
姜黄素对人肝癌SMMC-7721细胞裸鼠移植瘤的抑制作用及其机制
2022-05-31 04:24孙俊徐伟陆荣柱朱孝仁徐新明蒋一全陈健
江苏大学学报(医学版) 2022年3期
孙俊, 徐伟, 陆荣柱, 朱孝仁, 徐新明, 蒋一全, 陈健
(1. 江苏大学附属昆山医院肝胆外科,江苏 苏州 215300; 2. 江苏大学医学院,江苏 镇江 212013; 3. 南京中医药大学附属盐城市中医院病理科,江苏 盐城 224001; 4. 江苏大学附属昆山医院中心实验室,江苏 苏州 215300)
内质网是细胞内蛋白折叠及翻译后加工、脂质合成等重要场所,对维持细胞稳态极为重要;当发生缺氧、pH变化等情况时,大量错误折叠或未折叠的蛋白质蓄积于内质网内,破坏内质网稳态,诱发内质网应激(endoplasmic reticulum stress,ERS)[1-2]。ERS主要有如下3种类型:未折叠蛋白质反应、内质网的超负荷反应、调节固醇元件的结合蛋白激活的固醇级联反应;其中,未折叠蛋白反应最为多见,故一般情况下ERS指代的就是未折叠蛋白质反应。内质网膜上有3种重要的跨膜传感器蛋白,分别为需肌醇酶1(inositol requiring enzyme1,IRE1)、蛋白激酶RNA样内质网激酶(protein kinase RNA-like ER kinase,PERK)和激活转录因子6(activating transcription factor 6,ATF6)。当发生ERS时,3种蛋白分别与葡萄糖调节蛋白78(glucose regulated protein 78, GRP78)解离,识别并结合内质网中积累的未折叠或错误折叠蛋白,以对抗应激,维持内质网稳态[2]。当内质网稳态无法维持时,ERS则会诱导细胞凋亡。
原发性肝癌的发病率及死亡率高[3],目前化疗药物的治疗效果不理想,且不良反应较大。姜黄素是一种从姜黄的根茎中提取出的天然植物活性成分,能够明显抑制乳腺癌[4]、前列腺癌[5]、人胶质母细胞瘤[6]和胰腺癌[7]等肿瘤细胞增殖并促进其凋亡。本课题组前期实验表明,姜黄素在体外可以通过激活ERS通路,抑制肝癌细胞增殖并促进其凋亡[8]。但是,姜黄素对人肝癌SMMC-7721细胞裸鼠移植瘤的效应及其潜在机制仍不明确。因此,本研究拟建立裸鼠皮下肝癌移植瘤模型,检测瘤体组织中ERS相关蛋白及凋亡相关蛋白的表达情况,并评价姜黄素在体内对肝癌的抑制效应。
1 材料与方法
1.1 动物、细胞及主要试剂
SPF级BALB/c雌性裸鼠,4周龄,20只,购自上海斯莱克实验动物有限责任公司,许可证号为SCXK(沪)2017-0005;人肝癌SMMC-7721细胞株购自湖南丰晖生物科技有限公司;优质胎牛血清购自上海双洳科技生物有限公司;胰蛋白酶购自美国Gibco公司;RPMI 1640培养基购自美国HyClone公司;姜黄素购自美国Sigma公司;兔抗鼠GRP78、IRE1和β-微管蛋白抗体购自美国Abcam公司;兔抗鼠磷酸化PERK(p-PERK)抗体购自北京博奥森生物公司;兔抗鼠磷酸化真核细胞起始因子2α(phospho-eukaryotic initiation factor 2α,p-eIF2α)、Bcl-2、Cleaved Caspase-3抗体购自美国Cell Signaling Technology公司;兔抗鼠C/EBP同源蛋白(C/EBP homologous protein,CHOP)抗体购自杭州华安生物技术有限公司;HRP标记的羊抗兔IgG抗体购于美国Santa Cruz公司;RIPA裂解液、BCA蛋白浓度测定试剂盒、分离胶配制试剂盒均为上海碧云天生物公司产品;DAB染色液、苏木素染色剂、免疫组化抗原修复试剂盒均为上海桂浦生物科技有限公司产品。
1.2 细胞培养与传代
将人肝癌SMMC-7721细胞株用含10%优质胎牛血清及1%双抗(青霉素和链霉素)的RPMI 1640培养基,培养于37 ℃、5% CO2细胞培养箱中。传代次数在10代以内。
1.3 裸鼠皮下肝癌移植瘤模型的建立
取传代后处于对数生长期的人肝癌SMMC-7721细胞,胰蛋白酶消化后离心,用PBS重悬,定量细胞密度至2×107个/mL。取100 μL肝癌细胞(约2×106个)接种于裸鼠右侧腋下,继续在SPF环境中饲养至实验结束。接种第5天,所有受试裸鼠皮下均可见一个明显、清晰的结节,其边界清晰,质地较硬,可推动。由此可证实人肝癌SMMC-7721细胞裸鼠移植瘤模型构建成功。
1.4 动物分组及处理
待裸鼠皮下肿瘤组织最长径均大于5 mm后,将20只成瘤裸鼠随机均分为4组,即对照组、10 mg/kg姜黄素组、50 mg/kg姜黄素组、100 mg/kg姜黄素组,每组5只。均采用腹腔注射给药,分别给予生理盐水及对应剂量姜黄素,每日给药1次,每次200 μL,连续16 d。
1.5 指标测量
自给药当天起,每隔一日测量各组裸鼠体重及裸鼠皮下移植瘤的最长径a(mm)和最短径b(mm),采用公式V(mm3)=ab2/2计算皮下移植瘤体积。同时观察各裸鼠的神经行为及营养状况。给药结束后第2天,给予所有裸鼠2%水合氯醛麻醉,完整剥离肿瘤组织,PBS清洗,滤纸吸干水分后测量瘤体重量,并计算抑瘤率。抑瘤率(%)=(对照组平均瘤重-姜黄素组平均瘤重)/对照组平均瘤重×100%。
1.6 蛋白质免疫印迹法检测瘤体组织中ERS通路相关蛋白及凋亡蛋白表达
称取各组瘤体组织,每20 mg组织加入150 μL RIPA裂解液,研磨、离心后提取总蛋白。用BCA法测定蛋白浓度。配制8% SDS-PAGE分离胶,80 V电泳分离蛋白90 min;300 mA转膜90 min至PVDF膜;5%奶粉室温封闭2 h;将PVDF膜加入对应的一抗稀释液中(GRP78、IRE1、p-PERK、p-eIF2α、CHOP、Cleaved Caspase-3及Bcl-2抗体稀释比均为1 ∶1 000;内参为GAPDH或β-微管蛋白,稀释比为1 ∶1 000);TBST洗涤3次;加入二抗稀释液(1 ∶10 000),室温孵育1 h;TBST洗涤;膜上滴加荧光试剂ECL,经凝胶成像系统显影。取目的条带灰度值与内参条带灰度值的比值作为目的蛋白相对表达量。
1.7 免疫组织化学法检测瘤体组织中GRP78和IRE1蛋白表达
取出液氮中瘤体组织,切取部分组织标本,用10%甲醛液固定脱水,常规石蜡包埋,制备4 μm连续切片。60 ℃烘烤1 h;经二甲苯脱蜡3次,每次10 min;再经从高到低浓度梯度的乙醇浸泡、水化,每次5 min;将pH=6.0抗原修复试剂置于微波炉中高火加热5~6 min,放入载玻片后转中高火修复15 min;将配好的3%过氧化氢滴加于切片,室温孵育15 min;PBS清洗3次;滴加山羊血清后室温封闭30 min弃去上清液;分别加入GRP78和IRE1一抗稀释液(1 ∶500),4 ℃孵育过夜;加二抗稀释液(1 ∶200),孵育30 min;清洗5 min,甩干;加入DAB溶液显色,双蒸水冲洗,苏木素染色液复染10 s;脱水、透明后中性树脂封片。
在普通光学显微镜(×400)下,随机选取5个不同视野,计数细胞总数及阳性细胞数,按阳性细胞所占的百分比计分。评分方法如下:① 按阳性细胞率计分:阳性率≤10%计1分,10%~50%计2分,50%~75%计3分,≥75%计4分;② 按染色程度评分:阴性1分,弱染色2分,中等强度染色3分,强染色4分。由两位病理科医师在双盲情况下观察记分;将两种方法计得分数的乘积用来判断结果,即:阴性(-)为≤4分,弱阳性(+)为5~8分,阳性(++)为9~12分,强阳性(+++)为>12分[9]。
1.8 数据分析
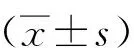
2 结果
2.1 姜黄素处理对荷瘤裸鼠行为及体重的影响
在整个给药期间,不同剂量姜黄素组受试裸鼠行为未发现异常,营养状态良好;4组裸鼠体重均呈逐渐增长趋势。对照组裸鼠体重增加略高于各姜黄素处理组,但各组裸鼠体重增长并没有明显差异(P>0.05)。见图1。
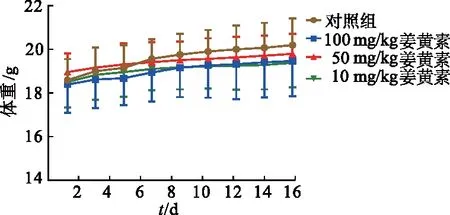
图1 各组裸鼠体重变化比较
2.2 姜黄素对裸鼠移植瘤体积的影响
由图2可见,随着给药时间延长,各组裸鼠瘤体体积均呈增大趋势。但第10天,100 mg/kg姜黄素组瘤体体积明显小于对照组及10 mg/kg姜黄素组(P均<0.05);自第12天起,50 mg/kg和100 mg/kg姜黄素组瘤体体积均明显小于对照组及10 mg/kg姜黄素组(P均<0.05),且100 mg/kg姜黄素组明显小于50 mg/kg姜黄素组(P均<0.05)。
a:P<0.05,与对照组比较;b:P<0.05,与10 mg/kg姜黄素组比较;c:P<0.05,与50 mg/kg姜黄素组比较
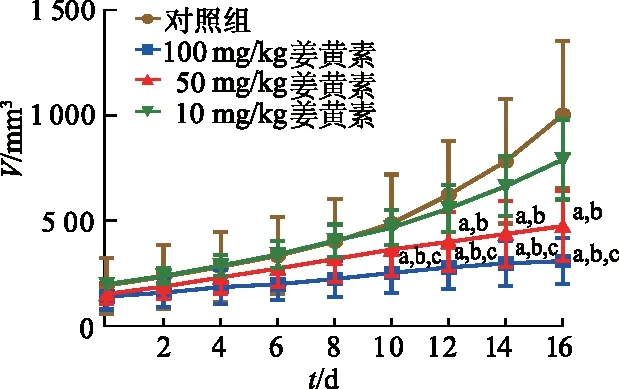
2.3 姜黄素对瘤体重量及抑瘤率的影响
随着姜黄素剂量增加,各组中瘤重逐渐降低,100 mg/kg姜黄素组瘤重较50、10 mg/kg姜黄素组分别降低35.4%、16.5%。与对照组相比,10、50和100 mg/kg姜黄素组裸鼠移植瘤瘤重明显降低(q=4.067、5.258、6.289,P均<0.05)。10、50和100 mg/kg姜黄素组抑瘤率分别为35.3%、45.7%、54.7%。见图3。
a:P<0.05,与对照组比较
2.4 姜黄素对移植瘤中ERS通路相关蛋白及凋亡相关蛋白表达的影响
2.4.1 姜黄素对移植瘤中ERS通路标志性蛋白GRP78表达的影响 与对照组相比,姜黄素组GRP78蛋白表达均出现增加趋势,且50和100 mg/kg姜黄素组GRP78表达量明显高于对照组(q=6.426、11.190,P均<0.05);100 mg/kg姜黄素组GRP78表达量显著高于10和50 mg/kg姜黄素组(q=7.799、6.769,P均<0.05)。见图4。
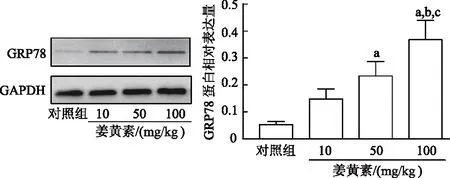
a:P<0.05,与对照组比较;b:P<0.05,与10 mg/kg姜黄素组比较;c:P<0.05,与50 mg/kg姜黄素组比较
2.4.2 姜黄素对移植瘤中未折叠蛋白反应下游通路蛋白表达的影响 与对照组相比,姜黄素组中p-PERK和p-eIF2α表达量均显著增加(P均<0.05),且各剂量组间p-PERK蛋白表达差异均有统计学意义(P<0.05),呈一定的剂量依赖性。与对照组及10 mg/kg姜黄素组相比,IRE1在50、100 mg/kg姜黄素组中表达明显增加(P均<0.05),且100 mg/kg姜黄素组明显高于50 mg/kg姜黄素组(q=6.713,P<0.05)。CHOP在50、100 mg/kg姜黄素组中表达水平较对照组明显增加(q=5.473、10.160,P均<0.05),且100 mg/kg姜黄素处组明显高于10、50 mg/kg姜黄素组(q=6.822、4.688,P均<0.05),10、50 mg/kg姜黄素组间差异无统计学意义(P>0.05)。见图5。
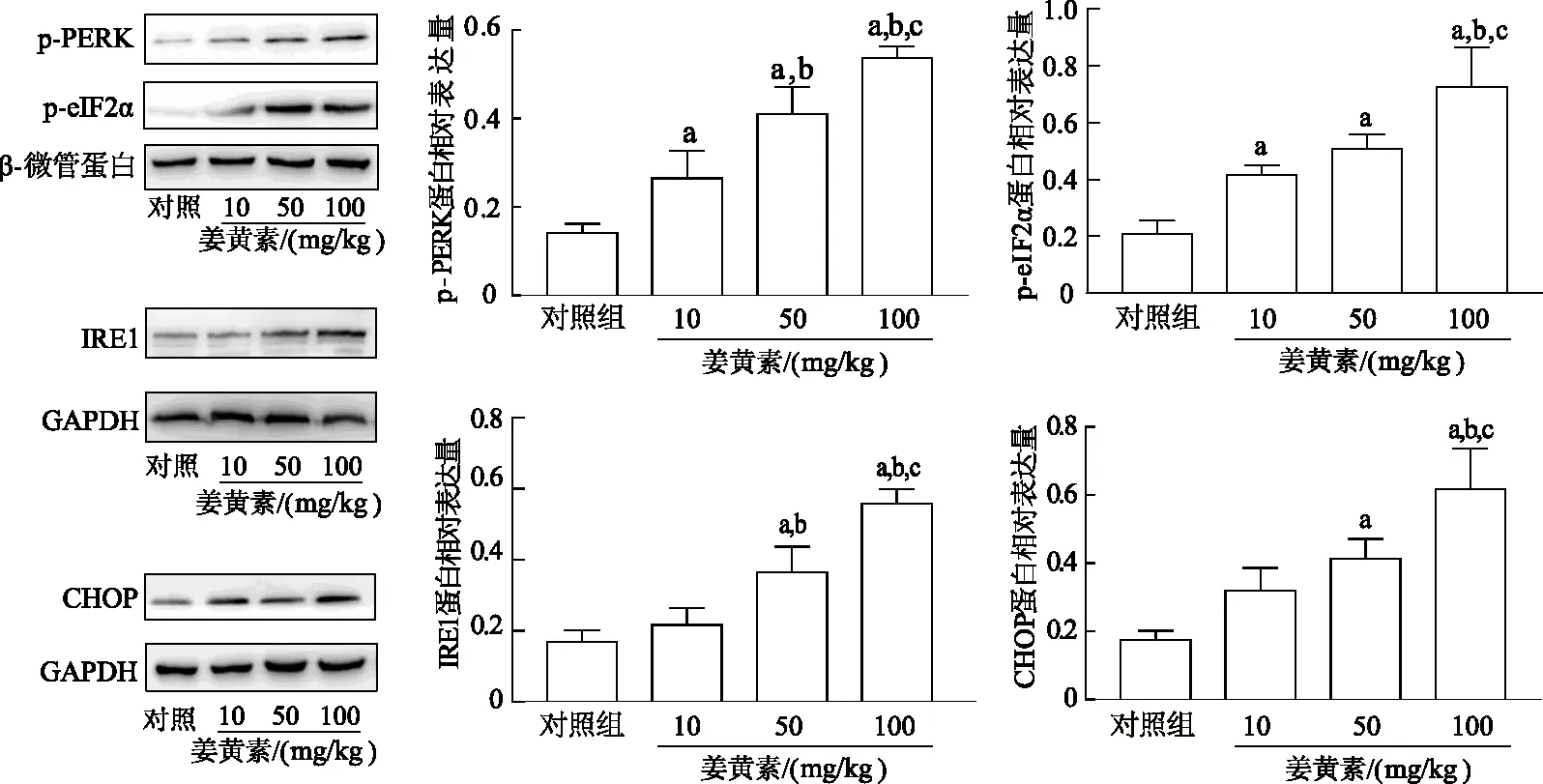
a:P<0.05,与对照组比较;b:P<0.05,与10 mg/kg姜黄素组比较;c:P<0.05,与50 mg/kg姜黄素组比较
2.4.3 姜黄素对移植瘤凋亡相关蛋白表达的影响 免疫印迹结果显示,与对照组比较,10、50和100 mg/kg姜黄素组中抗凋亡蛋白Bcl-2表达明显减少(P均<0.05),50和100 mg/kg姜黄素组凋亡蛋白Cleaved Caspase-3表达明显增加(q=5.631、10.160,P均<0.05);与10 mg/kg姜黄素组比较,50和100 mg/kg姜黄素组中Cleaved Caspase-3表达亦明显增加(q=4.850、9.375,P均<0.05)。见图6。

a:P<0.05,与对照组比较;b:P<0.05,与10 mg/kg姜黄素组比较
2.5 姜黄素对瘤体组织中GRP78和IRE1表达的影响
免疫组化染色结果显示,与对照组比较,10、50、100 mg/kg姜黄素组中GRP78和IRE1蛋白阳性表达评分明显增加(P均<0.05),且各剂量组间GRP78和IRE1阳性表达评分差异均具有统计学意义(P均<0.05),呈一定的剂量依赖性。见图7。

a:P<0.05,与对照组比较;b:P<0.05,与10 mg/kg姜黄素组比较;c:P<0.05,与50 mg/kg姜黄素组比较
3 讨论
本研究采用人肝癌SMMC-7721细胞裸鼠皮下移植瘤作为研究模型,实验中所有裸鼠均植瘤成功,成瘤率为100%。此外,随着用药时间延长,所有组别裸鼠体重都呈缓慢上升趋势,其体重增加无明显差异;未发现各组裸鼠有消瘦或生长活动异常情况。由此表明,在一定时间内姜黄素用药对裸鼠自身生长无明显影响,可能是一种相对安全的药物。
本研究发现,10、50、100 mg/kg姜黄素组移植瘤体积和瘤重均明显小于对照组,表明姜黄素能有效抑制裸鼠肝癌皮下移植瘤生长。10 mg/kg姜黄素组移植瘤体积与对照组比较差异无统计学意义,可能是姜黄素剂量过低,导致其对肿瘤抑制作用不如50、100 mg/kg姜黄素组,但10 mg/kg姜黄素组移植瘤体积增长幅度仍小于对照组。本研究结果与戴莉等[10]相一致,由此表明,就瘤重而言,所用姜黄素剂量越高,抑瘤效果越明显。
ERS激活是诱导肿瘤细胞凋亡的重要机制之一[11-12]。大量研究表明姜黄素可能是通过激活ERS通路进而破坏肿瘤细胞的内质网稳态达到抗癌目的[13-15]。Zhang等[16]发现姜黄素通过破坏细胞内Ca2+稳态来激发ERS诱导人乳头状甲状腺癌BCPAP细胞凋亡。本课题组前期研究也证实姜黄素能在体外通过激活ERS来诱导人肝癌SMMC-7721细胞凋亡[8]。本研究发现,50和100 mg/kg姜黄素处理组ERS标志性蛋白GRP78蛋白表达明显上调,而在10 mg/kg姜黄素组中该蛋白表达较对照组有升高趋势,但是差异无统计学意义,可能提示药物剂量较低,导致ERS虽已激活,但作用不强烈,从而对肿瘤抑制效果不明显。Yang等[17]研究表明,虽然25 mg/kg姜黄素抗前列腺肿瘤效果与对照组相比有差异,但是其抗癌效果不如50、100 mg/kg姜黄素组,究其原因可能是较低剂量的姜黄素不足以刺激肿瘤细胞产生过于持续而强烈的ERS。
当发生ERS时,PERK与GRP78解离后迅速自磷酸化,磷酸化的PERK具有更强的活性。本实验中p-PERK在3个姜黄素组中表达均较对照组明显上调,且呈一定的剂量依赖性。p-PERK介导eIF2α磷酸化,p-eIF2α活性远大于eIF2α且相对更容易检测。在刘沙等[18]研究中,重组新城疫病毒在抑制肺癌移植瘤增长的同时,致瘤体组织内p-eIF2α表达明显增加,证实裸鼠肿瘤细胞内发生ERS。本实验中p-eIF2α在各姜黄素组中表达均较对照组明显增加,其中50 mg/kg姜黄素组表达高于10 mg/kg姜黄素组,但差异无统计学意义,可能由于50 mg/kg姜黄素对活性更强的p-eIF2α蛋白激活效应仍不够强烈。未折叠蛋白反应另一感受器蛋白IRE1在10 mg/kg姜黄素组中的表达与对照组比较差异无统计学意义,表明该剂量无法刺激细胞产生过强的ERS。相对IRE1通路而言,PERK通路可能对低剂量的姜黄素更敏感。在郭立达等[19]研究中,单独使用50 mg/kg姜黄素能较好地抑制结肠癌裸鼠移植瘤生长;同时还发现,姜黄素和奥沙利铂联用可协同抗癌,抑瘤效果更好。在本实验中,ERS通路相关蛋白在50 mg/kg姜黄素组中表达量明显高于对照组,提示50 mg/kg可能为姜黄素有效抗癌剂量。最后免疫组化结果辅助证实ERS相关蛋白GRP78和IRE1阳性表达随着姜黄素剂量增大而增加,与蛋白免疫印迹结果基本一致,提示ERS抑制肿瘤生长。
Caspase-3是Caspase级联反应中最关键的细胞凋亡执行因子,而Bcl-2作为Caspase-3上游的调控因子,在细胞中具有明显的抗凋亡作用[20-22]。本实验结果显示,经过姜黄素作用后Bcl-2表达较对照组明显降低,虽然3个姜黄素组间表达并无明显差异,但是随着姜黄素剂量增大,Bcl-2表达量呈下降趋势。后续实验中可测定Bax表达水平,计算Bcl-2与Bax表达水平的比值,以便更好地阐释在姜黄素作用下Bcl-2对细胞凋亡的调控[19]。此外,本研究结果显示,50、100 mg/kg姜黄素组中Cleaved Caspase-3表达明显高于对照组和10 mg/kg姜黄素组,提示在50、100 mg/kg剂量时,姜黄素促进肿瘤细胞凋亡的作用明显,且呈一定的剂量依赖性。CHOP是未折叠蛋白反应中介导细胞凋亡的关键蛋白,且其与未折叠蛋白反应的3个通路均有关[23]。在姜黄素作用下,CHOP信号通路被激活,同时Bcl-2表达下调引发线粒体外膜通透性改变,继而释放细胞色素C,进而释放凋亡小体,活化Caspase-9,产生级联反应,最终活化Caspase-3,诱导肿瘤细胞凋亡[24-26]。本实验发现,50、100 mg/kg姜黄素组中CHOP表达明显高于对照组及10 mg/kg姜黄素组,而10 mg/kg姜黄素组CHOP表达虽较对照组有上升趋势,但是差异无统计学意义;由此提示,当姜黄素剂量大于50 mg/kg时,可激活ERS的关键蛋白CHOP,进而促进肝癌细胞凋亡,这也印证了前文所述的推测,即50 mg/kg可能是姜黄素的最低抗癌有效剂量。此外,敲除或过表达ERS相关基因后姜黄素在体内抗肝癌的效果有待进一步探索。同时,姜黄素与临床上常用的其他与ERS相关抗肝癌化疗药物联合应用的抗癌效果亦有待研究。
综上所述,本研究结果表明,姜黄素可抑制人肝癌SMMC-7721细胞裸鼠移植瘤生长,其机制可能与激活ERS通路有关,进而通过共同的CHOP途径抑制下游的抗凋亡蛋白Bcl-2、增强凋亡蛋白Cleaved Caspase-3表达来抑制肝癌细胞增殖,促进其凋亡。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国医学物理学杂志(2022年9期)2022-10-09
中国现代医生(2022年19期)2022-08-25
中国药房(2022年9期)2022-05-17
家庭百事通·健康一点通(2021年1期)2021-02-24
现代养生·上半月(2021年1期)2021-01-28
科教导刊·电子版(2018年25期)2018-10-31
文萃报·周五版(2018年25期)2018-08-13
文萃报·周二版(2018年26期)2018-08-04
新课程·中学(2018年10期)2018-02-19
