
基于EST-SSR标记的青榨槭天然种群遗传多样性分析
2022-05-24 10:37张梦璐白云海张睿鹂窦德泉
植物资源与环境学报 2022年2期
穆 莹, 张梦璐, 白云海, 张睿鹂, 郑 健, 窦德泉
(北京农学院园林学院 城乡生态环境北京实验室, 北京 102206)
青榨槭(AcerdavidiiFranch.)隶属于槭树科(Aceraceae)槭属(AcerLinn.),为落叶乔木,该种生长迅速,树冠整齐,树姿优美,树皮呈蛙绿色,叶片在秋季可变为黄色、红色或橙色,具有极佳的观赏价值,是城市园林绿化的优良树种[1]。此外,青榨槭还具有叶片富含蛋白酶、材质优良等特点,具有一定的经济价值和医用价值[2,3]。然而,由于青榨槭存在天然生境遭到人为破坏和自身更新困难等问题,其遗传多样性面临巨大危机。目前,关于青榨槭天然种群遗传多样性的研究报道甚少,仅He等[4]利用青榨槭叶绿体基因组开发出20对SSR引物,并对陕西7个青榨槭种群的遗传多样性进行了分析。因此,亟需对青榨槭遗传多样性和遗传结构进行系统研究,这对其遗传多样性保护策略的制定具有重要意义。
微卫星标记(microsatellite marker)又称简单重复序列标记(simple sequence repeat, SSR),广泛分布于真核生物基因组的编码区和非编码区[5],具有共显性、多态性高、对DNA要求不严格、重复性好及符合孟德尔遗传规律等优点,被广泛应用于遗传多样性分析、比较作图、濒危物种保护、品种鉴定及种质资源保存等方面研究[6-9]。EST-SSR标记是基于表达序列标签(expressed sequence tag,EST)开发的一种新型分子标记,在同科不同属或同属不同种间具有良好的通用性,被广泛应用于植物基因组学研究[10-13]。
鉴于此,本研究利用28对多态性稳定、条带清晰的EST-SSR引物对北京、河南、陕西、广东和浙江5个地区的青榨槭8个天然种群231株样株的遗传多样性和遗传结构进行了系统分析,以期为青榨槭天然种群的遗传多样性保护及该遗传资源的开发利用奠定理论基础。
1 材料和方法
1.1 材料
根据青榨槭在中国的实际分布情况,选取北京、河南、陕西、广东和浙江的青榨槭8个天然种群作为研究对象,共采集231株样株的完整叶片,各种群的地理分布和样株数见表1。其中,北京天池峡谷和陕西南五台山地质公园2个种群的叶片样本于2019年10月采集;北京白羊沟自然风景区、北京三岔万涧、北京黄柏寺村、河南嵩山地质公园和广东南岭国家森林公园5个种群的叶片样本于2020年10月采集;浙江西天目山森林公园种群的叶片样本于2020年11月采集。样本采集时保证样株间距在30 m以上,每个种群采集30株以上,样株数量少于30株的种群则采集种群中的全部样株。采集样株顶部枝条上的健康、无病虫害的完整嫩叶,置于装有硅胶的塑封袋中保存并带回实验室。
1.2 方法
1.2.1 基因组DNA的提取和质量检测 使用CTAB植物基因组DNA快速提取试剂盒(北京爱博森生物科技有限公司)提取每株样株的基因组DNA。使用质量体积分数1%琼脂糖凝胶电泳检测基因组DNA样品的完整性,使用ND-1000超微量分光光度计(美国NanoDrop公司)检测基因组DNA样品的浓度和纯度,并将DNA质量浓度稀释至50 ng·μL-1后,置于-20 ℃冰箱中保存、备用。
1.2.2 PCR扩增条件及扩增结果检测 以每株样株的基因组DNA为模板,使用28对多态性稳定且扩增条带清晰的EST-SSR引物(表2)进行PCR扩增。

表1 青榨槭8个天然种群的地理分布和样株数

表2 用于青榨槭8个天然种群样株基因组DNA扩增反应的引物序列
所有扩增反应均使用S1000 PCR仪(美国ABI公司)完成,扩增体系总体积10.00 μL,包括基因组DNA样品1.00 μL、2×Taq Master Mix for PAGE(北京康为世纪生物科技有限公司)5.00 μL、10 μmol·L-1正向和反向引物各0.25 μL、ddH2O 3.50 μL。扩增程序为94 ℃预变性5 min;94 ℃变性30 s、相应退火温度下退火30 s、72 ℃延伸45 s,共32个循环;最后,72 ℃延伸10 min。
使用质量体积分数1%琼脂糖凝胶电泳检测扩增结果,检测完毕后将扩增产物转交北京睿博兴科生物技术有限公司进行毛细管电泳检测分析,检测仪器为ABI3730基因测序仪(美国ABI公司)。
1.3 数据处理
利用Genemarker Version 2.2.0软件读取SSR荧光标记片段大小,统计每个样本的基因型,据此构建231个样本的基因型数据库。利用POPGENE 32软件计算观测等位基因数、有效等位基因数、观测杂合度、期望杂合度、Shannon’s多样性指数、Nei’s基因多样性指数、基因流、遗传分化系数、遗传距离和遗传相似系数。利用GenAlEx 6.5软件进行Mantel检验,分析种群间遗传距离与地理距离的相关性,并进行分子方差分析(AMOVA),计算种群内及种群间的遗传变异贡献率。利用Cervus Version v3.0.7软件计算多态性信息含量。基于遗传距离、利用NTSYS-pc2.1软件对供试种群进行UPGMA聚类分析和绘图。利用STRUCTURE 2.3.4软件对供试种群进行分组和遗传结构分析,其中,遗传结构分析采用混合模型与等位基因相关模型,具体参数如下:分组K设为1~8,每个K值重复运算20次,将马尔科夫链蒙特卡罗(MCMC)和burn-in均设为100 000。运行结束后,将结果压缩(zip文件)并上传到Structure Harvester网站(http:∥taylor0.biology.ucla.edu/structureHarvester/)上,计算ΔK,得到最佳分组的K值。
2 结果和分析
2.1 青榨槭天然种群的遗传多样性分析
基于28对EST-SSR引物对青榨槭8个天然种群基因组DNA的扩增结果,供试青榨槭天然种群的遗传多样性分析结果见表3。
由表3可见:广东南岭国家森林公园(P7)种群和浙江西天目山森林公园(P8)种群的观测等位基因数为4,其余6个种群的观测等位基因数均为3,供试种群观测等位基因数的均值为3.2。P7种群的有效等位基因数最多,为1.8;北京黄柏寺村(P4)种群的有效等位基因数次之,为1.7;北京天池峡谷(P3)种群和P8种群的有效等位基因数最少,仅为1.5;其余4个种群的有效等位基因数均为1.6;供试种群有效等位基因数的均值也为1.6。
由表3还可见:P4种群的观测杂合度最低(0.197 6),P7种群的观测杂合度最高(0.298 9),供试种群观测杂合度的均值为0.237 9;P8种群的期望杂合度最低(0.244 4),P7种群的期望杂合度最高(0.307 7),供试种群期望杂合度的均值为0.276 9;河南嵩山地质公园(P5)种群的Shannon’s多样性指数最低(0.441 5),P7种群的Shannon’s信息指数最高(0.599 9),供试种群Shannon’s多样性指数的均值为0.512 7;P5种群的Nei’s基因多样性指数最低(0.240 1),P7种群的Nei’s基因多样性指数最高(0.302 8),供试种群Nei’s基因多样性指数的均值为0.271 9;P5种群的多态性信息含量最低(0.214 8),P7种群的多态性信息含量最高(0.276 0),供试种群多态性信息含量的均值为0.243 6。
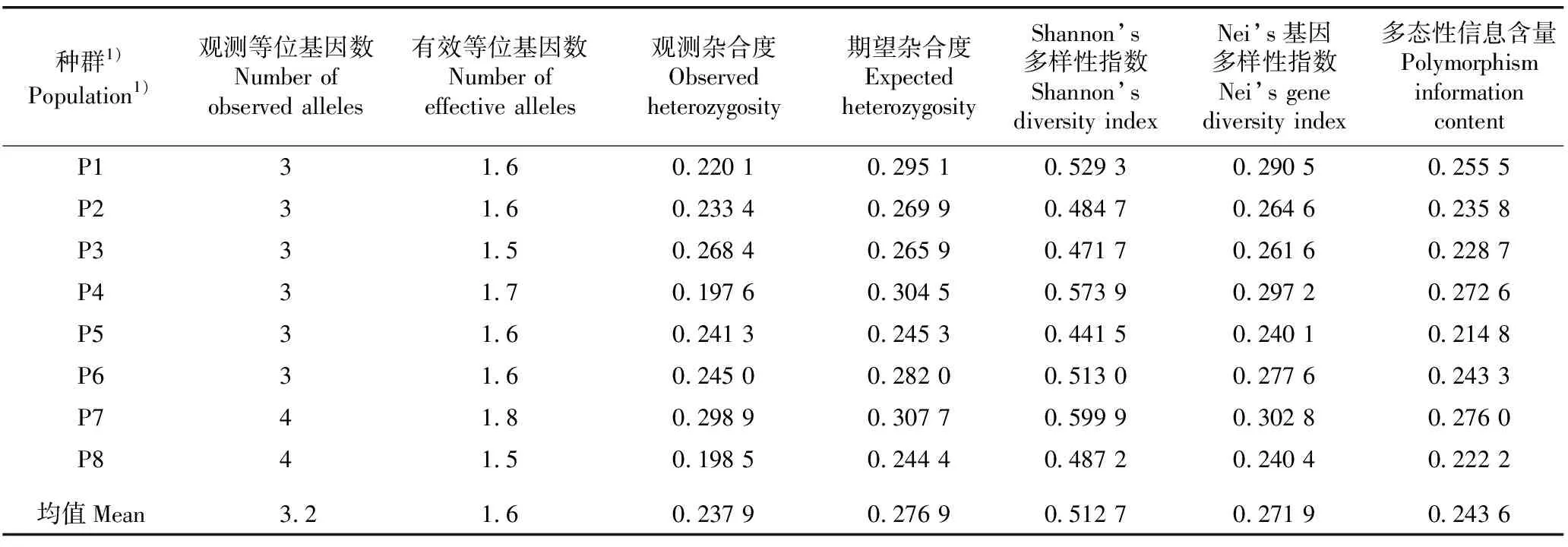
表3 基于EST-SSR引物扩增结果青榨槭8个天然种群的遗传多样性分析
整体来看,在供试种群中,P7种群的遗传多样性最高,P5种群的遗传多样性最低,且各种群的不同遗传多样性指标存在一定差异。例如:基于Shannon’s多样性指数进行排序,供试种群的遗传多样性从大到小依次为P7种群、P4种群、北京白羊沟自然风景区(P1)种群、陕西南五台地质公园(P6)种群、P8种群、北京三岔万涧(P2)种群、P3种群、P5种群;基于Nei’s基因多样性指数进行排序,供试种群的遗传多样性从大到小依次为P7种群、P4种群、P1种群、P6种群、P2种群、P3种群、P8种群、P5种群。
2.2 青榨槭天然种群的遗传分化和遗传结构分析
分子方差分析(AMOVA)结果表明:青榨槭8个天然种群间的遗传分化系数为0.334 5,种群间基因流为0.508 8,种群内遗传变异贡献率为66.55%,而种群间遗传变异贡献率为33.45%。说明青榨槭天然种群的遗传分化主要分布于种群内。
青榨槭8个天然种群间的遗传相似系数和遗传距离见表4。结果显示:供试种群间的遗传相似系数为0.584 3~0.960 3,其中,北京白羊沟自然风景区(P1)种群与北京三岔万涧(P2)种群间的遗传相似系数最大,北京黄柏寺村(P4)种群与浙江西天目山森林公园(P8)种群间的遗传相似系数最小。值得注意的是,广东南岭国家森林公园(P7)种群和P8种群与其余6个种群间的遗传相似系数相对较低,但这2个种群间及其余6个种群间的遗传相似系数却较高。供试种群间的遗传距离为0.040 5~0.537 3,其中,P1种群与P2种群间的遗传距离最小,P4种群和P8种群间的遗传距离最大。同样,P7种群和P8种群与其余6个种群间的遗传距离相对较大,而这2个种群间及其余6个种群间的遗传距离却较小。据此推断青榨槭天然种群间的遗传距离与地理距离呈正关联。Mantel检验结果表明:各种群间的遗传距离与地理距离呈极显著正相关(R2=0.568 6,P=0.002)。
UPGMA聚类分析结果(图1)显示:在遗传距离0.51处,供试的青榨槭8个天然种群被分成2个集群,其中,P1种群、P2种群、北京天池峡谷(P3)种群、P4种群、河南嵩山地质公园(P5)种群和陕西南五台地质公园(P6)种群聚为一个集群,P7种群和P8种群聚为另一个集群。
使用STRUCTURE 2.3.4软件对青榨槭8个天然种群的遗传结构进行分析,结果(图2)显示:K=2时,ΔK值最大,遗传结构最清晰,说明供试青榨槭8个天然种群可划分为2个集群。

表4 青榨槭8个天然种群间的遗传相似系数和遗传距离

P1: 北京白羊沟自然风景区Baiyanggou Natural Scenic Area of Beijing; P2: 北京三岔万涧Sancha Wanjian of Beijing; P3: 北京天池峡谷Tianchi Canyon of Beijing; P4: 北京黄柏寺村Huangbaisi Village of Beijing; P5: 河南嵩山地质公园Songshan Mountain Geopark of Henan; P6: 陕西南五台地质公园Nanwutai Geopark of Shaanxi; P7: 广东南岭国家森林公园Nanling National Forest Park of Guangdong; P8: 浙江西天目山森林公园West Tianmu Mountain Forest Park of Zhejiang.

P1: 北京白羊沟自然风景区Baiyanggou Natural Scenic Area of Beijing; P2: 北京三岔万涧Sancha Wanjian of Beijing; P3: 北京天池峡谷Tianchi Canyon of Beijing; P4: 北京黄柏寺村Huangbaisi Village of Beijing; P5: 河南嵩山地质公园Songshan Mountain Geopark of Henan; P6: 陕西南五台地质公园Nanwutai Geopark of Shaanxi; P7: 广东南岭国家森林公园Nanling National Forest Park of Guangdong; P8: 浙江西天目山森林公园West Tianmu Mountain Forest Park of Zhejiang.
3 讨论和结论
3.1 青榨槭天然种群的遗传多样性
相关研究发现,与青榨槭同属的庙台槭(A.miaotaienseP. C. Tsoong.)(Shannon’s多样性指数为0.290 9,Nei’s基因多样性指数为0.193 3)[14]、五小叶槭(A.pentaphyllumDiels)(Shannon’s多样性指数为0.392 4,Nei’s基因多样性指数为0.249 8)[15]和五角枫〔A.pictumsubsp.mono(Maxim.) H. Ohashi〕(Shannon’s多样性指数为0.568 6,Nei’s基因多样性指数为0.389 2)[16]的遗传多样性普遍较高。本研究中青榨槭8个天然种群的基因多样性也较高,Shannon’s多样性指数均值为0.512 7,Nei’s基因多样性指数均值为0.271 9,明显高于庙台槭和五小叶槭的相应指标,但却低于五角枫的相应指标。槭属植物的遗传多样性水平较高可能与物种分布范围广有关[17]53,[18]。青榨槭是分布于华北、华东、中南和西南各省份的广布种,地理分布范围广和风媒传播的特性可能是其遗传多样性水平较高的重要原因[17]53,[18]。
本研究中,广东南岭国家森林公园种群的遗传多样性水平最高,河南嵩山地质公园种群的遗传多样性水平最低。植物种群的遗传多样性可能与自然环境中种群的大小或个体密度等有关[17]5,[18]。笔者在野外调查时发现,广东南岭国家森林公园种群中青榨槭个体数量最多,且该种群的人为干扰较少,生境完整;而河南嵩山地质公园种群中青榨槭个体数量较少,受人为干扰影响,该种群的生境遭到严重破坏,种群内的青榨槭个体呈片段化分布状态。据此推测种群个体数量多、人为干扰较少、生境完整是广东南岭国家森林公园种群遗传多样性高的重要原因。值得注意的是,本研究调查的青榨槭各天然种群的生境总体上受人为干扰较大,其天然种群呈片段化分布,种群规模较小。研究发现,种群片段化分布会迫使植物进行自交繁殖,而自交和近交繁殖势必导致植物的遗传多样性下降[18]。据此认为,青榨槭天然种群的遗传多样性有下降风险,应引起人们的重视。
3.2 青榨槭天然种群的遗传分化和遗传结构
本研究结果显示:青榨槭的遗传分化主要存在于种群内(种群内遗传变异贡献率为66.55%),而种群间的遗传分化相对较小(种群间遗传变异贡献率为33.45%),青榨槭的种群间遗传变异贡献率与同属植物庙台槭(41.54%)[14]和五小叶槭(37.22%)[15]的种群间遗传变异贡献率相近。本研究还发现,青榨槭种群间基因流仅0.508 8,明显小于l,说明供试青榨槭8个天然种群间的基因交流较少。上述研究结果均可能与各青榨槭天然种群所处的地形地貌差异或地理距离较远有关。供试青榨槭8个天然种群间相距35~1 792 km,种群地分属于秦岭山脉、太行山山脉、燕山山脉和南岭山脉,除太行山山脉与燕山山脉相连,其余山脉均片段化分布,据此推断地理屏障也是造成青榨槭天然种群间基因交流困难的主要原因,导致青榨槭天然种群间的遗传分化较为严重。另外,槭属植物还存在花粉流不同期、种群间种子流难形成的问题[14-16],也会造成种群间的基因交流较少,导致青榨槭的遗传分化主要存在于种群内的个体间。总体来看,供试青榨槭8个天然种群的观测杂合度小于期望杂合度,表明供试青榨槭天然种群内存在一定程度的自交或近交现象。
综合聚类和遗传结构分析结果,供试青榨槭8个天然种群被分成2个集群,即广东南岭国家森林公园种群和浙江西天目山森林公园种群为一个集群,其余6个种群为另一个集群。Mantel检验结果表明:供试青榨槭8个天然种群间的遗传距离与地理距离呈极显著正相关。
3.3 青榨槭天然种群的保护策略
上述研究结果表明:虽然青榨槭天然种群的遗传多样性水平较高,但是各种群内均存在一定程度的自交或近交现象,导致种群逐渐衰退,因此,应加强对青榨槭天然种群的保护力度。相关部门既要重视对青榨槭各天然种群内不同个体的保护,又要注重对不同种群的保护。广东南岭国家森林公园种群的遗传多样性水平最高,建议优先保护。当然,还要对各天然种群进行适当的人工干预(如人为促进种群间种子与花粉的传播),以减少种群内植株的自交和近交现象。此外,还可以收集不同天然种群的种质资源,选择合适地点构建种质圃;开展青榨槭的引种栽培研究,使栽培资源能够满足市场需求,从而减少其野生资源被破坏。
猜你喜欢
儿童故事画报·自然探秘(2021年12期)2021-08-16
智慧健康(2021年17期)2021-07-30
种子(2021年3期)2021-04-12
华人时刊(2020年21期)2021-01-14
科技传播(2019年24期)2019-06-15
新课程·下旬(2018年9期)2018-11-14
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
剑南文学(2016年14期)2016-08-22
中学生理科应试(2016年7期)2016-05-14
