
糠椴ATP合酶基因生物信息学分析
2022-05-31 05:19王玉文关乃千吴蒙嘉穆怀志
北华大学学报(自然科学版) 2022年1期
王玉文,关乃千,周 末,吴蒙嘉,孙 昆,穆怀志
(北华大学林学院,吉林 吉林 132013)
ATP合酶为多亚基复合体,广泛分布于质膜、线粒体内膜和类囊体膜,是光合作用中光合磷酸化和呼吸作用氧化磷酸化反应的关键酶[1].ATP合酶可分为3大类,分别为储能F-型、质膜P-型和液泡V-型,其中F-型ATP合酶(F1F0-ATP合酶)利用光合电子传递过程中产生的跨膜质子动力势催化ADP与无机磷合成ATP,实现光能向化学能的转化,产生细胞生命活动所需的ATP[2].因此,开展植物ATP合酶研究具有重要意义.
糠椴(Tiliamandshurica)又名辽椴,系椴树科(Tiliaceae)椴树属(Tiliaspp.)植物,广泛分布于我国东北、内蒙古、河北、山东、江苏北部,以及朝鲜、俄罗斯远东地区[3].糠椴树形通直、树冠高大圆满,不仅可以作绿化树种,同时也是重要的蜜源植物,具有重大的经济价值[4].近年来,关于糠椴的研究主要集中在种群分布格局[5-6]、解剖结构[7-9]、种子休眠[10-11]、无性繁殖[12-13]、光合特性[14-16]、化学成分[17-18]等方面,而关于糠椴ATP合酶的研究未见报道.本研究利用生物信息学方法分析糠椴ATP合酶基因,研究结果可为后续糠椴ATP合酶的基因克隆提供参考.
1 材料及方法
1.1 材 料
用NCBI(https:∥www.ncbi.nlm.nih.gov)已有的糠椴6个ATP合酶亚基基因序列,以及紫椴(Tiliaamurensis)、少脉椴(Tiliapaucicostata)、粉椴(Tiliaoliveri)、蝴蝶树(Heritieraparvifolia)、长柄银叶树(Heritieraangustata)、木棉(Bombaxceiba)、美丽梧桐(Firmianapulcherrima)、陆地棉(Gossypiumhirsutum)、梭罗树(Reevesiapubescens)、滇桐(Craigiayunnanensis)、面包树(Artocarpuscommunis)、黄秋葵(Abelmoschusesculentus)、木槿(Hibiscussyriacus)、梧桐(Firmianasimplex)、云南梧桐(Firmianamajor)、可可树(The-obromacacao)、朱槿(Hibiscusrosa-sinensis)、榴莲(Duriozibethinus)、施温迪茫棉(Gossypiumschwendim-anii)、海滨木槿(Taliparitihamabo)等ATP合酶的核苷酸序列和氨基酸序列作为试验材料.
1.2 方 法
分别利用ORF Finder、ProtParam、Protscale、TMHMMSERVER、PBIL、SWISS-MODE和MEGA7分析ATP合酶基因序列开放阅读框、基本性质、亲水性/疏水性、跨膜结构域、二级结构、同源建模、物种的同源进化树.
2 结果与分析
2.1 同源性
NCBI数据库中糠椴ATP合酶的序列号为NC_028589.1.糠椴ATP合酶由CF1和CF0两部分atpA、atpB、atpE、atpF、atpH、atpI共6个亚基组成.其中,CF1由atpA、atpB和atpE组成,CF0由atpF、atpH和atpI组成.BLAST分析结果表明:糠椴ATP合酶每个亚基基因与不同物种该基因的同源性并不相同.与糠椴atpA亚基基因相似度达到100%的物种为紫椴、少脉椴、粉椴、蝴蝶树、长柄银叶树、木棉、美丽梧桐、陆地棉和梭罗树;与糠椴atpB亚基基因相似度达到100%的物种为紫椴、少脉椴、粉椴、木棉、滇桐、梭罗树、梧桐、蝴蝶树和面包树;与糠椴atpE亚基基因相似度达到100%的物种为紫椴、少脉椴、粉椴、美丽梧桐、梧桐、施温迪茫棉、蝴蝶树、黄秋葵和木槿;与糠椴atpF亚基基因相似度达到100%的物种为紫椴、粉椴、少脉椴、梭罗树、陆地棉、云南梧桐、蝴蝶树、美丽梧桐和可可树;与糠椴atpH亚基基因相似度达到100%的物种为朱槿、紫椴、粉椴、梭罗树、木槿、施温迪茫棉、蝴蝶树、木棉和榴莲;与糠椴atpI亚基基因相似度达到100%的物种为紫椴、少脉椴、粉椴、梭罗树、蝴蝶树、木棉、榴莲、可可树和海滨木槿.与糠椴ATP合酶各亚基基因序列相似度达到100%的物种见表1.

表1 与糠椴ATP合酶各亚基基因序列相似度达到100%的物种Tab.1 Species with 100% similarity of ATP synthase subunit gene sequences in Tilia mandshurica
2.2 开放阅读框
利用ORF Finder分析糠椴ATP合酶不同亚基序列的开放阅读框,结果见表2.由表2可见:6个亚基的序列长度差异较大,其中,atpA的基因序列最长,达到1 523 bp;其次为atpB(1 496 bp)和atpF(1 355 bp);atpH的基因序列最短,仅为245 bp.

表2 糠椴ATP合酶亚基基因位置及长度Tab.2 Location and length of ATP synthase subunits in Tilia mandshurica /bp
2.3 氨基酸组成
利用ProtParam分析糠椴ATP合酶各亚基的氨基酸组成、相对分子质量和理论等电点,结果见图1、表3.结果表明:atpA亚基含有19种氨基酸,其中,丙氨酸含量最高(9.86%),半胱氨酸含量最低(0.20%),分子式为C2442H3974N678O764S12,相对分子质量为55 441.22,等电点为5.26;atpB亚基含有19种氨基酸,其中,亮氨酸含量最高(10.04%),半胱氨酸含量最低(0.20%),分子式为C2382H3680N648O728S16,相对分子质量为53 737.72,等电点为5.29;atpE亚基含有19种氨基酸,其中,丙氨酸和异亮氨酸含量最高,均为12.03%,半胱氨酸、组氨酸、苯丙氨酸含量最低,均为0.75%,分子式为C633H1059N189O195S4,相对分子质量为14 565.76,等电点为5.83;atpF亚基含有19种氨基酸,其中,亮氨酸含量最高(12.50%),色氨酸含量最低(0.54%),分子式为C915H1481N265O283S3,相对分子质量为20 818.61,等电点为6.33;atpH亚基含有16种氨基酸,其中,丙氨酸含量最高(19.75%),酪氨酸和赖氨酸含量最低,均为1.24%,分子式为C264H599N93O103S2,相对分子质量为7 990.44,等电点为4.94;atpI亚基含有20种氨基酸,其中,亮氨酸含量最高(11.74%),半胱氨酸含量最低(0.41%),分子式为C1273H1947N297O339S5,相对分子质量为26 996.95,等电点为5.06.

A.丙氨酸;R.精氨酸;N.天冬酰胺;D.天冬氨酸;C.半胱氨酸;Q.谷氨酰胺;E.谷氨酸;G.甘氨酸;H.组氨酸;I.异亮氨酸;L.亮氨酸;K.赖氨酸;M.甲硫氨酸;F.苯丙氨酸;P.脯氨酸;S.丝氨酸;T.苏氨酸;W.色氨酸;Y.酪氨酸;V.缬氨酸.图1 糠椴ATP合酶亚基的氨基酸组成Fig.1 Amino acid composition of ATP synthase subunits in Tilia mandshurica

表3 糠椴ATP合酶亚基氨基酸数量、相对分子质量和理论等电点Tab.3 Amino acid quantity,molecular weight and theoretical isoelectric point of ATP synthase subunit genes in Tilia mandshurica
2.4 蛋白质亲/疏水性
糠椴ATP合酶的亲/疏水性分析结果见图2.结果表明:atpA、atpB、atpE和atpF为亲水蛋白,atpH和atpI为疏水蛋白.atpA的GRAVY值为-0.102,其最大值(2.7)和最小值(-2.8)分别出现在190和20位点;atpB的GRAVY值为-0.038,最大值(2.3)出现在260位点,最小值(-2.7)出现在210位点;atpE的GRAVY值为-0.044,最大值(1.9)出现在40位点,最小值(-2.6)出现在107位点;atpF的GRAVY值为-0.352,最大值(3.3)出现在40位点,最小值(-2.5)出现在120位点;atpH的GRAVY值为1.002,最大值(3.3)出现在72位点,最小值(-2.0)出现在43位点;atpI的GRAVY值为0.668,最大值(3.6)出现在205位点,最小值(-1.8)出现在60位点.

图2 糠椴ATP合酶各亚基的亲/疏水性Fig.2 Hydrophobic and hydrophilic properties of ATP synthase subunits in Tilia mandshurica
2.5 跨膜结构域
TMHMM分析显示,糠椴ATP合酶不同亚基的跨膜结构不尽相同.其中,atpI的跨膜区最多,高达5个;atpH和atpF的跨膜区分别为2个和1个;atpA、atpB和atpE不存在跨膜结构(图3).

图3 糠椴ATP合酶各亚基的跨膜结构域Fig.3 Transmembrane domain of ATP synthase subunits in Tilia mandshurica
2.6 二级结构与同源建模
利用PBIL分析糠椴ATP合酶的二级结构,结果见表4.由表4可知:不同亚基的二级结构组成不同,其中,atpA、atpB、atpE和atpI的二级结构由α螺旋、β折叠和无规则卷曲组成,而atpF和atpH仅由α螺旋和无规则卷曲组成,无β折叠.α螺旋比例较高的是atpH和atpF,分别达到86.42%和86.41%;其次是atpI,为71.26%;atpE最低,仅为29.32%.β折叠比例最高的是atpE,高达37.59%;其次是atpB和atpA,分别为18.27%和14.00%;atpI最低,仅为1.62%.无规则卷曲比例最高的是atpB,高达43.98%;其次是atpA,为43.40%;atpH最低,仅为13.58%.利用SWISS-MODEL对糠椴ATP合酶各亚基的氨基酸序列进行同源建模,得到的三维结构见图4.
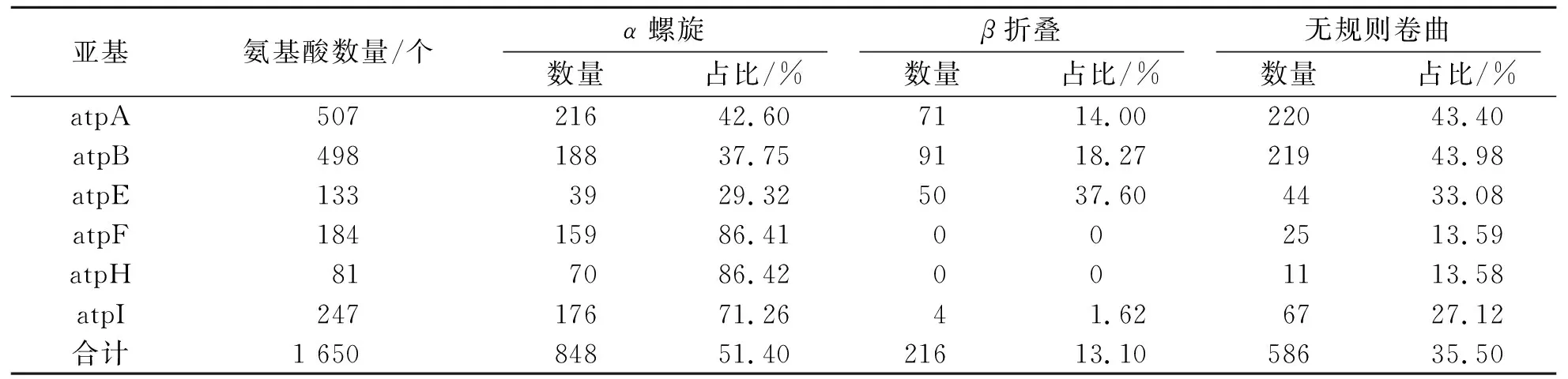
表4 糠椴ATP合酶亚基二级结构数量及比例Tab.4 Quantity and proportion of secondary structure of ATP synthase subunits in Tilia mandshurica

图4 糠椴ATP合酶及各亚基三维结构Fig.4 Three-dimensional structure of ATP synthase and its subunits in Tilia mandshurica
2.7 系统进化树
利用MEGA7分别构建糠椴ATP合酶6个亚基与紫椴、少脉椴、粉椴、美丽梧桐、梧桐、云南梧桐、蝴蝶树、长柄银叶树、陆地棉、木棉、梭罗树、滇桐、面包树、黄秋葵、施温迪茫棉、木槿、海滨木槿、朱瑾、榴莲和可可树的进化树,10个亲缘关系较近树种构建的进化树见图5.由图5可见:不同亚基对应的进化树结构并不相同.糠椴atpA、atpB、atpE、atpF和atpI亚基基因与粉椴、少脉椴和紫椴亲缘关系较近;糠椴atpH亚基基因与木槿亲缘关系较近,而与紫椴和粉椴的亲缘关系相对较远.
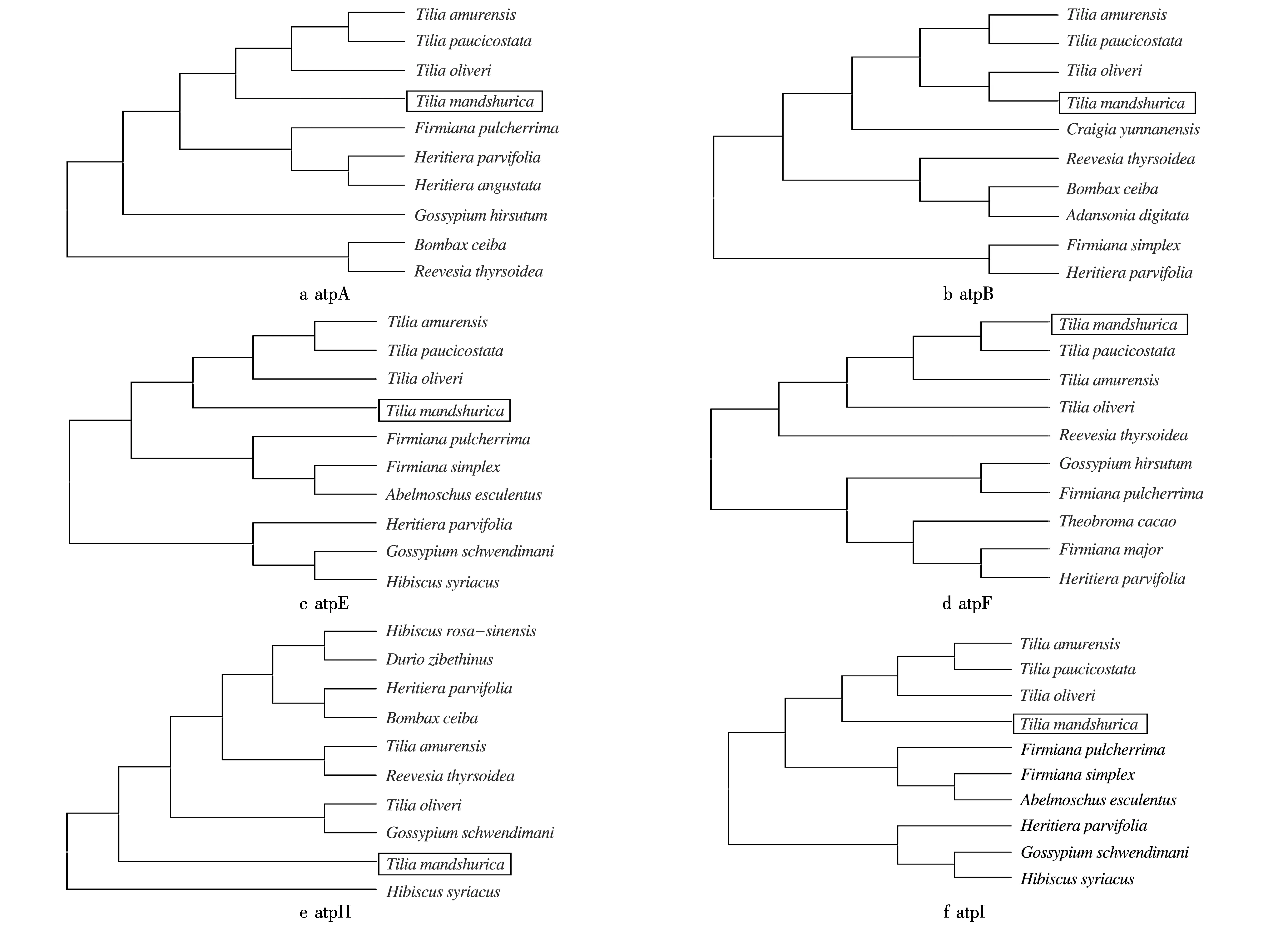
图5 糠椴ATP合酶不同亚基基因系统进化树Fig.5 Phylogenetic tree of ATP synthase subunit genes in Tilia mandshurica
3 结论与讨论
ATP合酶广泛分布于线粒体内膜、叶绿体类囊体、异养菌和光合菌的质膜上,参与氧化磷酸化和光合磷酸化,是生物能量转换的关键酶,影响细胞生命活动所需ATP的产生[1,19].目前,已有很多有关ATP合酶基因的报道.林莉莉等[20]对ATP合酶的C1F1F0-ATP 基因进行红蓝光不同光质配比处理,发现白光促进杉木幼苗生长,说明C1F1F0-ATP 基因对白光的响应与杉木木材生长发育的调控有着密切关系.对砂藓ATP合酶Ⅱ亚基的基因表达研究表明,该亚基在脱水胁迫和复水过程中的表达发生了明显变化,说明ATP合酶Ⅱ亚基可能参与了砂藓干旱胁迫的应答[21].MAO等[22]将水稻幼苗在低温下处理不同时间,Northern杂交分析显示其atpH亚基基因转录水平在低温处理1 d时明显下降,低温处理2 d后完全被抑制.李生广等[23]研究发现,乙醇对ATP合酶表现为非竞争性抑制,说明乙醇并未直接与反应底物竞争结合位点,而是作用于ATP合酶上的乙醇敏感部位,导致CF1构象发生改变,从而使CF1上催化位点与调节位点解聚,进而影响活性部位,最终造成ATP水解受到抑制.姜晓旭等[24]从蒙古沙冬青幼苗中克隆得到ATP合酶的atpD亚基基因,发现该基因在低温和干旱胁迫下的表达量明显升高,说明该基因可受低温和干旱胁迫诱导表达.
本研究表明,糠椴ATP合酶共有6个亚基,分别为atpA、atpB、atpE、atpF、atpH和atpI.糠椴ATP合酶由51.40%的α螺旋、13.10%的β折叠和35.50%的无规则卷曲组成.在6个亚基中,atpA、atpB、atpE和atpF为亲水性蛋白;atpH和atpI为疏水蛋白.atpI有5个跨膜区,atpH和atpF分别有2个和1个跨膜区,atpA、atpB和atpE不存在跨膜结构.综上所述,糠椴ATP合酶是一个镶嵌在膜上且具有跨膜结构的酶,既有亲水性部分,又有疏水性部分,且各亚基具有明显差异.上述结果可为深入了解糠椴ATP合酶的结构以及通过基因克隆和转基因技术验证其功能提供参考.
猜你喜欢
分子催化(2022年1期)2022-11-02
分析测试学报(2022年9期)2022-09-21
中国农业科学(2022年16期)2022-09-19
新疆农业科学(2022年6期)2022-07-13
麦类作物学报(2022年3期)2022-05-19
临床与实验病理学杂志(2022年3期)2022-04-06
作文大王·低年级(2022年3期)2022-03-19
电脑知识与技术(2018年19期)2018-11-01
学生天地·小学低年级版(2018年7期)2018-10-11
传奇故事(破茧成蝶)(2015年6期)2015-02-28
