原发灶组织中显著升高(图2)。

图1 采用TCGA联合GTEx数据库和TIMER2.0数据库分析不同肿瘤组织和癌旁正常组织中ABHD5 mRNA表达情况Fig.1 Expressions of ABHD5 mRNA in different tumor tissues and adjacent normal tissues analyzed by TCGA,GTEx and TIMER2.0 databases

图2 根据CPTAC数据库比较不同肿瘤原发灶组织和癌旁正常组织中ABHD5蛋白表达水平Fig.2 Expressions of ABHD5 protein levels in different tumor tissues and adjacent normal tissues analyzed by CPTAC database
使用免疫组织化学法检测神经母细胞瘤组织中ABHD5 的表达,结果显示,与MYCN 非扩增型(MYCN non-amplified,Non-MNA)神经母细胞瘤组织相比,MYCN 扩增型(MYCN amplified,MNA)神经母细胞瘤组织中ABHD5 的表达量显著上调(U=29.50,Z=-2.061,P=0.039;图3)。
使用免疫印迹法检测神经母细胞瘤细胞、人正常胰腺导管上皮细胞及胰腺癌细胞中ABHD5 的表达,结果显示MYCN 非扩增型SH-SY5Y 细胞的ABHD5 表达水平与MYCN 扩增型细胞SK-N-BE(2)相比较低(t=4.589,P=0.010);与人正常胰腺导管上皮细胞HPDE6-C7 相比,ABHD5 在人胰腺癌细胞PANC1 中表达较低(t=9.196,P<0.001;图4)。
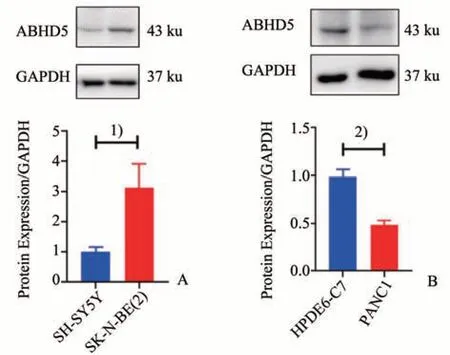
图4 Western Blot检测不同细胞系中ABHD5的蛋白表达水平Fig.4 Expressions of ABHD5 protein levels in different cell lines were tested by Western Blot
2.2 ABHD5 在膀胱尿路上皮癌不同临床分期中的表达差异
使用TCGA数据库探索ABHD5表达与肿瘤临床分期之间的关联,结果显示,在膀胱尿路上皮癌(bladder urothelial carcinoma,BLCA)中,与T1 期相比,ABHD5 在T2、T3、T4 期患者肿瘤中表达下调(H=9.019,P=0.029,T1 期与T3 期比较,P.adj=0.034);与N0、N1、N2 期相比,ABHD5 在N3 期患者肿瘤中表达下调(H=13.095,P=0.004,N2 期与N3 期比较,P.adj=0.009);与病理分期Ⅰ期相比,ABHD5在Ⅱ、Ⅲ、Ⅳ期患者肿瘤中表达下调(H=8.838,P=0.032,Ⅰ期与Ⅲ期比较,P.adj=0.041;图5)。

图5 采用TCGA数据库分析BLCA患者肿瘤组织中ABHD5 mRNA在不同临床分期中的表达差异Fig.5 Expression levels of ABHD5 in BLCA patients with different clinical stages analyzed by TCGA database
2.3 ABHD5在癌症中的预后价值
本研究进一步评估ABHD5 表达量与肿瘤患者预后之间的关系,Kaplan-Meier Plotter数据库mRNA gene chip预后分析显示,ABHD5 高表达在肺癌中与较好的OS、PPS 相关,在胃癌中与较好的FP 相关;RNA-seq 预后分析显示,ABHD5 高表达在卵巢癌中与较好的OS 相关,在肺鳞状细胞癌中与较差的RFS 相关。在oncolnc 数据库(基于TCGA 数据库)的预后分析显示,ABHD5 高表达在结肠癌(colon adenocarcinoma,COAD)、肾乳头状细胞癌(kidney renal papillary cell carcinoma,KIRP)、膀胱尿路上皮癌(BLCA)、卵巢浆液性囊腺癌(ovarian serous cystadenocarcinoma,OV)、肝细胞癌(liver hepatocellular carcinoma,LIHC)中与较好的预后相关。R2数据库的预后分析显示,ABHD5高表达在神经母细胞瘤中与其较差的预后相关(图6)。

图6 利用生存曲线评估ABHD5的预后价值Fig.6 Survival curve evaluating the prognostic value of ABHD5
2.4 ABHD5 表达与免疫浸润及免疫检查点基因表达的相关性
本研究使用TCGA数据库评估ABHD5表达水平在泛癌中与肿瘤组织免疫细胞浸润水平的相关性,发现ABHD5 与结肠癌(COAD)、弥漫性大B 细胞淋巴瘤(DLBC)、急性髓细胞样白血病(LAML)、脑低级别胶质瘤(LGG)、子宫肉瘤(uterine carcinosarcoma,UCS)、卵巢浆液性囊腺癌(OV)、胰腺癌(pancreatic adenocarcinoma,PAAD)、直肠腺癌(rectum adenocarcinoma,READ)、胃 癌(stomach adenocarcinoma,STAD)组织的多种免疫细胞浸润呈明显正相关关系。使用TIMER2.0数据库进一步评估40种癌组织中ABHD5 表达水平与各类免疫细胞浸润水平的相关性,发现ABHD5 在泛癌中与中性粒细胞浸润呈密切正相关关系(图7)。本研究使用TIMER2.0 数据库评估ABHD5 在肿瘤中与免疫检查点基因表达的相关性,结果表明ABHD5 表达在多种肿瘤中与多种免疫检查点基因表达正相关,其中以CD274、IL10RB、KDR、TGFBR1 基因最为显著,在结肠癌(COAD)、胰腺癌(PAAD)、胃癌(STAD)中ABHD5 与免疫检查点基因表达的正相关性最为显著(图7)。

图7 ABHD5表达与免疫细胞浸润的相关性及免疫检查点基因表达水平Fig.7 Correlation analyses of ABHD5 expression levels with immune cell infiltration and immune checkpoint genes
2.5 ABHD5在癌症中的功能富集分析
使用STRING 数据库获得经实验评估的ABHD5结合蛋白。使用GEPIA 数据库分析100 个ABHD5 相关基因,其中六个代表性靶基因(ARL8B、OXSR1、UBP1、NEK4、EIF4E、ZNF35)与ABHD5 表达在多肿瘤中呈显著正相关。根据ABHD5 结合及共表达基因进行GO 和KEGG 分析,结果显示ABHD5 共表达基因在肿瘤中与长寿调节途径(longevity regulating pathway)、mRNA 监视途径(mRNA surveillance pathway)、内吞作用(endocytosis)等KEGG 信号通路相关。GO 富集分析结果表明,ABHD5 共表达基因与蛋白去烷基化(protein dealkylation)、组蛋白赖氨酸去甲基化(histone lysine demethylation)、脂滴(lipid droplet)、组蛋白甲基转移酶复合体(histone methyltransferase complex)、GDP 结合(GDP binding)密切相关(图8)。

图8 ABHD5在肿瘤中相关基因的GO和KEGG功能富集分析Fig.8 ABHD5 associated gene enrichment pathway analysis
3 讨论
ABHD5(CGI-58),是脂肪甘油三酯脂肪酶ATGL(PNPLA2)的辅因子,近年研究发现ABHD5 可通过代谢或非代谢途径影响肿瘤的发生发展。为了进一步明确ABHD5 在肿瘤中的作用,本研究对ABHD5 在泛癌中的表达模式、与临床分期、预后、免疫浸润的相关性进行分析,并对其相关基因进行功能富集分析,结果提示ABHD5 不仅是肿瘤预后预测因子,也是肿瘤免疫调节因子。
本研究首先使用GTEx 联合TCGA 数据库及TIMER2.0数据库分析ABHD5 在泛癌中的mRNA 表达水平,结果表明ABHD5 mRNA 水平在肾上腺皮质癌(ACC)、乳腺浸润癌(BRCA)、弥漫性大B 细胞淋巴瘤(DLBC)、头颈鳞状细胞癌(HNSC)、肾嫌色细胞癌(KICH)、肾透明细胞癌(KIRC)、急性髓细胞样白血病(LAML)、肺腺癌(LUAD)、肺鳞癌(LUSC)、睾丸癌(TGCT)、甲状腺癌(THCA)、胸腺癌(THYM)中显著下调,在宫颈鳞癌和腺癌(CESC)、多形成性胶质细胞瘤(GBM)、脑低级别胶质瘤(LGG)、子宫内膜癌(UCEC)中显著上调。CPTAC 数据库分析结果表明,ABHD5蛋白表达在乳腺癌、结肠癌、头颈鳞癌、肝细胞癌、肺腺癌、卵巢癌、胰腺癌的原发灶组织中显著降低,但是在肾透明细胞癌、多形性胶质母细胞瘤的原发灶组织中显著升高。免疫组织化学结果显示,与MYCN 非扩增型(MYCN nonamplified,Non-MNA)神经母细胞瘤组织相比,MYCN 扩增型(MYCN amplified,MNA)神经母细胞瘤组织中ABHD5 的表达量显著上调。免疫印迹结果表明,与MYCN 扩增型SK-N-BE(2)细胞相比,MYCN 非扩增型SH-SY5Y 细胞的ABHD5 表达水平较低;与人正常胰腺导管上皮细胞HPDE6-C7 相比,ABHD5 在人胰腺癌细胞PANC1 中的表达较低。TCGA 数据库分析结果表明,在膀胱尿路上皮癌(BLCA)中,与T1 期相比,ABHD5 在T2、T3、T4 期患者肿瘤中表达下调;与N0、N1、N2 期相比,ABHD5 在N3 期患者肿瘤中表达下调;与病理分期Ⅰ期相比,ABHD5 在Ⅱ、Ⅲ、Ⅳ期患者肿瘤中表达下调。Kaplan-Meier Plotter 数据库mRNA gene chip 预后分析显示,ABHD5 高表达在肺癌中与其较好的OS、PPS 相关,在胃癌中与其较好的FP 相关;RNA-seq 预后分析显示,ABHD5 高表达在卵巢癌中与其较好的OS 相关,在肺鳞状细胞癌中与其较差的RFS 相关。oncolnc 数据库中的预后分析显示,ABHD5 高表达在结肠癌(COAD)、肾乳头状细胞癌(KIRP)、膀胱尿路上皮癌(BLCA)、卵巢浆液性囊腺癌(OV)、肝细胞癌(LIHC)中与其较好的预后相关。R2数据库的预后分析显示,ABHD5高表达在神经母细胞瘤中与其较差的预后相关。
结合各类数据库分析及实验结果,ABHD5 表达量在结直肠癌、卵巢癌、肺腺癌中显著降低,在膀胱尿路上皮癌中ABHD5 在其较高的临床分期中表达较低,且其高表达与较好的预后有关;ABHD5 表达量在胰腺癌的原发灶组织中显著降低,与人正常胰腺导管上皮细胞相比,ABHD5 蛋白在人胰腺癌细胞中表达较低,这提示ABHD5 在结直肠癌、卵巢癌、肺腺癌、膀胱尿路上皮癌、胰腺癌中可能发挥抑癌基因的作用。在宫颈鳞癌和腺癌、多形成性胶质细胞瘤、脑低级别胶质瘤、子宫内膜癌中,ABHD5 的表达量上调,可能发挥促癌基因的作用。而在神经母细胞瘤中,原癌基因MYCN 编码转录因子N-myc,其扩增状态是影响患者预后的重要因素,当患者年龄大于一岁、临床分期较高并伴有MYCN 扩增时,通常提示预后不良[14]。MYCN 可以调控神经母细胞瘤中P53、S 期相关蛋白激酶2(S-phase associated kinase 2,SKP2)、鸟氨酸脱羧酶1(ornithine decarboxylase 1,ODC1)、双微体2(murine double minute 2,MDM2)等分子的表达,还可调节微小RNA(miRNA)及长链非编码RNA(lncRNA)[15],MYCN 基因扩增还与多药耐药相关蛋白(multidrug resistence-associated protein,MRP)高表达相关[16]。MYCN 基因上调通过加速神经母细胞瘤细胞细胞增殖、迁移和侵袭,调节细胞周期、分化、凋亡等,促进神经母细胞瘤演进。许多基因的失调,也可导致MYCN 的扩增,其互相调控的机制是神经母细胞瘤研究的热点,但机制非常复杂,至今仍不明确。本研究发现与MYCN 非扩增型神经母细胞瘤组织及细胞相比,MYCN 扩增型神经母细胞瘤组织及细胞中ABHD5的表达量显著上调,且R2数据库分析提示ABHD5 高表达与神经母细胞瘤较差的预后相关,提示ABHD5可能是与神经母细胞瘤预后不良相关的潜在调控分子,其相关功能可能与MYCN复杂的分子调控机制有关。
肿瘤微环境(tumor microenvironment,TME)由肿瘤细胞、基质细胞(包括肿瘤相关成纤维细胞和间充质基质细胞)、免疫细胞(淋巴和髓系细胞)及细胞外基质组成[17]。正常情况下,免疫细胞可以识别并清除异常的肿瘤细胞[18],但是肿瘤细胞为存活和生长能采用不同策略,抑制人体的免疫系统的抗肿瘤效果,TME 中的基质细胞和免疫细胞通过自分泌和旁分泌,诱导细胞外基质硬度增加、血管和淋巴管形成、坏死区形成和肿瘤转移[19],因此免疫细胞在肿瘤发展过程中起着至关重要的作用。ABHD5 长期以来一直被认为是PNPLA2 的辅助因子。近期也有研究逐渐揭示了ABHD5在癌症领域的独立于PNPLA2的作用[4-5]。本研究从肿瘤免疫调节出发,利用TCGA 数据库对ABHD5 与免疫细胞标志物相关性进行分析,结果显示:ABHD5基因与9种肿瘤组织的免疫细胞浸润呈明显正相关关系,分别为结肠癌(COAD)、弥漫性大B 细胞淋巴瘤(DLBC)、急性髓细胞样白血病(LAML)、脑低级别胶质瘤(LGG)、子宫肉瘤(UCS)、卵巢浆液性囊腺癌(OV)、胰腺癌(PAAD)、直肠腺癌(READ)及胃癌(STAD);TIMER2.0 数据库分析结果显示,在37 种癌组织中ABHD5 表达水平与中性粒细胞浸润水平呈明显正相关关系,该结果提示ABHD5 是与中性粒细胞浸润相关的泛癌预后生物标志物。中性粒细胞占循环白细胞的70%,是抵御病原体的第一道防线[20],肿瘤相关中性粒细胞(tumor associated neutrophils,TANs)对肿瘤具有促瘤和抑瘤的双重作用[21],TANs 在癌症中的复杂作用不仅包括促进或抑制肿瘤进展的两面性,还包括各种极化状态及与其他免疫细胞的相互作用,具有作为临床生物标志物和治疗靶点的潜力,针对特定炎症条件设计中性粒细胞相关抑制剂可能为肿瘤靶向治疗提供新思路[22]。
免疫监视即免疫系统具有的识别、杀伤并及时清除体内突变细胞,防止肿瘤发生的功能,影响癌症患者的预后,肿瘤细胞通过利用免疫检查点伪装成人体正常细胞,从而逃避免疫监视;通过阻断免疫检查点途径阻止肿瘤细胞伪装成人体的正常组分,是实现抗肿瘤免疫的有效方法[23]。本文探讨了ABHD5 表达水平与免疫检查点基因表达水平的关系,结果提示ABHD5 可能通过调节特定免疫检查点基因的表达来调节各种肿瘤中的肿瘤免疫模式,ABHD5 在子宫内膜癌(UCEC)中上调,高表达的ABHD5 与免疫检查点基 因CD274、CD96、IL10RB、KDR、PDCD1LG2、TGFBR1、VTCN1 的表达正相关;ABHD5 在多形成性胶质细胞瘤(GBM)中上调,高表达的ABHD5 与免疫检查点基因CD274、CSF1R、KDR、PDCD1LG2 的表达正相关,这提示在高表达ABHD5 的肿瘤中阻断特定的免疫检查点可能为这些肿瘤的免疫治疗提供新思路。
相关性基因筛选提示ABHD5 在肿瘤中与小GTP 酶ARL8B、氧化应激反应蛋白OXSR1、上游结合蛋白UBP1、NIMA 相关激酶NEK4、真核转录因子EIF4E、锌指蛋白ZNF35的表达呈显著正相关关系。其中ARL8B是抗前列腺癌的药物作用靶标,降低ARL8B 的表达会显著提升细胞的自噬水平,进而导致肿瘤消亡[24],此外,ARL8B 介导的溶酶体运输促进了在辐射中存活的癌细胞的侵袭[25]。提示了ABHD5 可能在自噬、溶酶体运输方面发挥潜在作用。TFCP2/TFCP2L1/UBP1转录因子在癌症、阿尔茨海默病和其他疾病中发挥重要作用,可能成为药物开发有吸引力的靶标[26-27],也可为探索ABHD5 在肿瘤中表达量发生变化的原因或其发挥作用的机制提供可能线索。功能富集分析提示ABHD5 表达在多肿瘤中与组蛋白去甲基化相关,已有研究表明ABHD5 在细胞质中与SET1A 甲基转移酶复合物的核心亚基DPY30 相互作用,抑制DPY30 的核转位和SET1A 的活性,阻止了SET1A 介导的YAP 和组蛋白H3 甲基化,降低染色质的可及性及YAP 减少诱导的c-Met 转录,从而减少CRC 细胞干性[5]。ABHD5 与长寿调节途径、mRNA 监视途径、内吞作用及蛋白质去烷基化的相关性鲜有报道,其调控机制有待进一步阐明。
综上所述,ABHD5 在多种肿瘤组织中表达下调,在少数肿瘤组织中表达上调,可能成为潜在的肿瘤预后预测因子,且ABHD5 在肿瘤发生发展过程中发挥潜在的免疫调节因子作用,与中性粒细胞浸润存在较强的相关性。本研究结果为进一步了解泛癌中ABHD5 的功能提供了初步的依据,其具体生物学作用及机制还需进一步的实验验证。
猜你喜欢
延安大学学报(医学科学版)(2022年2期)2022-11-23
医学信息(2022年17期)2022-10-11
临床军医杂志(2022年7期)2022-08-06
健康护理(2022年3期)2022-05-26
中国典型病例大全(2022年11期)2022-05-13
计算机系统应用(2022年4期)2022-05-10
癌症进展(2021年9期)2021-07-20
科学24小时(2020年11期)2020-12-14
家庭医药(2020年5期)2020-05-14
电脑知识与技术(2018年18期)2018-11-01
