
基于植物功能-结构模型的玉米-大豆条带间作光截获行间差异研究
2022-05-30 22:38:33李双伟朱俊奇JochemB.EVERSWopkeVANDERWERF郭焱李保国马韫韬
智慧农业(中英文) 2022年1期
李双伟 朱俊奇 Jochem B. EVERS Wopke VAN DER WERF 郭焱 李保国 马韫韬


摘要:间作种植形成了异质冠层空间结构,但因此导致的作物生长、表型和光截获的行间差异目前还少有定量化。为解析条带间作生产力的行间差异,本研究基于田间观测数据构建植物功能-结构模型( Func? tional-Structural Plant Model ,FSPM ),量化间作系统中光截获的行间差异。于2017—2018年开展了玉米和大豆单作、2行玉米和2行大豆的2:2 MS 间作以及3行玉米和6行大豆的3:6 MS 间作田间试验。基于植物生长平台GroIMP开发了玉米-大豆间作的 FSPM ,模型较好地模拟了叶面积指数(Leaf Area Index ,LAI )、株高和光截获系数动态三个指标,均方根误差(Root Mean Square Error ,RMSE )分别为0.24~0.70 m2/m2、0.06~0.17 m 和0.06~0.10。田间试验结果表明,间作种植显著增加了玉米节间直径。受玉米遮阴影响,大豆节间变长、变细,且随大豆条带变窄差异越明显。模型模拟的2:2 MS 间作玉米光截获比单作玉米高35.6%,3:6 MS 边行玉米和内行玉米的光截获分别比单作玉米高27.8%和20.3%。2:2 MS 与3:6 MS 边行大豆的光截获比单作大豆分别少36.0%和28.8%;3:6 MS 大豆内 I 行和内 II 行比单作大豆的光截获分别少4.1%和1.8%。基于三维 FSPM ,未来可进行不同生长环境下间作种植模式等的布局优化,以达到最佳系统光截获优势。
关键词:玉米-大豆间作;植物功能-结构模型;光截获;三维结构;表型可塑性;行间差异
中图分类号:S565.1;S344.2 文獻标志码:A 文章编号:SA202202002
引用格式:李双伟, 朱俊奇, EVERS Jochem B., VAN DER WERF Wopke, 郭焱, 李保国, 马韫韬.基于植物功能-结构模型的玉米-大豆条带间作光截获行间差异研究[J].智慧农业(中英文), 2022, 4(1):97-109.
LI Shuangwei, ZHU Junqi, EVERS Jochem B., VAN DER WERF Wopke, GUO Yan, LI Baoguo, MA Yuntao. Esti‐ mating the differences of light capture between rows based on functional-structural plant model in simultaneous maize-soybean strip intercropping[J]. Smart Agriculture, 2022, 4(1):97-109.(in Chinese with English abstract)
1 引言
玉米-大豆条带间作是中国北方和西南地区普遍应用的一种重要种植方式。间作系统中不同形态结构的作物条带种植,增加了物种多样性,提高了系统生产力。间作系统的产量增加可通过种内和种间相互关系进行解释[1]。玉米-大豆条带间作时,植物根据所处环境调节自身生长,改善并提高了系统内植物冠层的光分布,研究表明条带边行的种间相互作用起到了决定作用[2,3]。一般来说,优势作物玉米的边行可截获更多的光,产量明显高于内行,而从属作物大豆的边行由于玉米遮阴,光截获明显减少,产量降低[4,5]。但目前条带间作中带宽对光截获行间差异影响的定量化研究还鲜有报道。
间作系统中冠层结构的空间异质性导致光截获的测定比植物单一种植时更困难。模型模拟是研究间作系统光截获的有效工具。最初,基于水平层和平行行向的垂直层将间作复合空间划分成不同单元计算间作系统的光截获[6,7],并未考虑间作系统内的种间相互作用。随后,通过引入叶面积指数( Leaf Area Index , LAI )、冠层几何参数和消光系数等冠层结构特征进一步精确计算了从属作物的光截获[8,9]。这些模型多假设冠层内的叶面积为水平均质,仅考虑光的空间分布,很难准确量化冠层结构的时空变异性[9, 10]。同时,传统模型很难考虑间作系统光截获的行间差异,这对于间作系统种植配置及优化等的分析非常重要。
植物功能-结构模型 ( Functional-Structural Plant Model , FSPM )基于试验数据,可真实地反映作物结构的时空动态,量化群体空间内植物的表型可塑性和相互竞争[11-13]。目前,基于植物生长模型研究的间作系统光分布主要注重于区分系统内不同作物的光截获[14-17]。条带间作在中国种植面积广泛[18],先后构建了小麦-玉米[19]、小麦-棉花[20]和玉米-大豆[5] 的条带间作模型,计算了间作系统的光分布,评价了由植物表型可塑性和群体结构对系统光分布的贡献。植物 FSPM 通过模拟不同情境下间作系统中几种作物的生长发育,可明确解决间作系统内时空差异下植物的表型可塑性反应对光截获和光能利用的影响,进一步弥补田间试验测量的局限。然而,目前鲜有对同种同收玉米-大豆间作系统作物光截获行间差异的研究。
本研究的主要目的为量化不同间作系统内植物光截获的行间差异。因此,本研究量化了种植模式、带宽和行位置对植物表型性状的影响,并用于构建玉米-大豆间作 FSPM;利用 FSPM的方法量化不同间作系统中光截获的行间差异,为解释条带间作光截获对产量优势的贡献以及不同光环境下的行配比优化等提供依据。
2 材料与方法
2.1 田间试验
2.1.1 试验概况
田间试验于2017—2018年在吉林省四平市梨树县的中国农业大学梨树实验站(43°16′N,124°26′E)进行。试验区0~30 cm的土层土壤为粉质粘壤土。试验共设置4个处理,分别为单作玉米( SM )、单作大豆( SS )、玉米-大豆2:2间作(2行玉米:2行大豆,2:2 MS )和玉米-大豆3:6间作(3行玉米:6行大豆,3:6 MS),每个处理3次重复。2:2 MS 间作中各行均为边行,3:6MS 间作中 M1、M3、S1和 S6为边行,M2为玉米内行, S2和 S5为大豆内 I行, S3和 S4为大豆内 II行(图1)。单作种植时玉米和大豆的行距均为50 cm ,株距分别为20 cm和10 cm 。间作种植时玉米行与大豆行相距50 cm ,玉米和大豆条带内的株行距与单作时相同,所有处理均为东南—西北行向种植。试验地每个小区的面积均约为24 m×10 m。
供试玉米(Zea mays)品种为郑单958,大豆(Glycine max)品种为吉育47。玉米和大豆同时播种,播种日期为2017年5月11日和2018年5月10日。两年各种植模式采用相同的施肥方案。播种前,分别施用基肥 N 80 kg/ha 、P2O5120 kg/ha 、 K2O 100 kg/ha;玉米 V8和 V16期(出现叶龄的叶片数分别为8和16片)单作玉米和间作玉米行间追施 N 80 kg/ha 。试验期间的气象数据由实验站内的气象站(雨根,中国)提供,图2为实验期间的日平均气温、降雨量和光合有效辐射。
2.1.2 测定内容
不同处理均设置田间原位观测和破坏性取样测量。田间原位观测从作物出苗后开始,从植株底部依次向上记录叶位信息,每3~7 d田间原位观测玉米和大豆的叶片出现、完全展开和衰亡时间。观测时,单作小区随机选择6株植株,间作小区随机选择一个完整条带,且每行随机选择6株植株。出叶间隔指连续叶尖出现所需的热时间,为热时间与叶片数线性拟合的斜率( oCd/leaf)。叶片完全展开后,用数字量角器(误差:±0.1o )测量叶倾角(叶片偏离茎秆生长时产生的叶片或叶柄与茎秆的最小角度)。
作物出苗25 d后,每15 d对各小区的作物器官,如株高、叶片和节间进行破坏性测量。单作小区和间作条带每行随机选取具有代表性的2株作物,测定每个叶位上每个器官的形态数据(叶片长度、宽度、节间长度、节间中部直径和叶柄长度)。单作小区和间作条带每行随机选取4株作物,用 LI-COR叶面积仪( Lincoln ,NB ,US )测量作物的单株叶面积,根据作物所占地面面积计算不同种植模式时的 LAI。
試验期间田间用 QSO-S 光合有效辐射(Photosynthetically Active Radiation , PAR ) 单点传感器(Decagon , US )每隔10 min 连续记录2:2 MS 间作冠顶和冠底的 PAR 变化,用于评估所建模型。监测样点分布在一个完整间作条带的作物行间,冠底共3个探头,水平间距50 cm。冠顶共3个探头,水平间距50 cm ,样点始终保持在冠顶上方50 cm处。同一行间的冠顶和冠底探头在一条垂直线上(图1(c))。
测得的动态辐射值先换算为日辐射值,冠顶日辐射值Ltop与冠底日辐射值Lbottom之差为冠层的日光截获量Lcanopy。因此,冠层光截获系数f为公式( 1)所示。
2.2模型原理
基于GroIMP (http ://www. sourceforge. net/ projects/groimp/)开源平台开发了玉米-大豆间作的 FSPM [11,21]。此模型主要包括植物器官生长发育和外界辐射环境两个子模块。由于本研究的科学问题需要精确计算与植物表型相关的光截获,因此模型构建时以田间实测器官尺寸为输入,计算不同处理的光截获量。
模型中用有效积温来模拟植株的发育进程,用 Beta生长函数描述所有器官的扩展[22-24],用反向 Monte-Carlo 光线追踪算法模拟冠层的光分布[25]。
模型模拟时单作群体为6×10株,间作群体包括一个玉米条带和一个大豆条带,即 2:2 MS为2×10株玉米和2×20株大豆,3:6 MS 为3×10株玉米和6×20株大豆。为使入射光引起的边行效应最小化,利用GroIMP平台中的复制器功能将模拟群体分别在x和y方向上复制10次。
2.2.1 有效积温与植物生长发育关系
作物的物候学发育决定于种植到收获期间的总有效积温(或热时间, Tsum ,oCd )。 Tsum 的计算基于日平均气温( Tave )和作物基础温度( Tb ),本研究中玉米和大豆生长的基础温度分别为8 oC和10 oC [26,27]。Tsum 也是模拟叶片发育(叶片生长和衰亡)和节间发育的指标,如公式(2) 和公式(3) 所示。
Tsum =∑max (0,( Tave - Tb )) ( 2)
Ni = a×Tsum,i (3)
其中,Ni 表示植物出苗后第i天时的叶片数,个;Tsum,i表示从种植到出苗后第i天时的有效积温,oCd;a表示线性回归线的斜率; max指当日平均气温低于 Tb 时,Tsum 为0。
2.2.2 器官尺寸与叶位关系
模型中玉米、大豆叶片和大豆叶柄最终长度与叶位的关系均用 Cauchy 分布函数描述[5,28,29],如公式(4) 所示。
其中,i指代叶片时为 B ,指代叶柄时为 P;Li,m为器官最终长度的最大值, cm; bi 为尺度参数;ri,m为器官最终长度达到最大值时的叶位。
线性关系模型用于描述叶长LB,r和叶宽LB,w之间的关系[5],如公式(5) 所示。
其中,LB,r为叶位 r 时的最终叶长, cm; a1为线性模型的斜率,cm/cm; b1为线性模型的截距,cm。
Logistic 方程用于描述玉米节间的最终长度[5],如公式(6) 所示。
其中,Im,r为玉米在叶位 r 时的节间最终长度,cm;Im为节间长度最大值,cm;r1为斜率转折点叶位,k1为斜率。
Cauchy 分布函数用于描述大豆不同节间最终长度随叶位的变化[5],如公式(7) 所示。
其中,Is,r表示第 r 叶位时的节间最终长度,cm;II,m表示节间最终长度的最大值,cm;rI,m为达到节间最终长度最大值时的叶位;bI为尺度参数。
节间直径与叶位的关系用递减的 Logistic 方程描述[5,30],如公式(8) 所示。
其中,Dr 表示第 r 叶位时的节间最终直径, mm; Dm 表示植物基部节间所能达到的最大直径, mm; k2表示斜率系数; r2为拐点出现的叶位。
玉米叶倾角和大豆沿主茎叶柄倾角的分布用负指数方程描述[5],如公式(9) 所示。
其中,βr 为玉米和大豆沿主茎叶位 r 时的叶倾角, ( o ); a2 + c 为理论0叶位时的叶倾角,(°); b2为水平范围; c 为当 r 逐渐变大时的渐近线。
2.2.3 太阳辐射模拟
为计算叶片水平的光截获,模型引入了光源及叶片光学性质参数。模型中入射光源由直射辐射和散射辐射组成,直射辐射由24个定向光源组成,代表一天中不同时刻的太阳位置[11, 19,31]。尽管模型的时间步长为1天,但模型综合考虑了一天内不同时刻的光变化[5]。散射辐射由天空半球72个方向的定向光源组成[11, 19]。本研究所构建的辐射模型已得到广泛验证和应用[5, 11, 19,31]。总入射辐射强度的日变化由纬度和一年中的天数计算,并用于计算不同种植模式下的植物光截获,大气透射率為0.339。总辐射中的67%为散射辐射,其余的33%为直射辐射[32]。尽管模型的直射辐射和散射辐射比例不变,但模型综合考虑了生育期内的实际天气变化,模型输出日辐射强度与实际日辐射强度间的均方根误差( Root MeanSquare Error ,RMSE )为4.3 MJ/(m2·d)。
玉米叶片的 PAR 反射率为9.2%,透射率为1.3%[19]。大豆叶片的 PAR 反射率为7.9%,透射率为8.2%[33]。由于试验中未对玉米叶鞘进行测量,因此模型中并未考虑此器官。模型节间设为不透光对象,其反射率为叶片反射率和透射率的和。
模型中分别在作物行间的冠顶和冠底设置0.1 m×0.1 m的光传感器测量辐射值,行间布设同田间试验,利用公式(1) 计算光截获系数的模型模拟值。
2.3模型参数化
利用 R 语言‘bblme包中的嵌套模型(mle2) 分析叶片最终尺寸、节间最终尺寸、叶柄角度和叶柄最终长度的分布[5]。
Li 等[4] 的研究结果表明,3:6 MS 间作中S2~S5的作物表型差异不显著,故模型参数化时未再区分大豆内行( S2~S5)植株的表型差异。本研究根据四种不同的行位置(单作、2:2 MS、3:6 MS 边行和3:6 MS 内行)对器官尺寸分布(公式(4) ~公式(9) )进行拟合,以判断不同处理间植物表型性状是否具有显著性差异。具体来说,数据拟合时的数据集合分类为(表1):①利用四个不同的数据集合,如每个行位置单独一个集合;②利用三个不同的数据集合,如单作一个,2:2 MS 一个,3:6 MS 边行和3:6 MS 内行为一个集合;或者单作一个,2:2 MS 和3:6 MS边行一个,3:6 MS内行为一个集合;或者单作和3:6 MS 内行一个,2:2 MS 一个,3:6 MS 边行为一个集合;③利用两个不同的集合,如单作一个,2:2 MS 、3:6 MS 边行和3:6 MS 内行为一个集合;或者单作和3:6 MS 内行一个,2:2 MS 和3:6 MS边行为一个集合;④利用一个集合,如四个行位置合成一个集合。
为明确不同行位置对植物表型性状的影响,公式参数拟合时有不同的假设。假如拟合公式有3个待拟合参数,对于不同行位置的公式参数共有4种假设情况:①3个参数均相同;②只有一个参数相同;③其中的两个参数相同和④3个参数均不相同。
为避免拟合参数冗余, Akaike 信息准则( Akaike Information Criterion , AIC ) 中以最小的 AIC 值为依据选择最好的数据拟合结果:ΔAIC <2时表示模型间无差异,选择最简单的模型;ΔAIC 在4~7之间表示模型间差异显著;ΔAIC >10时表示模型完全不同[34]。根据所选模型判断处理间植物表型相同或不同。
2.4模型评估
模型构建研究中一般将前一年的数据参数化,为验证模型参数或评价模型构建的准确性、精确性,通常只改变外界环境条件,验证之后年份或不同处理的群体表现[30]。因此,本研究将2017年的田间植物表型数据用于模型参数化,2018年的植株叶面积、株高和冠层光截获系数用于模型验证。
采用 RMSE 和一致性系数 d 检验株高、LAI 和光截获系数模型模拟值与实测值间的吻合程度:
其中,Pi 为模拟值;Si 为实测值;Pi'= Pi -S,Si'= Si -S ;n为模拟值或实测值的数量,个。d值的变化范围为0~1,其值越大表示模拟值与实测值越相近,模拟结果越精确。
3 结果与分析
3.1器官表型可塑性
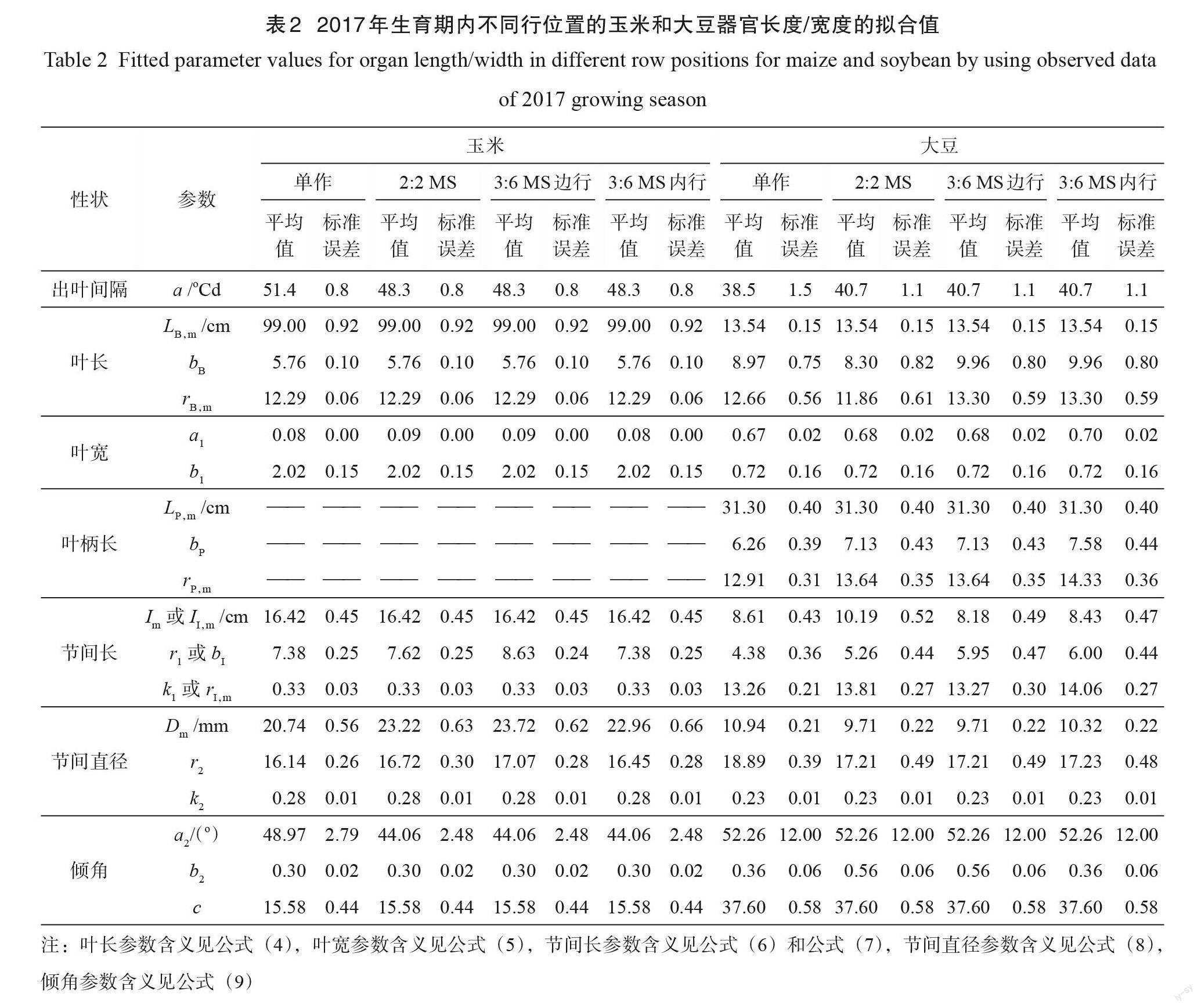
3.1.1 出叶间隔
间作玉米的出叶间隔,即连续叶尖出现的热时间为(48.3±0.8) oCd,比單作玉米短(51.4±0.8) oCd (表2)。间作种植增加了大豆的出叶间隔,且带宽对间作大豆的出叶间隔无显著影响(表2)。因此,模型中单作大豆的出叶间隔设为38.5 oCd,间作大豆的出叶间隔均设为40.7 oCd。
3.1.2 叶片
叶长随叶位呈先增后减的趋势。玉米叶片在单作和间作之间略有不同:虽然间作种植对玉米叶长未有影响,却增加了叶宽/叶长比值,且3:6 MS种植时边行玉米的叶片比内行宽(表2)。
与单作大豆相比,2:2 MS间作大豆的高叶位叶长显著减小,但叶宽稍大于单作大豆。3:6 MS 间作大豆高叶位的叶片大于单作,且内行大豆比边行大豆和2:2 MS 大豆叶片宽(表2)。间作种植降低了大豆3~13叶位的叶柄长,增加了第14叶位以上的叶柄长(表2)。3:6 MS 间作种植时边行大豆中低叶位的叶柄比内行长,高于第14叶位时边行大豆叶柄反而比内行短。
3.1.3 节间
处理间玉米节间长的最大值无显著差异,节间直径却差异显著(表2)。间作玉米的节间直径显著大于单作玉米,且3:6 MS 边行玉米的节间直径大于内行玉米。单作玉米节间直径的最大值仅为间作玉米的89%。
间作大豆节间长于单作大豆,且窄条带(2:2 MS )间作大豆的节间长于宽条带(3:6 MS )间作大豆(表2)。3:6 MS 间作中边行大豆高叶位(>14叶位)节间长小于内行大豆。间作大豆的节间直径小于单作大豆(表2)。行位置显著影响大豆节间直径,边行大豆的节间更细。单作大豆基部节间直径是间作边行大豆的1.13倍,是间作内行大豆的1.06倍。
3.1.4 倾角
玉米叶倾角随叶位升高而减小,且当叶位低于7时,单作玉米的叶倾角大于间作玉米。带宽和行位置对玉米叶倾角无显著影响。间作大豆中低叶位(<13叶位)的叶柄倾角显著小于单作大豆,且行位置对叶柄倾角影响显著:边行大豆的叶柄倾角显著小于内行大豆(表2)。
3.2玉米-大豆 FSPM 与冠层光截获校验
FSPM 真实地描述了玉米和大豆的结构发育,包括生长季内株高和叶面积的变化(以图3为例)。 LAI模拟值随叶片的产生和扩展而增加,随叶片的衰老和凋落而减小。株高随节间的产生和扩展而增加。出苗后65 d时,大豆单作地面的光照强度最低,2:2 MS 处理的地面光照强度最高,因此单作地面的颜色较间作深,冠层截获的光照与地面正好相反。玉米单作时冠层截获的光比例最低,因此穗位叶以下叶片颜色较深,2:2 MS 间作冠层截获的光最高(图3)。
尽管模型低估了单作和间作玉米的最大叶面积指数,但总体而言模型模拟值和实测值之间具有较好的一致性(表3)。处理间玉米和大豆 LAI 的 RMSE变化范围为0.24~0.70 m2/m2,一致性系数 d 的变化范围为0.931~0.992。处理间玉米和大豆株高的 RMSE 变化范围为0.06~0.17 m ,一致性系数 d 为0.983~0.998。2:2 MS 间作不同行位置的光截获系数也表明模型能够较好地模拟间作冠层不同位置的植物光截获, RMSE 在0.06~0.10之间(图4)。
3.3生育期内光截获行间差异定量化
基于构建的玉米-大豆间作 FSPM 计算间作条带中作物净单位土地面积光截获的行间差异,以明确带宽和行位置对条带光截获的影响。结果表明,出苗30 d后间作玉米的日光截获量高于单作,且3:6 MS 间作中边行( M1和 M3)的日光截获量高于内行( M2)(图5(a)和图5(b))。出苗20 d 后2:2 MS 间作大豆的日光截获量低于单作;出苗30~100 d时3:6 MS间作大豆的日光截获量低于单作,且边行( S1和 S6)的日光截获量低于内行 ( S2~S5) (图 5 (c) 和图5 (d))。
单作、2:2 MS 和3:6 MS 间作种植时,玉米生育期内净单位土地面积的累积光截获量分别为(819.9±0.6) MJ/m2、 (1111.9±3.7) MJ/m2 和(1027.1±5.0) MJ/m2;大豆生育期内净单位土地面积的累积光截获量分别为(709.3±0.6) MJ/m2、(453.6±3.3) MJ/m2和( 627.3±2.0) MJ/m2。生育期内玉米的光截获增加了25.3%~35.6%,大豆的光截获减少了11.6%~36.0%。
与单作相比,2:2 MS间作中的边行大豆( S1和 S2)光截获减少了36.0%(图6(a));3:6 MS间作大豆条带光截获具有一定的对称性,边行( S1和 S6)、内Ⅰ行( S2和 S5)和内Ⅱ行( S3和S4)的光截获量分别比单作减少了28.8%、4.1%和1.8%(图6(b))。2:2 MS 间作时边行大豆( S1和 S2)的光截获比3:6 MS 间作边行( S1和S6)的光截获量低10.2%;3:6 MS间作边行大豆( S1和 S6)的光截获量最少,比内Ⅱ行低27.5%。间作种植时玉米生育期内的光截获为单作的1.3倍,间作内行玉米的光截获比边行减少了5.9%(图6)。
4 讨论
条带间作中的高、矮秆作物搭配种植最终使植株表型出现明显的可塑性[3,35]。本研究结果表明由于高秆玉米的遮阴作用,间作大豆表现出明显的表型可塑性,主要表现为大豆节间伸长、节间变细(表2)、叶面积减小,进而间作大豆株高高于单作大豆。本研究的种植行向为东南—西北,宽条带间作(3:6 MS )大豆的避阴反应较弱,间作叶片大小较单作差异较小,而当大豆处于严重遮阴状态(窄条带间作,2:3 MS )且种植行向为南北时,大豆的避阴反应较强,间作叶片较单作差异较大[4,5]。由于种内竞争降低,玉米也表现出一定的表型可塑性,如叶片变宽和节间增粗(表2)。
本研究所建立的玉米-大豆間作 FSPM 能够准确模拟单个器官的生长信息,进而能够考虑间作种植时植株表型的可塑性,如株高增加、LAI 减少及冠层光截获的变化等(图3~图6、表3)。虽然本模型与 Li等[5]建立的玉米-大豆间作模型相似,但 Li 等[5] 的模型主要用于量化表型可塑性和冠层结构对光截获的贡献,同时量化了单个大豆表型对群体光截获的影响,探明了间作大豆光截获降低的主要限制表型。本研究意在量化由种植模式、带宽和行位置引起的间作植物表型和光截获的行间差异(表2、图5和图6),可为解释间作产量的行间差异提供科学依据。
间作种植改变了单一作物种植时群体的冠层结构,最终影响间作系统的光截获[5,36,37]。本研究基于构建的玉米-大豆间作 FSPM 计算了不同行配比下玉米和大豆冠层的光截获,结果表明由于玉米处于优势地位,间作玉米的光截获高于单作玉米。由于间作系统中高秆玉米对矮秆大豆的遮阴作用明显,间作大豆的光截获明显低于单作大豆。玉米大豆间作种植时,大豆冠顶入射辐射因间作系统中行配比不同而不同。Liu等[10] 的研究表明玉米-大豆1:1 MS间作时,大豆受玉米遮阴作用更明显,冠顶平均入射辐射比例比2:2 MS处理低23.8%,间作系统中大豆条带越宽冠顶入射辐射越多。因此,随着大豆条带的增加,大豆条带整体光截获得到改善 (图5和图6)。3:6MS间作时大豆冠层单位土地面积上的光截获是2:2 MS间作时的1.4倍,地面的光截获比例则比2:2 MS间作时少(图3和图5)。在不降低大豆节间韧性的同时增加其长度,有助于大豆规避玉米遮阴的影响,促进光截获量的增加[5]。
条带间作中高秆作物对矮秆作物的遮阴作用主要受种植时间[19,38]、带宽[4,39]、种植行向[38]、叶面积密度以及株高的综合影响[8]。间作 FSPM能够综合考虑这些指标及其变化,对间作组成物种的光截获贡献等能够很好地分解和阐述。本研究中玉米和大豆虽为东南—西北行种植,但两作物同种同收,大豆受玉米遮阴作用仍较显著。条带间作大豆不同行之间的光截获也明显不同,距离玉米越近的大豆光截获越低,光截获最多相差27.6%(图5和图6)。受试验条件和仪器的限制,本研究并未对不同处理、不同叶位叶片的光学参数进行测定。品种不同这些参数差异很大,且作物对间作不同行配置的环境响应不同,未来将进一步补充测定数据,使模型对作物光截获的模拟更接近真实情况。
大田环境是一个非常复杂的农业生态系统,作物的生长发育受温度、光照、水分、养分、CO2浓度以及风等多个环境因子及生物因素的综合影响,且这些因素具有年季间差异。田间环境因素和耕种管理措施的改变均会引起作物表型和干物质累积的变化。尽管大田环境因素众多,本研究构建的作物功能-结构模型是在考虑温度和光照两大可变因素、耕种和管理两大不变因素条件下模拟的作物器官形态发育及冠层内的光分布,最终计算冠层的光截获量。此模型既考虑了作物与环境因素的相互作用,又考虑了作物与作物间的相互作用。尽管此模型仅考虑了部分环境因素和生物因素的作用,但模型可用于探索这些因素综合作用下作物表型的响应,探索这些因素最终引起的光截获差异,解释间作产量行间差异。作物功能-结构模型的发展是持续的,在国内外研究人员的共同努力下该模型已引入光合作用、源库分配关系、氮素利用等因素。同时还在构建地下根系模型,进一步完善了作物与环境的互作以及地上部与地下部的反馈。未来该模型还可以引入更多的环境因素,用于探索更深层次的作物生长机理。
5 结论
本研究基于2017—2018年田间试验,构建了玉米-大豆间作 FSPM ,模拟了由种植方式、带宽和行位置引起的植株表型可塑性变化,量化了不同间作处理作物生长发育和光截获的行间差异。间作种植显著增加了玉米节间直径。受玉米遮阴影响,大豆节间变长、变细,且随大豆条带变窄差异越明显。2:2 MS间作玉米光截获比单作玉米高35.6%,3:6 MS边行玉米和内行玉米分别比单作玉米高27.8%和20.3%。2:2 MS与3:6 MS 边行大豆的光截获比单作分别少36.0%和28.8%;3:6 MS大豆内 I行和内 II行比单作大豆的光截获分别少4.1%和1.8%。
间作系统中大豆叶片变大、节间变长有助于改善间作系统的生产力。间作种植时,带宽影响玉米对大豆的遮阴作用。因此,利用本模型可研究矮化玉米品种、增加玉米条带与大豆条带的间距、去除玉米雄穗或部分叶片等管理措施对间作大豆生长、光截获和产量的影响。本研究还可为间作种植的优化配置、大豆耐阴品种的筛选等提供依据。
参考文献:
[1] WANG Q, BAI W, SUN Z, et al. Does reduced intra‐specific competition of the dominant species in inter‐crops allow for a higher population density?[J]. Foodand Energy Security, 2021, 10(2):285-298.
[2] RAZA M A, FENG L Y, IQBAL N, et al. Growth anddevelopment of soybean under changing light environ‐ments in relay intercropping system[J]. PeerJ, 2019, 7:ID e7262.
[3] LIU X, RAHMAN T, SONG C, et al. Relationshipsamong light distribution, radiation use efficiency andland equivalent ratio in maize-soybean strip intercrop‐ping[J]. Field Crops Research, 2018, 224:91-101.
[4] LI S, EVERS J B, VAN DER WERF W, et al. Plant ar‐chitectural responses in simultaneous maize/soybeanstrip intercropping do not lead to a yield advantage[J].Annals of Applied Biology, 2020, 177(2):195-210.
[5] LI S, VAN DER WERF W, ZHU J, et al. Estimatingthe contribution of plant traits to light partitioning in si‐multaneous maize/soybean intercropping[J]. Journal ofExperimental Botany, 2021, 72(10):3630-3646.
[6] SINOQUET H, BONHOMME R. Modeling radiativetransfer in mixed and row intercropping systems[J].Agricultural and Forest Meteorology, 1992, 62(3-4):219-240.
[7] TSUBO M, WALKER S. A model of radiation inter‐ception and use by a maize-bean intercrop canopy[J].Agricultural and Forest Meteorology, 2002, 110(3):203-215.
[8] MUNZ S, GRAEFF-H?NNINGER S, LIZASO J I, etal. Modeling light availability for a subordinate cropwithin a strip-intercropping system[J]. Field Crops Re‐search, 2014, 155:77-89.
[9] WANG Z, ZHAO X, WU P, et al. Radiation intercep‐tion and utilization by wheat/maize strip intercroppingsystems[J]. Agricultural and Forest Meteorology, 2015,204:58-66.
[10] LIU X, RAHMAN T, YANG F, et al. PAR interceptionand utilization in different maize and soybean inter‐cropping patterns[J]. PLoS ONE, 2017, 12(1): IDe0169218.
[11] EVERS J B, VOS J, YIN X, et al. Simulation of wheatgrowth and development based on organ-level photo‐synthesis and assimilate allocation[J]. Journal of Exper‐imental Botany, 2010, 61(8):2203-2216.
[12] VOS J, EVERS J B, BUCK-SORLIN G H, et al. Func‐tional-structural plant modelling: A new versatile toolin crop science[J]. Journal of Experimental Botany,2010, 61(8):2101-2115.
[13] EVERS J B, VAN DER WERF W, STOMPH T J, et al.Understanding and optimizing species mixtures usingfunctional-structural plant modeling[J]. Journal of Ex‐perimental Botany, 2019, 70(9):2381-2388.
[14] SONOHAT G, SINOQUET H, VARLET-GRANCHERC, et al. Leaf dispersion and light partitioning in three- dimensionally digitized tall fescue-white clover mix‐tures[J]. Plant Cell and Environment, 2002, 25(4):529-538.
[15] CICI S Z, ADKINS S, HANAN J. A canopy architectural model to study the competitive ability of chickpea with sow thistle[J]. Annals of Botany, 2008, 101(9):1311-1318.
[16] BARILLOT R, FOURNIER C, HUYNH P, et al. Howdo variations of architectural parameters affect light partitioning within wheat-pea mixtures? A simulation study based on a virtual plant approach[C]// Interna‐tional Conference on Functional Structure Plant Mod‐els. Saariselk? , Finland: Finnish Society of Forest Sci‐ence, 2013:354-354.
[17] BARILLOT R, ESCOBAR-GUTI?RREZ A J,FOURNIER C, et al. Assessing the effects of architec‐tural variations on light partitioning within virtual wheat-pea mixtures[J]. Annals of Botany, 2014, 114(4):725-737.
[18] LI L, ZHANG W, ZHANG L. How above- and below-ground interspecific interactions between intercropped species contribute to overyielding and efficient re‐ source utilization: A review of research in China: Sci‐ence, practice, and sustainable management[M]// SHIMING L, GLIESSMAN S R. Agroecology in Chi‐na. Boca Raton: CRC Press, 2016:39-59.
[19] ZHU J, VAN DER WERF W, ANTEN N P R, et al.The contribution of phenotypic plasticity to comple‐mentary light capture in plant mixtures[J]. New Phytol‐ogist, 2015, 207(4):1213-1222.
[20] MAO L, ZHANG L, EVERS J B, et al. Identificationof plant configurations maximizing radiation capture in relay strip cotton using a functional-structural plant model[J]. Field Crops Research, 2016, 187:1-11.
[21] HEMMERLING R, KNIEMEYER O, LANWERT D,et al. The rule-based language XL and the modelling environment GroIMP illustrated with simulated tree competition[J]. Functional Plant Biology, 2008, 35(10):739-750.
[22] GUO Y, MA Y, ZHAN Z, et al. Parameter optimizationand field validation of the functional-structural model GREENLAB for maize[J]. Annals of Botany, 2006, 97(2):217-230.
[23] MA Y, WEN M, GUO Y, et al. Parameter optimizationand field validation of the functional-structural model GREENLAB for maize at different population densi‐ ties[J]. Annals of Botany, 2007, 101(8):1185-1194.
[24] YIN X, GOUDRIAAN J, LANTINGA E A, et al. Aflexible sigmoid function of determinate growth[J]. An‐nals of Botany, 2003, 91(6):753-753.
[25] VEACH E. Robust Monte Carlo methods for lighttransport simulation[D]. Stanford: Stanford University,1998.
[26] MAJOR D J, JOHNSON D R, TANNER J W, et al. Ef‐fects of daylength and temperature on soybean devel‐opment[J]. Crop Science, 1975, 15:174-179.
[27] RITCHIE J T, NESMITH D S. Temperature and cropdevelopment[M]// HANKS J, RITCHIE J T. ModelingPlant and Soil Systems. New York: The American Soci‐ety of Agronomy and Academic Press, 1991, 5-29.
[28] BUCK-SORLIN G H. L-system model of the vegeta‐tive growth of winter barley[M]// POLANI D, KIM J,MARTINEZ T. eds. Fifth German Workshop on Artifi‐cial Life. Lübeck: AkademischeVerlagsgesellschaftAka GmbH, 2002:53-64.
[29] EVERS J B, VOS J, FOURNIER C, et al. Towards ageneric architectural model of tillering in Gramineae,as exemplified by spring wheat (Triticum aestivum)[J].New Phytologist, 2005, 166(3):801-812.
[30] GU S, EVERS J B, ZHANG L, et al. Modelling thestructural response of cotton plants to mepiquatchlo‐ride and population density[J]. Annals of Botany, 2014,114(4):877-887.
[31] BUCK-SORLIN G, DE VISSER P H B, HENKE M, etal. Towards a functional-structural plant model of cut-rose: Simulation of light environment, light absorption,photosynthesis and interference with the plant struc‐ture[J]. Annals of Botany, 2011, 108(6):1121-1134.
[32] ZHOU J, WU Y, YAN G. Estimation of daily diffusesolar radiation in China[J]. Renewable Energy, 2004,29(9):1537-1548.
[33] KASPERBAUER M J. Far-red light reflection fromgreen leaves and effects on phytochrome-mediated as‐similate partitioning under field conditions[J]. PlantPhysiology, 1987, 85(2):350-354.
[34] BOLKER B M. Ecological models and data in R[M].Princeton: Princeton University Press, 2008.
[35] LIU X, RAHMAN T, SONG C, et al. Changes in lightenvironment, morphology, growth and yield of soy‐bean in maize-soybean intercropping systems[J]. FieldCrops Research, 2017, 200:38-46.
[36] WANG Q, SUN Z, BAI W, et al. Light interceptionand use efficiency differ with maize plant density inmaize-peanut intercropping[J]. Frontiers of Agricultur‐al Science and Engineering, 2021, 8(3):432-446.
[37] GAO Y, DUAN A, QIU X, et al. Distribution and useefficiency of photosynthetically active radiation instrip intercropping of maize and soybean[J]. ChineseJournal of Applied Ecology, 2010, 102(4):952-958.
[38] WANG Z, ZHAO X, WU P, et al. Border row effectson light interception in wheat/maize strip intercroppingsystems[J]. Field Crops Research, 2017, 214:1-13.
[39] WANG Q, SUN D, HAO H, et al. Photosyntheticallyactive radiation determining yields for an intercrop ofmaize with cabbage[J]. European Journal Agronomy,2015, 69:32-40.
Estimating the Differences of Light Capture Between Rows Based on Functional-Structural Plant Model inSimultaneous Maize-Soybean Strip Intercropping
LI Shuangwei1,2,3 , ZHU Junqi4 , Jochem B. EVERS3 , Wopke VAN DER WERF3 , GUO Yan1 , LI Baoguo1 , MA Yuntao1*
(1. College of Land Science and Technology, China Agricultural University, Beijing 100193, China;2. Institute of Agricultural Equipment, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China;3. Centrefor CropSystems Analysis, Wageningen University, Wageningen 430-6700 AK, The Netherlands;4. Marlborough ResearchCentre, The New Zealand Institutefor Plant and Food Research Limited, Blenheim 7240, New Zealand)
Abstract: Intercropping creates a heterogeneous canopy and triggers plastic responses in plant growth and structural develop‐ment. In order to quantify the effect of planting pattern, strip width and row position on the structural development and light cap‐ture of maize and soybean in simultaneous intercropping, both experimental and modelling approaches were used. Field experi‐ments were conducted in 2017-2018 with two sole crops (maize and soybean) and two intercrops: Two rows of maize alternat‐ing with two rows of soybeans (2:2 MS) and three rows of maize alternating with six rows of soybean (3:6 MS). The morpho‐ logical traits of maize and soybean e.g., leaf length and width, internode length and diameter, leaf and petiole declination angle in different rows and different planting patterns, and photosynthetically active radiation (PAR) above and below the canopy of 2:2 MS were measured throughout the growing season. A functional-structural plant model of maize-soybean intercropping was developed in the GroIMP platform. The model was parameterized based on the morphological data set of 2017, and was validat‐ ed with the leaf area index (LAI), plant height and PAR data set of 2018. The model simulated the morphological development of individual organs based on growing degree days (thermal time) and calculated the light capture at leaf level. The model well reproduced the observed dynamics of leaf area index and plant height (RMSE :0.24-0.70 m2/m2 for LAI and 0.06-0.17 m for plant height), and the fraction of light capture in the 2:2 MS intercropping (RMSE :0.06-0.10). Maize internode diameter in in‐tercrops increased, but the internode length did not change. Soybean internodes in intercrops became longer and thinner com ‐ pared to sole soybean probably caused by the shading imposed by maize, and the 2:2 MS had longer internodes than the 3:6 MS, indicating the effects of strip width. Simulated light capture of maize in 2:2 MS intercropping was 35.6% higher than sole maize. For maize in 3:6 MS intercropping, the light capture of the border rows and inner row were 27.8% and 20.3% higher than sole maize, respectively. Compared to sole soybean, the simulated light capture of soybean in border rows was 36.0% low ‐ er in 2:2 MS intercropping, and was 28.8% lower in 3:6 MS intercropping. For 3:6 MS intercropping, light capture of soybean in inner rows I and inner rows II were 4.1% and 1.8% lower than sole soybean, respectively. In the future, the model could be further developed and used to explore and optimize the planting patterns of maize soybean intercropping under different envi‐ronmental conditions using light capture as an indicator.
Key words: maize-soybean intercropping; functional-structural plant model; light capture; three-dimensional structure; pheno‐ type plasticity; row difference
猜你喜欢
农业与技术(2023年11期)2023-06-19 02:28:18
经济林研究(2022年4期)2023-01-12 09:14:46
干旱地区农业研究(2022年4期)2022-07-28 05:37:08
四川蚕业(2022年1期)2022-06-06 02:03:42
广西林业科学(2022年2期)2022-05-09 07:49:54
河北果树(2020年4期)2020-11-26 06:05:18
山东农业科学(2019年11期)2019-12-24 01:11:27
江苏农业科学(2019年11期)2019-07-22 01:32:45
干旱地区农业研究(2017年5期)2017-12-18 06:18:18
新疆农垦科技(2016年2期)2016-08-21 13:50:18
