
miR-495-3p对山羊骨骼肌细胞增殖分化的影响
2022-06-01 03:59杨丽丽张琼文刘笑笑江明生
中国畜牧兽医 2022年4期
杨丽丽,张琼文,张 瑜,刘笑笑,陈 婷,江明生
(广西大学动物科学技术学院,南宁 530004)
在山羊生长发育过程中,骨骼肌的生长发育尤为重要,不仅影响山羊生产性能和肉品质,而且对畜牧业的经济效益至关重要。因此,研究动物骨骼肌生长发育的调控机制成为育种工作的重要内容。近年来,越来越多的报道集中于研究非编码RNA对动物骨骼肌生长发育的调控机制,microRNA(miRNA)是其中研究热点之一。
miRNA是一种非编码RNA小分子,长度19~22 nt,广泛存在于病毒、动物和植物细胞中,主要在转录后发挥作用[1]。研究证明,miRNA参与骨骼肌生长发育,在细胞的增殖、分化等过程中起重要作用[2]。研究报道,大部分miRNA具有组织和不同阶段表达模式,只在特定组织和时期表达[3],如miR-1、miR-206、miR-133a等在心肌和骨骼肌中特异性表达,参与骨骼肌细胞增殖和分化[4];miR-1通过调控细胞蛋白周期D1,进而促进细胞增殖[5],通过靶向配对盒基因(Pax7)和组蛋白去乙酰化酶4(HDAC4)抑制Pax7和HDAC4的表达,从而促进牛骨骼肌细胞的分化[6]。李丹丹[7]研究表明,miRNA-101a能够促进山羊骨骼肌卫星细胞的分化,但对卫星细胞的增殖作用不显著。miR-495-3p作为一个重要的调节因子,在细胞增殖分化中起重要作用,参与转录调控,如miR-495-3p通过作为lncRNA NEAT1的分子海绵影响靶基因E2F转录因子3(E2F transcription factor 3,E2F3)的表达,从而促进黑素瘤细胞的增殖、迁移和侵袭[8]。研究发现,miR-495-3p能够与核基质结合区结合蛋白质1(SATB1)基因的3′-UTR结合,并与lncRNA-UCA1构成内源竞争RNA(competing endogenous RNAs,ceRNA)网络,通过对靶基因的调节促进胃癌细胞的增殖与侵袭[9]。在体外培养的血管平滑肌细胞(VSMC)中过表达肌生长抑制素(Myostatin),可使miR-495-3p表达量下降,推测miR-495-3p可能与肌细胞增殖相关[10]。以上报道表明,miRNA可以调控骨骼肌卫星细胞的增殖与分化,但是miRNA对隆林山羊骨骼肌卫星细胞的作用机制报道较少。
隆林山羊是广西地方品种,主要分布于云贵高原桂西北山区,具有极强的环境适应能力,但其生长速度慢,产肉量低,导致生产性能低。1月龄是山羊生长发育关键期,10月龄是成熟期,选择这2个阶段对研究其生长发育具有重要意义。广西大学动科院遗传育种课题组前期测序结果发现,miR-495-3p与隆林山羊肌肉发育相关(未发表)。因此,本研究利用生物学信息方法预测miR-495-3p的靶基因,实时荧光定量PCR检测其对骨骼肌发育分化调控基因生肌因子5(Myf5)、肌细胞生成素(MyoG)表达的影响,双荧光素酶报告实验验证其结合位点,研究miR-495-3p对山羊骨骼肌细胞增殖分化的调控,为提高肉羊的生产性能提供参考,对畜牧业动物品种培育也具有重要的意义。
1 材料与方法
1.1 材料
1.1.1 试验动物与细胞 隆林山羊来自广西大学动物科技学院试验羊场,分别选择健康、体重相近的1和10月龄隆林山羊各3只;山羊骨骼肌卫星细胞、293T细胞、大肠杆菌DH5α感受态细胞均由广西大学动物科技学院动物遗传育种实验室保存。
1.1.2 主要试剂及仪器 DMEM、胎牛血清(FBS)、特级马血清(FES)、0.25%胰蛋白酶、PBS、Lipofectamine 2000、DNA凝胶回收试剂盒、PrimeScriptTMⅡ 1st Strand cDNA Synthesis Kit均购自北京索莱宝科技有限公司;大肠杆菌DH5α感受态细胞购自北京全式金生物技术股份有限公司;氨苄青霉素购自上海华舜生物技术有限公司。空气恒温摇床(HY45)购自武汉科学仪器厂;荧光素酶检测仪(M200 PRO)购自广州博鹭腾生物科技有限公司。
基础培养基:90% DMEM+10% FBS;分化培养基:98% DMEM+2% FES。
1.1.3 质粒合成 根据miRbase上miR-495-3p的序列,运用化学方法合成miR-495-3p模拟物(miR-495-3p mimic)、模拟物对照(miR-495-3p mimic NC)及抑制物(miR-495-3p inhibitor)和抑制物对照(miR-495-3p inhibitor NC),均由广州锐博生物技术有限公司合成,ACTC1-WT+mimics NC、ACTC1-WT+miR-495-3p、ACTC1-Mut+mimics NC、ACTC1-Mut+miR-495-3p均由湖南普拉特泽生物科技有限公司合成。
1.2 方法
1.2.1 样品采集 屠宰过程按照《畜禽屠宰操作规程 羊》(NY/T 3469-2019)进行,在无菌状态下分离隆林山羊的心脏、肝脏、脾脏、肺脏、肾脏、背最长肌及腿肌组织,立即冻存至液氮中,在实验室将各组织整理后置-80 ℃保存备用。
1.2.2 miR-495-3p靶基因预测 利用生物信息学方法预测miR-495-3p的靶基因。
1.2.3 实时荧光定量PCR检测 参考GenBank中山羊的miR-495-3p、ACTC1、Pax7、Cyclin E、MyoG、Myf5的基因序列,利用Oligo 7.0软件设计引物,引物信息见表1,引物均由生工生物工程(上海)股份有限公司合成。根据Trizol法提取骨骼肌细胞1月龄山羊心脏、肝脏、脾脏、肺脏、肾脏、背最长肌、腿肌及10月龄山羊背最长肌总RNA,反转录合成cDNA,以U6为miRNA的内参,GAPDH为其他基因内参,用实时荧光定量PCR检测各基因相对表达量。 PCR反应体系:Premix ExTaqⅡ 5.0 μL,上、下游引物各0.3 μL,cDNA 2.5 μL,RNase-free ddH2O补至10 μL。PCR反应条件:95 ℃预变性30 s;95 ℃变性30 s,退火(温度见表1) 30 s,72 ℃延伸12 s,共35个循环。采用2-ΔΔCt法计算各基因相对表达量。

表1 引物信息Table 1 Primer information

续表
1.2.4 miR-495-3p及其靶基因的组织表达谱构建 以1月龄隆林山羊的心脏、肝脏、脾脏、肺脏、肾脏、背最长肌、腿肌cDNA进行实时荧光定量PCR,检验miR-495-3p及其靶基因在各组织中的表达水平。
1.2.5 山羊骨骼肌细胞培养、转染及分化 将冻存细胞复苏后加入培养基置37 ℃、5% CO2培养箱中培养,当细胞汇合度达到80%左右时,进行细胞传代。当传代细胞融合度达到60%~70%时,按照Lipofectamine 2000说明书进行转染,将miR-495-3p mimic、miR-495-3p inhibitor、miR-495-3p mimic NC、miR-495-3p inhibitor NC和双荧光载体转染细胞,48 h后收集一部分细胞,实时荧光定量PCR法检测过表达及干扰效率;另一部分传代细胞在培养24 h后更换含2% FES分化培养基,分化培养6 d收集细胞。用实时荧光定量PCR法检测肌细胞分化标志基因MyoG、Myf5的表达量,PCR反应体系和程序同1.2.3。
1.2.6 细胞增殖活力 将细胞均匀接种于96孔板,待细胞贴壁后,将miR-495-3p mimic、miR-495-3 pmimic NC、miR-495-3p inhibitor和miR-495-3p inhibitor NC根据1.2.5方法转染细胞,每组设置7个重复,以PBS溶液为空白对照;24 h后,收集细胞备用,或者每孔中加入10 μL CCK-8溶液,在培养箱继续孵育2 h,收集细胞,实时荧光定量PCR检测细胞增殖标志基因Pax7、CyclinE的表达量,PCR反应体系和程序同1.2.3;用酶标仪检测D450 nm值,并计算细胞增殖率。
细胞增殖率(%)=[(试验孔-空白孔)/(对照孔-空白孔)]×100%
1.2.7 双荧光素酶荧光活性分析 以psiCHECK2作为载体,根据预测到的miR-495-3p及其靶基因的结合位点,以XhoⅠ、NotⅠ为酶切位点,构建靶基因的野生型载体(ACTC1-WT+mimics NC、ACTC1-WT+miR-495-3p)和突变型载体(ACTC1-Mut+mimics NC、ACTC1-Mut+miR-495-3p);在24孔板进行293T细胞转染,每组设置3个重复孔,分组转入ACTC1-WT+mimics NC、ACTC1-WT+miR-495-3p、ACTC1-Mut+mimics NC、ACTC1-Mut+miR-495-3p质粒,在37 ℃培养箱中转染48 h。按照双荧光素酶检测试剂盒说明书测定miR-495-3p及其靶基因的结合情况。
1.3 数据统计分析
用SPSS 26.0软件进行单因素方差分析,并采用t检验进行多重比较,结果用平均值±标准差表示。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 隆林山羊肌肉发育相关miRNA和靶基因的筛选
生物信息学预测发现,miR-495-3p与肌肉发育相关的ACTC1基因有潜在结合位点(图1A),可能与肌肉发育有关。实时荧光定量PCR检测发现,miR-495-3p与ACTC1基因在1和10月龄背最长肌中表达量均差异极显著(P<0.01)(图1B、1C)。

①A,miR-495-3p与ACTC1结合预测图;B、C,miR-495-3p、ACTC1在1和10月龄背最长肌中的相对表达量。②*,差异显著(P<0.05);**,差异极显著(P<0.01);无*,差异不显著(P>0.05)。下同①A,Prediction diagram of the combination of miR-495-3p and ACTC1;B and C,The relative expression of miR-495-3p and ACTC1 in longissimus dorsi at 1 and 10 months old,respectively.②*,Significant difference (P<0.05);**,Extremely significant difference (P<0.01);No *,No significant difference(P>0.05).The same as below图1 miR-495-3p靶基因预测(A)及不同月龄山羊背最长肌中miR-495-3p(B)与ACTC1(C)的相对表达量Fig.1 Target gene prediction of miR-495-3p (A) and the expression levels of miR-495-3p (B) and ACTC1 (C) in longissimus dorsi muscle of goats at different growth ages
2.2 miR-495-3p与ACTC1基因的组织表达谱
实时荧光定量PCR结果表明,miR-495-3p与ACTC1基因在不同组织中均有表达,miR-495-3p在1月龄隆林山羊脾脏中表达量最高,依次为背肌、腿肌、肝脏、肾脏、心脏、肺脏,各组织间均有显著差异(P<0.05)(图2A);ACTC1基因在隆林山羊心脏中表达量最高,依次为背最长肌、肺脏、腿肌、脾脏、肾脏、肝脏,各组织间均有显著差异(P<0.05)(图2B)。
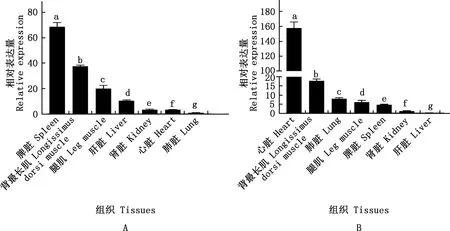
肩标不同字母表示差异显著(P<0.05)Values with different letter superscripts mean significant difference (P<0.05)图2 1月龄山羊miR-495-3p(A)和ACTC1(B)组织表达谱Fig.2 Tissue expression profile of miR-495-3p (A) and ACTC1 (B) in one-month-old goats
2.3 miR-495-3p过表达和干扰效率检测
实时荧光定量PCR检测结果表明,miR-495-3p mimic组miR-495-3p的相对表达量极显著高于miR-495-3p mimic NC组(P<0.01),miR-495-3p inhibitor组miR-495-3p的相对表达量极显著低于miR-495-3p inhibitor NC组(P<0.01)(图3)。
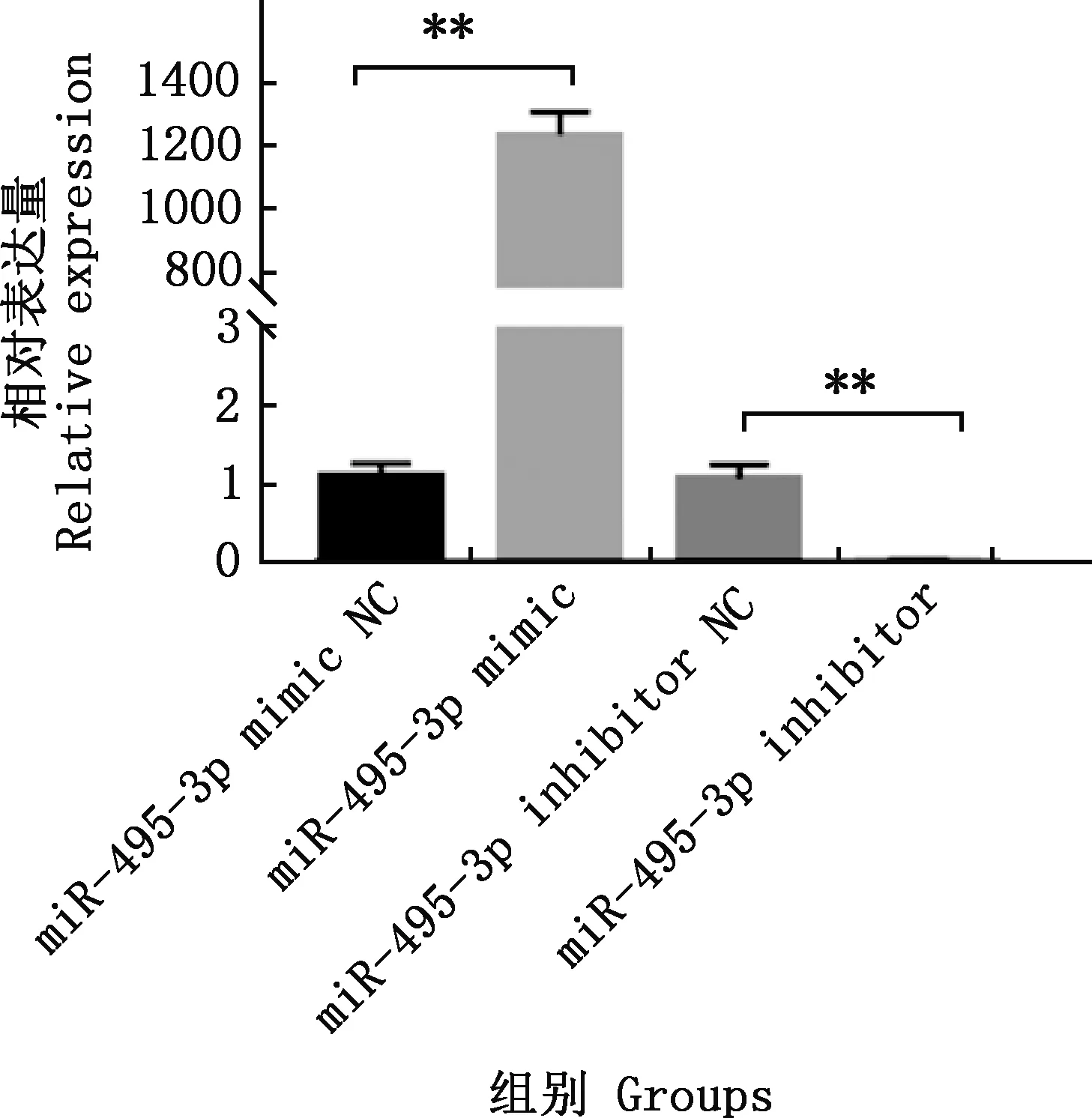
图3 miR-495-3p过表达和干扰效率Fig.3 Efficiency of miR-495-3p overexpression and inhibition
2.4 miR-495-3p过表达、干扰对细胞分化的影响
将miR-495-3pmimic、miR-495-3p inhibitor转染至骨骼肌细胞中,分化6 d时发现有明显的肌管产生,与miR-495-3p mimic NC组相比,miR-495-3p mimic组促进肌管的生成,miR-495-3p inhibitor组抑制肌管的生成(图4)。实时荧光定量PCR结果表明,miR-495-3p mimic组骨骼肌细胞分化标志基因MyoG、Myf5的表达水平显著高于miR-495-3p mimic NC组(P<0.05),miR-495-3p inhibitor组MyoG、Myf5基因的表达水平显著低于miR-495-3p inhibitor NC组(P<0.05)(图5)。
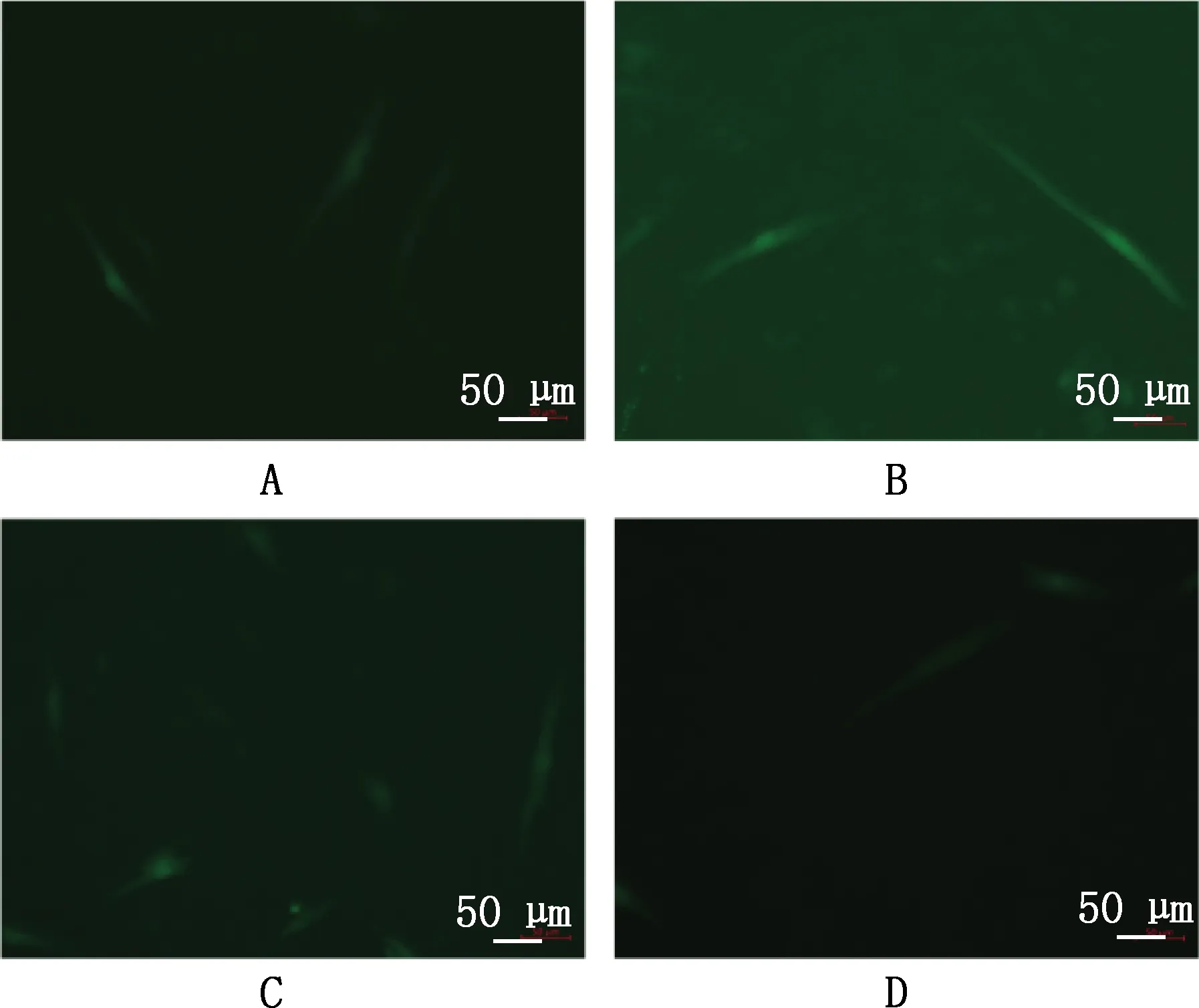
A~D,miR-495-3p mimic NC、miR-495-3p mimic、miR-495-3p inhibitor NC和miR-495-3p inhibitor组A-D,miR-495-3p mimic NC,miR-495-3p mimic,miR-495-3p inhibitor NC and miR-495-3p inhibitor groups,respectively图4 山羊骨骼肌卫星细胞转染后分化6 d的形态Fig.4 Morphology of goat skeletal muscle satellite cells after transfection and differentiation for 6 days
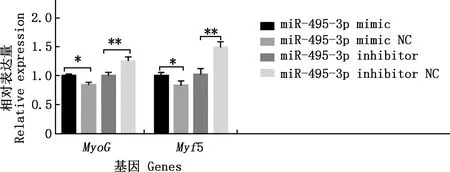
图5 各组山羊骨骼肌细胞分化标志基因的相对表达量Fig.5 Relative expression of differentiation marker genes in skeletal muscle cells of goats in each group
2.5 miR-495-3p过表达、干扰对细胞增殖的影响
实时荧光定量PCR检测结果表明,与对照组相比,miR-495-3p mimic和miR-495-3p inhibitor组Pax7、Cyclin E的表达量均无显著差异(P>0.05);CCK-8检测发现,miR-495-3p mimic和miR-495-3p inhibitor组对细胞增殖活力均没有显著影响(P>0.05)(图6)。
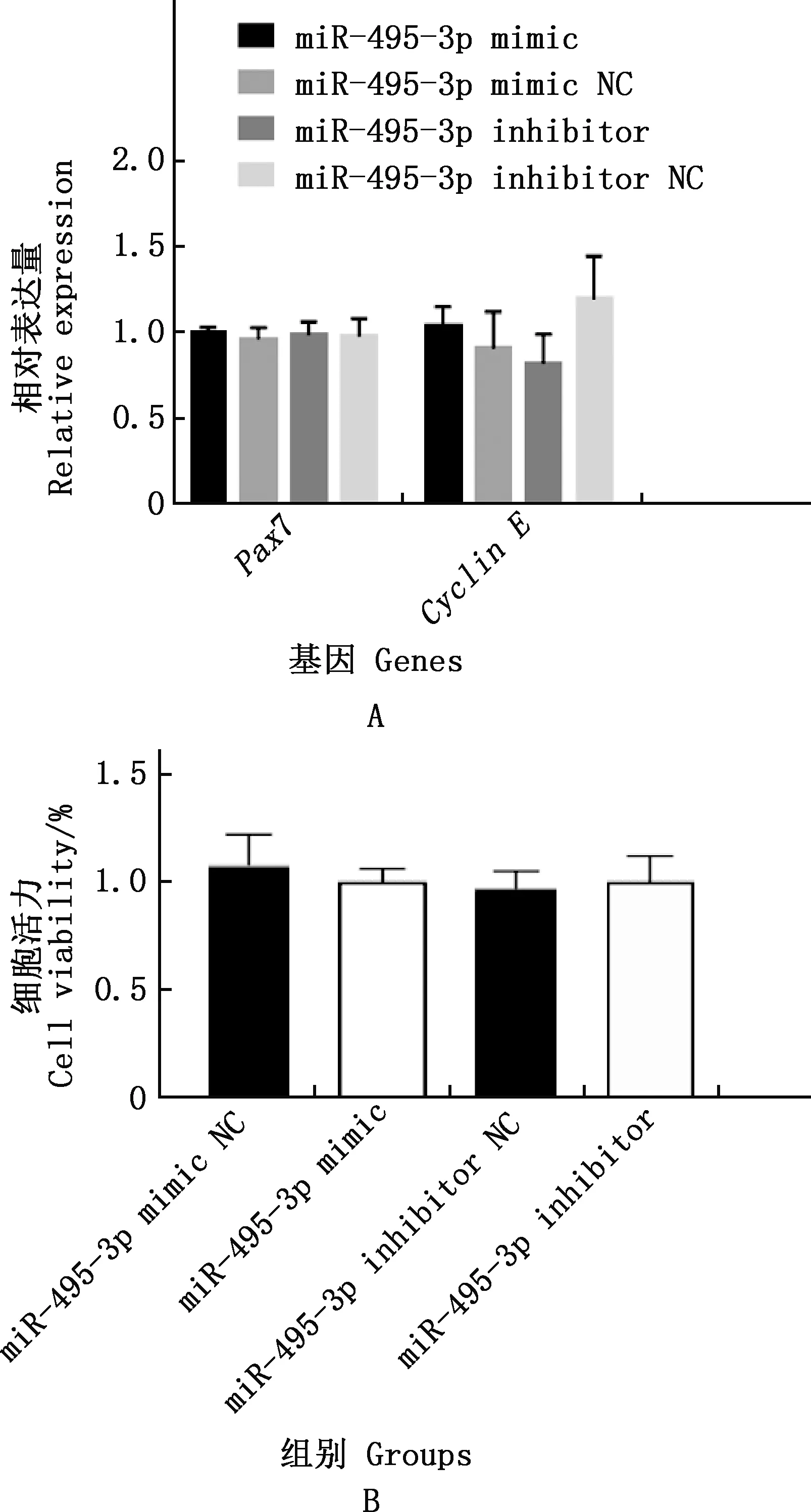
A,Pax7、Cyclin E基因相对表达量;B,细胞活力A,The relative expression of Pax7 and Cyclin E genes;B,Cell viability图6 各组山羊骨骼肌卫星细胞增殖相关基因的相对表达量及细胞活力Fig.6 The relative expression of proliferation-related genes and cell viability of goat skeletal muscle satellite cells in each group
2.6 双荧光素酶报告实验验证结合位点
双荧光素酶实验结果发现,与转染ACTC1-WT+mimics NC相比,共转染ACTC1-WT+miR-495-3p可极显著降低海肾荧光与萤火虫荧光的比值(P<0.01);与转染ACTC1-Mut+mimics NC相比,共转染ACTC1-Mut+miR-495-3p对海肾荧光比萤火虫荧光无显著影响(P>0.05)(图7),表明miR-495-3p可靶向ACTC1基因。

图7 miR-495-3p与ACTC1共转染双荧光素素酶活性检测结果Fig.7 Detection results of bifluorinase activity by co-transfection of miR-495-3p and ACTC1
3 讨 论
骨骼肌是动物生长发育的重要组成部分,对山羊产肉量和肉品质具有重要影响。骨骼肌的生长发育是极其复杂的生物学过程,包括细胞的增殖、分化、凋亡等,同时受到多种调节因子和信号通路的调控,miRNA作为重要的调控因子[11-12],在骨骼肌发育过程中发挥着重要作用。研究表明,miRNA在动物骨骼肌发育中的机制和作用研究大多采用高通量测序结合生物信息学分析的方法[13]。有研究报道,与肌肉发育相关的miRNA已经被鉴定出来,如miR-1靶向HDAC4基因促进肌肉发育[6],miR-486靶向Pax7基因促进肌肉发育[14]。然而,大部分与肌肉发育相关的miRNA尚未被鉴定,其作用机制和分子功能还有待于进一步研究。
研究报道,Lee等[15]通过高通量测序在小鼠成肌细胞和腓肠肌肌肉中筛选鉴定出miR-3074-3p,miR-3074-3p通过靶向Cav1促进成肌细胞分化。Keel等[16]利用RNA-Seq技术在牛背最长肌中鉴定出ACTC1基因,证明其可影响牛肌肉增重和采食量。本研究通过生物信息学分析预测确定miR-495-3p的靶基因为ACTC1,并通过构建1月龄隆林山羊miR-495-3p和ACTC1基因的组织表达谱发现,miR-495-3p及ACTC1在隆林山羊的背最长肌及腿肌中表达量均较高,符合特定组织和不同阶段表达模式[3],说明miR-495-3p和ACTC1基因均参与调控山羊肌肉发育。Boutilier等[17]研究结果表明,ACTC1基因是心脏α-肌动蛋白,是成熟心脏和胎儿骨骼肌中表达量最高的横纹肌α-肌动蛋白。以上研究结果表明,miRNA与肌肉生长发育密切相关。Goossens等[10]通过在体外培养的血管平滑肌细胞(VSMC)中过表达肌生长抑制素(myostatin,MYST),发现miR-495-3p的表达量下降,推测miR-495-3p与肌细胞增殖相关。Lee等[18]研究结果发现,过表达miR-146b-5p能促进鸡成肌细胞增殖相关基因的表达,但对肌管分化产生抑制作用,说明miR-146b-5p促进成肌细胞增殖,抑制其分化。Ye等[19]研究发现,miR-92b-3p能够靶向血清-糖皮质激素激酶3(serum-glucocorticoid-inducible kinase 3,SGK3)抑制C2C12细胞增殖并对C2C12细胞的迁移产生抑制作用,但是其对C2C12细胞分化并无显著影响。Qin等[20]从不同年龄组荣昌猪背最长肌中筛选出miR-323-3p,发现miR-323-3p可靶向白细胞抑制因子2(Smad2)抑制成肌细胞增殖并促进其分化。本研究通过将miR-495-3p模拟物和抑制物转染到山羊骨骼肌细胞中,发现miR-495-3p过表达可以促进MyoG、Myf5的表达,对增殖相关基因Pax7、CyclinE的表达无影响,抑制miR-495-3p表达时MyoG、Myf5的表达下降;最后通过双荧光素酶报告实验发现miR-495-3p靶向ACTC1基因,说明miR-495-3p通过靶向ACTC1基因促进骨骼肌细胞的分化。
4 结 论
本研究通过生物信息学的方法筛选出miR-495-3p的靶基因为ACTC1,miR-495-3p和ACTC1基因在1月龄隆林山羊心脏、肝脏、脾脏、肺脏、肾脏、背肌、腿肌中均有表达,在背最长肌中表达水平较高,具有组织特异性表达特点;miR-495-3p通过靶向ACTC1基因促进山羊骨骼肌细胞的分化,参与骨骼肌的生长发育,结果可为肉羊生产性能、肉质性状的分子选育研究提供参考。
猜你喜欢
中国畜牧杂志(2022年10期)2022-10-12
中国食物与营养(2022年5期)2022-06-17
中国食物与营养(2022年5期)2022-06-17
河南教育·职成教(2022年5期)2022-05-06
中国典型病例大全(2022年7期)2022-04-22
中国药房(2022年7期)2022-04-14
家庭医学·下半月(2020年2期)2020-04-26
幸福家庭(2019年12期)2019-01-07
现代仪器与医疗(2017年1期)2017-03-29
山东体育学院学报(2016年2期)2016-05-25
