
2020年天津近岸海域鱼类群落结构及多样性
2022-05-20 09:02梁鹏飞张树林张达娟王泽斌毕相东
广东海洋大学学报 2022年3期
梁鹏飞,张树林,张达娟,李 琦,王泽斌,戴 伟,毕相东
2020年天津近岸海域鱼类群落结构及多样性
梁鹏飞,张树林,张达娟,李 琦,王泽斌,戴 伟,毕相东
(天津农学院水产学院 / 天津市水产生态及养殖重点实验室,天津 300392)
【】调查天津近岸海域鱼类资源现状,了解该海域鱼类种类组成、群落和生物多样性的变化规律,利用底拖网于2020年4、6-10月逐月在天津近海进行6航次鱼类资源调查,分析种类组成、数量分布,运用聚类、RDA分析、ABC曲线等方法分析天津近海鱼类群落结构现状。【2020年共捕获鱼类21种,隶属于4目11科17属,生态类型以暖温性、大陆架浅水底层鱼类为主。年优势种主要以小型鱼类为主,包括斑鰶()、短吻红舌鳎()、矛尾虾虎鱼()和六丝矛尾虾虎鱼()。鱼类年平均丰度和单位捕捞努力量(CPUE)分别为588.48尾/(网·h)、4.79 kg /(网·h),渔获物的相对质量中斑鰶、短吻红舌鳎和矛尾虾虎鱼占比较高,分别占总渔获量的37.83%、20.84%和11.84%。鱼类群落年平均多样性指数、丰富度指数、均匀度指数分别为1.93、1.81和0.53。聚类分析表明,在59%相似性水平上,鱼类群落按丰度分为A、B和C三组,其中斑鰶和短吻红舌鳎在组间相异性贡献率最高;RDA分析表明,温度变化是构成群落结构变化的重要因素;ABC曲线表现为丰度曲线位于生物量曲线上方(= -0.038 < 0),表明鱼类群落受外界扰动影响较大。
鱼类资源;群落结构;优势种;天津近岸海域;ABC曲线
天津位于渤海西岸,其近岸海域营养盐丰富,历史上是鱼虾的重要索饵场[1-2]。近年来,天津近岸海域富营养化程度不断提高[4],渤海生态稳定性转差,鱼类群落结构发生变化[5]。鱼类是海洋游泳动物的重要组成部分,对海洋环境剧烈变化及过度捕捞产生一系列响应。自20世纪80年代开始,渤海鱼类资源密度降低,资源量大幅下降,鱼类群落正在向小型化、低龄化、低质化转变,经济价值高的优质种类逐渐减少[6-7],莱州湾鱼类群落长期受外界干扰,群落多样性不断下降[8],严重阻碍了渔业资源的可持续发展。天津是渤海重要组成部分,已有天津近岸海域鱼类群落结构和渔业资源的调查研究[9-10]。张青田等[11]指出,天津海域鱼类群落结构有明显的月变化,3、4月鱼类种类明显少于其他月份,季节性变化显著。王泽斌等[12]还指出,鱼类种类组成有较高月间更替率,月平均更替率高达110.0%。为进一步掌握天津近海鱼类资源现状,笔者于2020年4、6‒10月对该海域进行拖网调查,分析该海域鱼类种类组成及群落变化,为该海域渔业资源评估和养护提供参考。
1 材料与方法
1.1 调查区域与调查方法
于2020年4、6-10月于天津近岸海域(117°40′―118°00′ E,38°30′―38°50′ N)9个站位(图1),依据《海洋调查规范》[13],共进行6航次调查采样,调查海域面积为693 km2。原计划进行8航次调查研究,由于受3月天气影响、5月禁渔期影响,因此实际完成6航程调查。
使用“津港渔05006”渔船,采样单船底拖网采捕,船只功率110.3 kW,囊网网目尺寸20 mm,网口宽8 m,上纲长度为44 m。每站拖网1 h,拖速2 kn,扫海面积共0.03 km2/h。渔获物鉴定参考《黄渤海鱼类图志》[14],渔获物生态类型划分参考刘静等[15]、王泽斌等[16]方法。用溶氧仪、pH检测仪、氧化还原电位自动测定仪、盐度计、温度计分别现场测定溶解氧(DO)、pH、氧化还原电位(ORP)、盐度()和温度()。
1.2 鱼类群落多样性分析
根据每一站位拖网时间,对鱼类调查数据进行标准化处理[17],换算为单位时间生物量 [ kg / (网·h)]和丰度 [ 尾 / (网·h)]。
计算生态优势度[18]、Shannon-Wiener多样性指数(′)[19]、Margale丰度指数()[20]、Pielou均匀度指数()[21]。
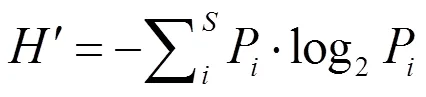
= (– 1) / log2,
=′ / log2,
式中,为样品中的物种总数;P为样品中第种的丰度与各站位总丰度()比值。
用Pinkas相对重要性指数(IRI)确定每种鱼在群落中优势度:IRI(W + N)××104。式中,W为某一种类的生物量占总生物量的百分比;为某一种类的数量占总数量的百分比;为某一种类出现站数占调查总站数的百分比。IRI大于10 000时为绝对优势种,IRI为1 000 ~ 10 000时为优势种。
1.3 鱼类群落结构特征
用等级聚类分类法分析鱼类群落结构特征。将调查月所有数据用Excel 2010整合。由于稀少种类对群落结构分析产生一定影响,需对原始数据进行二次方根转换,再进行Bray-Curtis相似性计算[22-23]。
Bray-Curtis相似性系数()矩阵公式:
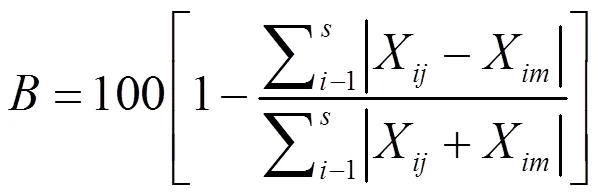
式中,为种类数,X、X分别为第种类在第、个月的平均单位网次渔获量。统计分析及图件绘制用PRIMER 6和Excel2010完成。
1.4 鱼类群落结构与环境因子的相关性分析
运用Canoco4.5软件的冗余分析(RDA)法分析鱼类与环境因子之间的关系,解释各环境因子对鱼类分布的影响。对物种进行趋势对应分析(DCA),根据每个轴的梯度长度(LGA) 选择适宜的排序方法。当LGA﹤3,采用冗余分析;LGA﹥4,采用典范对应分析;3≤LGA≤4,两种分析方法均可采用[24]。
1.5 群落稳定性分析
用丰度/生物量比较法(ABC曲线)分析。ABC曲线是根据丰度优势度曲线与生物量优势度曲线的变化情况以及相对位置判断群落的变化状况,两条曲线与坐标轴围成的面积为值[25]。
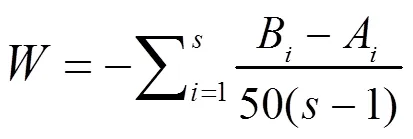
式中,A和B分别为第种物种对应的丰度、生物量累计百分比,为鱼类的总种类数。
判断依据为:整条丰度优势度曲线位于生物量优势度曲线的下方,值为正值,表明群落稳定,未受到干扰;整条丰度优势度曲线位于生物量优势度曲线的上方,值为负值,表明群落处于不稳定、受到严重干扰的状态。
2 结果与分析
2.1 种类组成及生态类型
调查共捕获鱼类21种,隶属于4目11科17属。其中鲈形目鱼类14种,占66.7%;鲱形目鱼类4种,占19.0%;鲽形目鱼类2种,占9.5%;鲉形目鱼类1种,占4.8%。鱼类组成以暖温性、大陆架浅水底层鱼类为主(表1)。

表1 天津近岸海域鱼类种类组成及生态类型
注:HT,栖所类型;ST,适温类型;CD,大陆架浅水底层鱼类;CBD,大陆架浅水中底层鱼类;CPN,大陆架浅水中上层鱼类;CRA,大陆架岩礁性鱼类;OEP,大陆架大洋洄游性中上层鱼类;WT,暖温性;WW,暖水性;CT,冷温性。
Notes: HT, habitat type; ST, suitable temperature type; CD, continental shelf demersal fish; CBD, continental shelf benthopelagic fish; CPN, continental shelf pelagic neritic fish; CRA, continental shelf reef-associated fish; WT, warm temperate species; WW, warm water species; CT, cold temperate species.
2.2 优势种变化
2020年鱼类年优势种有4种,分别为斑鰶、短吻红舌鳎、矛尾虾虎鱼、六丝矛尾虾虎鱼。鱼类各月的优势种共有6种,矛尾虾虎鱼、短吻红舌鳎、斑鰶分别为4、6、7月绝对优势种(IRI﹥10 000)(表2)。各月鱼类优势种的丰度占当月总丰度的77.4% ~ 96.6%,鱼类优势种生物量占当月总生物量的69.8% ~ 97.6%,可见天津近岸海域优势类群主要以小型鱼类为主。
2.3 丰度和生物量变化
从时间分布看,2020鱼类平均丰度为588.48 尾/(网·h),7月最高,为1 385.0 尾/(网·h);4月最低,为215.6 尾/(网·h)。鱼类平均生物量为4.79 kg/(网·h),7月最高,为9.06 kg/(网·h);4月最低,为2.34 kg/(网·h)。从空间分布看,鱼类丰度在A4站位丰度最低,为347.0 尾/(网·h);在A3站位最高,为880.8尾/(网·h)。鱼类生物量在A4站位最低,为2.27 kg/(网·h);在A6站位最高,为7.03 kg/(网·h)(图2)。渔获物的相对质量中斑鰶、短吻红舌鳎和矛尾虾虎鱼占比较高,分别占总渔获量的37.83%、20.84%和11.84%。

表2 鱼类优势种组成
2.4 群落结构
2.4.1 物种多样性 2020年多样性指数′、丰富度指数和均匀度指数的平均值分别为1.93、1.81和0.53。各月多样性指数′ 为1.47 ~ 2.71,9月最高,7月最低。各站位多样性指数′ 为1.63 ~ 2.55,A3站位最低,A4站位最高。
2.4.2 群落结构的聚类分析 图3表明,在59%的相似性水平上,各月鱼类群落可分为A、B、C三组。SIMPER分析表明,典型种对C组累计相似贡献率为90.59%,斑鰶相似性贡献率最高,为31.64%;分歧种对A组和B组累计相异性贡献率为92.01%,短吻红舌鳎相异性贡献率最高,为38.93%;分歧种对A组和C组累计相异性贡献率为91.9%,斑鰶相异性贡献率最高,为29.54%;分歧种对B组和C组累计相异性贡献率为91.21%,其中短吻红舌鳎和斑鰶相异性贡献率分别为24.12%、22.71%(表3)。
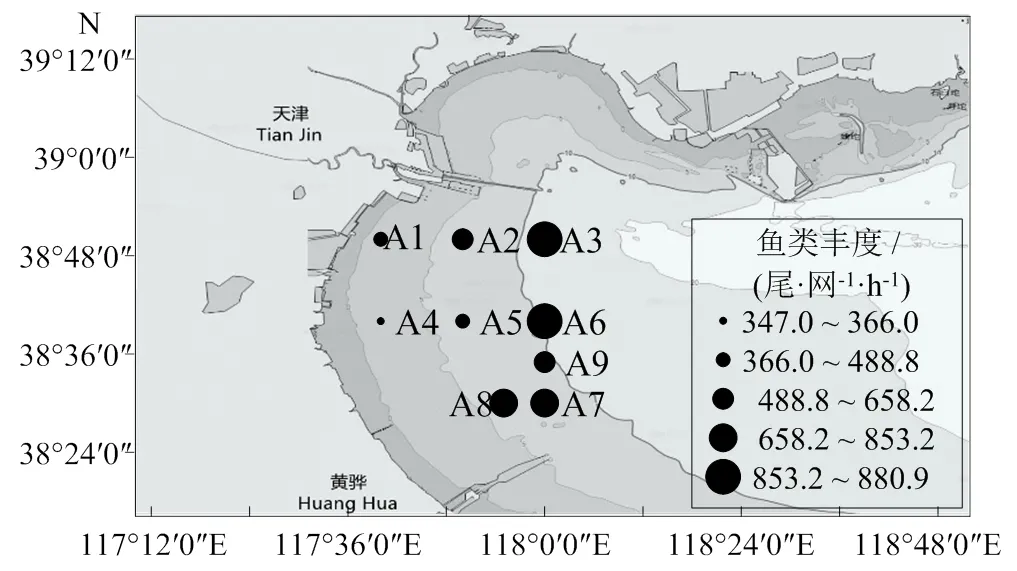
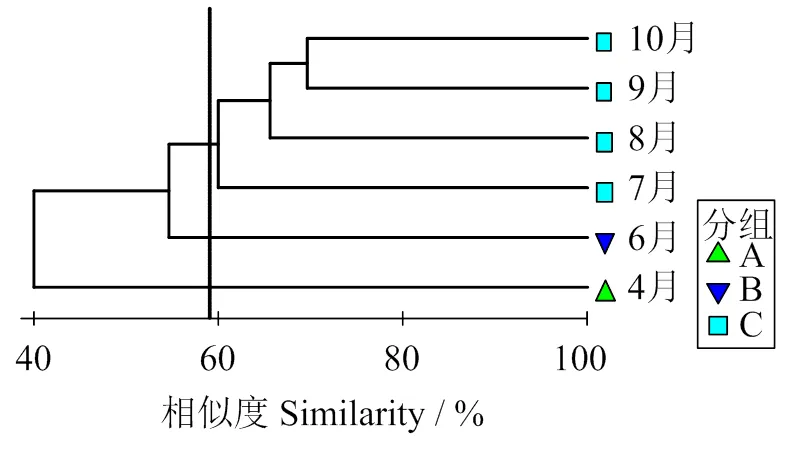
图3 鱼类聚类分析
2.4.3 鱼类分布与环境因子的关系 根据DCA分析结果,3 < LGA = 3.06 < 4,单峰模型(CCA) 和线性模型(RDA)均可使用,本研究选择RDA分析。轴1和轴2的特征值分别为0.258、0.098,分别解释数据方差变化的25.80%和9.80%。蒙特卡罗检验表明,除盐度外,其余环境因子排序轴均有显著的相关性(< 0.05)。其中解释量最大为水温,解释了变异的24.5%,其次为DO、ORP、pH、盐度,分别解释14.8%、12.9%、10.9%、1.4%(图4)。
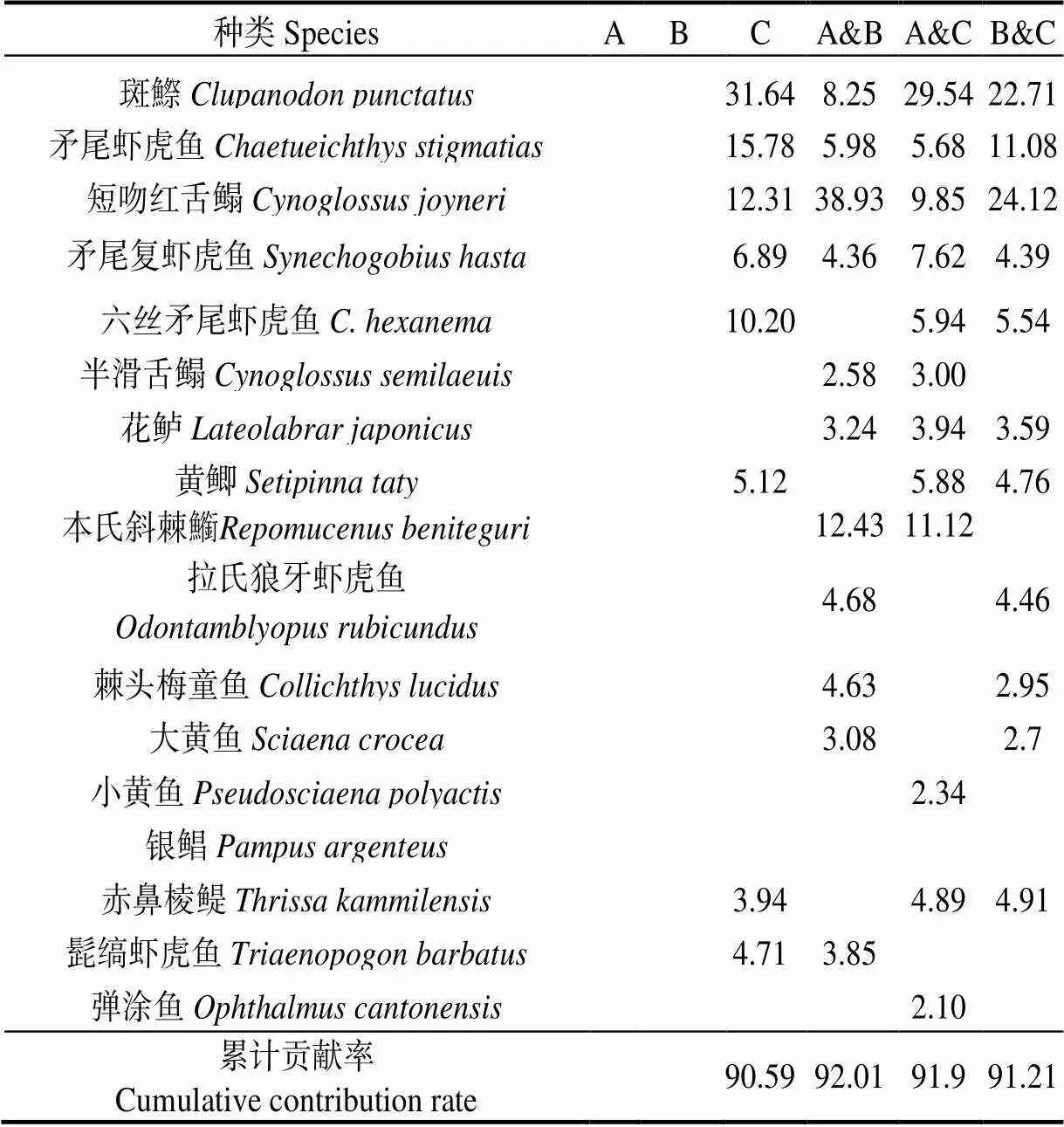
表3 鱼类组间相似性和差异性贡献率
2.4.4 鱼类群落结构的稳定性 由图5 可知,2020年丰度优势度曲线位于生物量优势度曲线上方,小于0 (–0.038),表明鱼类群落受到了较为严重干扰。图6显示,6月、7月和8月丰度优势度曲线在生物量优势度曲线上方,表明群落处于不稳定状态,受到严重干扰;4月和10月丰度优势度曲线在生物量优势度曲线的下方表明群落结构稳定,未受到干扰。9月曲线出现一定交叉,但交叉程度不大,表明该月份鱼类群落结构受到轻微干扰。
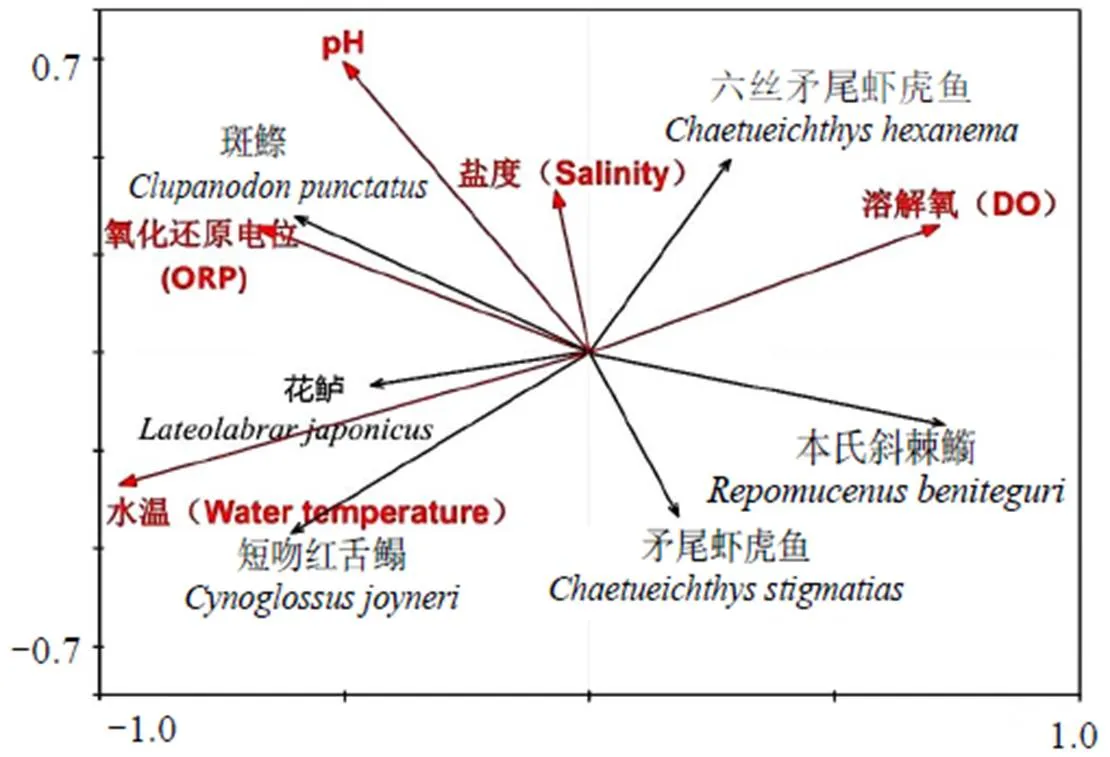
图4 优势种与环境因子RDA分析
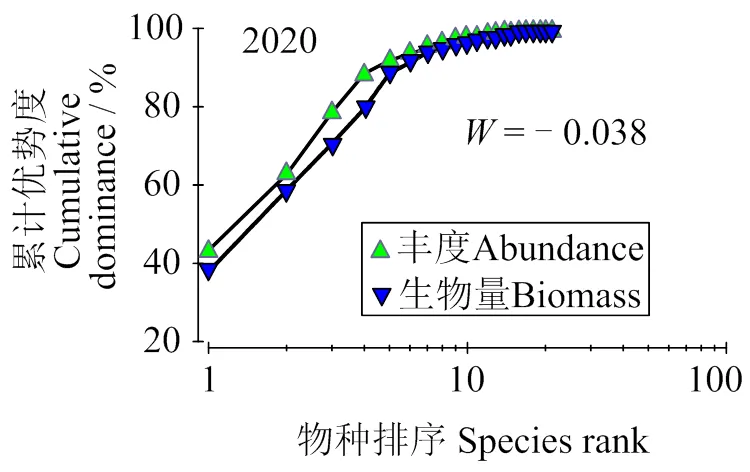
图5 2020年天津近岸海域鱼类资源ABC曲线
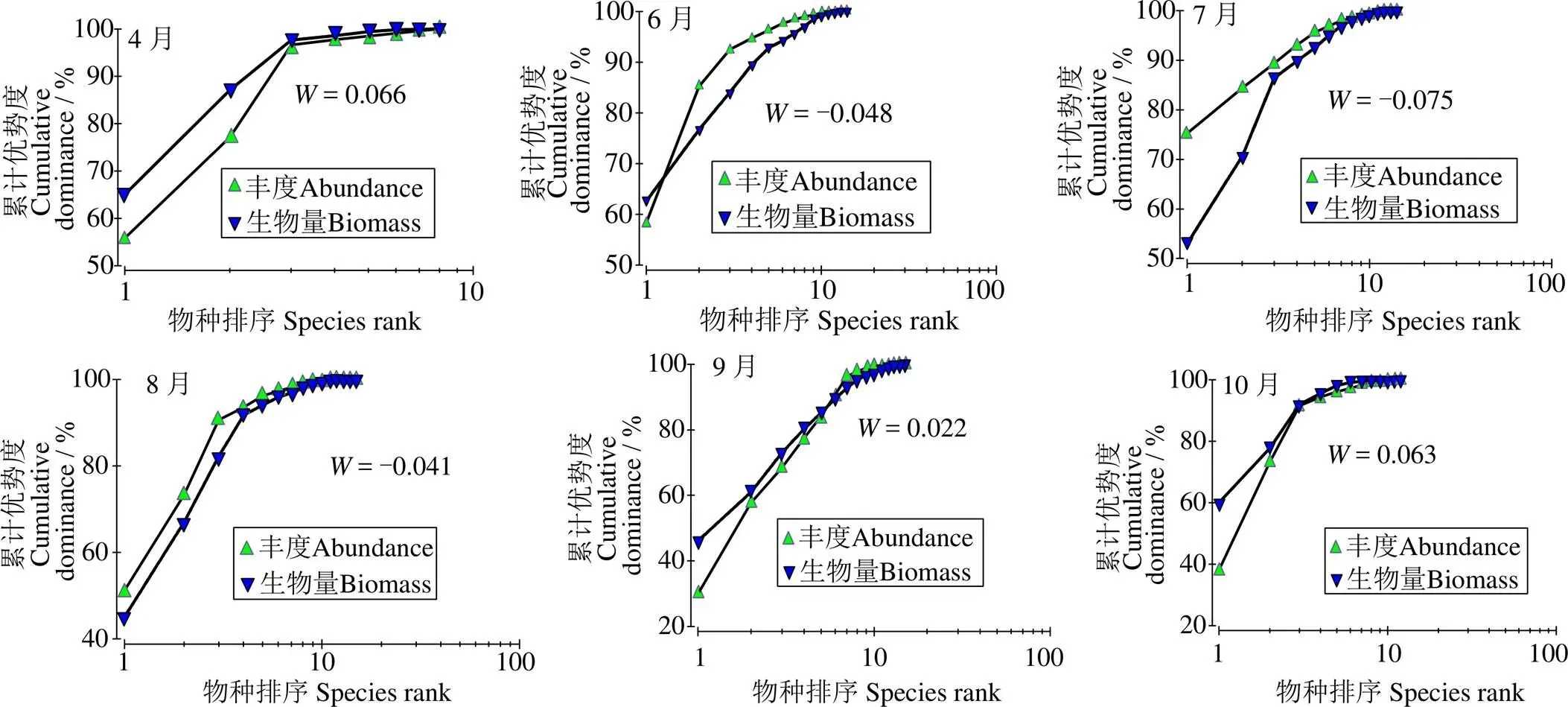
图6 2020年天津近岸海域鱼类月份ABC曲线
3 讨论
3.1 天津近岸海域鱼类种类及优势种变化
20世纪80年代对天津市海洋综合调查中共捕获鱼类50种,其中优势种为半滑舌鳎、斑鰶、黑鳃梅童鱼()、刀鲚()、黄鲫()等[26]。本调查鱼类总数量较20世纪80年代减少29种,优势种由经济价值高的半滑舌鳎、刀鲚转变为经济价值低的矛尾虾虎鱼、短吻红舌鳎。优势种出现明显的更替现象,经济鱼类群落结构正向小型化、低龄化转变,这与整个渤海鱼类变化情况一致。其原因一方面可能与所选站位有关,本调查的站位设置较20世纪有所减少,调查范围缩小。另一方面是受人类经济活动影响,大量污染物随入海径流进入渤海湾,而渤海湾属于半封闭海湾,交换能力差,因此整个海域受到不同程度污染;同时过度捕捞也加海洋渔业资源的衰退。2013和2014年共捕获鱼类26种[10];2017、2018年分别捕获鱼类20、23种,主要优势种六丝矛尾虾虎鱼、矛尾虾虎鱼、花鲈、短吻红舌鳎等[12,16]。天津近岸海域近几年拖网调查鱼类种类无明显变化,优势种趋于稳定,得益于近年来海洋法的制定和休渔制度的执行。
3.2 天津近岸海域鱼类群落结构变化
群落多样性指数是反映鱼类群落结构重要指标,一般为1.5 ~ 3.5[27]。本调查各月及站位多样性指数均在此范围之内。从时间角度来看,2020年多样性指数′ = 1.93,与2017(′ = 1.94)、2018年(′ = 2.04)相比无波动变化[28],表明近年来群落多样性相对稳定。多样性指数最高出现在9月(′ = 2.71),其次为8月(′ = 1.99),其原因为随着月份增加,鱼类种类数分别在8月和9月最高。9月,禁渔期结束,人工捕捞使鱼类优势种丰度下降,生物量占比趋于相对均匀,多样性指数′达到最大。8月多样性指数较9月低的原因为禁渔期前鱼类丰度和生物量高度集中,均匀度较低。最低值出现在7月,其原因为7月单一种类斑鰶高度集中,造成优势种与非优势种丰度、生物量差距较大,其中斑鰶丰度占比75.26%,因此7月多样性指数在所调查月中最低。从空间角度来看,′分别在A3站位达到最低值,在A4站位达到最高值。其原因为A3站位全年捕获鱼类种类数较少,但渔获量较大,导致均匀度较低,多样性指数最低。A4站位鱼类种类数较高,全年丰度和生物量均匀,因而多样性指数最高。
多样性指数无法反映具体物种对群落影响,多元分析可弥补这一不足。本研究通过聚类分析,将各月鱼类群落按丰度分为A、B、C三个时间组群,典型种对C组相似贡献率最高为斑鰶,斑鰶在7-10月均为优势种,且在7-10月渔获量占比较大,因此7-10月群落结构较为相似。分歧种对A、B组相异性贡献率中最高为短吻红舌鳎,A组(4月)与B组(6月)共同优势种为矛尾虾虎鱼和六丝矛尾虾虎鱼,主要差异种为短吻红舌鳎。分歧种对B(6月)、C组(7-10月)相异性贡献率中最高为短吻红舌鳎和斑鰶,4月短吻红舌鳎成为绝对优势种,在7-10月短吻红舌鳎虽为优势种,但在当月占比相对较小,所以B(6月)和C组(7-10月)短吻红舌鳎虽均为优势种,但群落存在差异。B组斑鰶6月未成优势种,分别在7-10月成为优势种。因此,丰度分布差异导致各月群落结构出现差异。
3.3 鱼类群落与环境因子关系
鱼类群落结构变化与温度、盐度和水深等环境因子有关[29],温度变化是鱼类群落结构变化的主要诱因[30]。由于本调查区域集中在近岸海域,因而未分析水深等空间环境因子。本调查中,天津近岸海域以冷温性、暖水性和暖温性鱼类为主,其中暖温性鱼类占60%,占据绝对优势。各月优势种丰度与环境因子的RDA分析表明,除盐度外,其余环境因子与优势种丰度均显著相关(< 0.05),其中温度解释量最大,且斑鰶、花鲈、短吻红舌鳎的丰度与温度呈正相关。斑鰶分布受温度影响较大,20.7 ~ 22.7 ℃为斑鰶孵化最适宜温度[31-32]。本调查中,从6至10月均可捕获斑鰶,7至10月斑鰶为优势种,7月为绝对优势种。因此,温度是影响群落结构的主要因素。
3.4 鱼类群落的稳定性
ABC曲线可反映鱼类群落受干扰程度及生态环境对鱼类的影响,广泛用于渔业资源调查[33-34]。本研究中,2020年丰度曲线在生物量曲线上方(= –0.039),表明群落受到严重干扰。其原因为天津近岸海域鱼类群落主要以经济价值低、规格小、年龄结构简单的小型鱼类为主。6-8月丰度曲线在生物量曲线上方,表明群落处于不稳定状态。这可能与禁渔期和环境月变化有关。由于6-8月为禁渔期,海水温度不断升高,浮游植物和浮游动物数量不断增加,为鱼类生长繁育提供了有利条件,鱼类数量不断增加,造成鱼类丰度大于生物量,群落不稳定。4、10月丰度曲线在生物量曲线下方,表明鱼类群落处于稳定状态。这是因为4月温度较低,鱼类种类较少,丰度、生物量较低,群落结构稳定。9-10月禁渔期结束后大量海上作业使得近岸海域优势种数量下降,物种丰度和生物量趋于均匀。但9月丰度曲线出现了一定的交叉,但交叉程度不大,表明9月鱼类群落结构受到轻微干扰。这主要由于人工捕捞无固定地点,区域捕捞不均匀,导致9月拖网捕捞的丰度和生物量发生变化。
[1] 张闰生, 孙秋岩. 天津市海岸带和海涂资源综合调查报告(海洋生物部分)[R]. 天津: 天津新闻出版管理局, 1986: 1-91.
[2] 王麒麟, 宓慧菁, 谷德贤, 等. 天津海域重要渔业资源产卵场生态环境初步研究[J]. 河北渔业, 2014(9): 15-17.
[3] 周滨, 孙丽娜, 王秋莲, 等. 天津市近岸海域氮、磷入海总量核算初探[J]. 海洋环境科学, 2021, 40(6): 851-859.
[4] 阚文静, 张亚楠, 张秋丰, 等. 天津近岸海域水体富营养化特征及健康状况分析[J]. 海洋湖沼通报, 2013(3): 147-151.
[5] 张波, 吴强, 金显仕. 莱州湾鱼类群落的营养结构及其变化[J]. 渔业科学进展, 2013, 34(2): 1-9.
[6] 金显仕. 渤海主要渔业生物资源变动的研究[J]. 中国水产科学, 2000, 7(4): 22-26.
[7] 冯昭信, 李久奇. 黄、渤海鱼类资源之现状与管理意见[J]. 海洋科学, 1990, 14(3): 60-63.
[8] 杨艳艳, 朱明明, 徐炳庆, 等. 莱州湾鱼卵、仔稚鱼群落结构及多样性的季节变化[J]. 海洋学报, 2021, 43(9): 59-70.
[9] 张秋丰, 徐玉山, 刘森. 天津市近岸海域海洋生态文明建设初探[J]. 海洋经济, 2016, 6(6): 29-36.
[10] 谷德贤, 刘国山, 王晓宇, 等. 基于GAM模型的天津海域鱼类资源和环境因子关系的初步研究[J]. 天津农学院学报, 2017, 24(1): 38-43.
[11] 张青田, 胡桂坤. 天津近海鱼类组成和分类学多样性的月变化[J]. 海洋湖沼通报, 2017(1): 133-140.
[12] 王泽斌, 张树林, 张达娟, 等. 天津近海鱼类月间种类组成及变化[J]. 大连海洋大学学报, 2019, 34(5): 733-738.
[13] 国家质检总局, 国家标准化委员会. 海洋调查规范第6部分: 海洋生物调查[M]. 北京: 中国标准出版社, 2007: 56-62.
[14] 刘静, 陈咏霞, 马琳. 黄渤海鱼类图志[M]. 北京: 科学出版社, 2015.
[15] 刘静, 宁平. 黄海鱼类组成、区系特征及历史变迁[J]. 生物多样性, 2011, 19(6): 764-769.
[16] 王泽斌, 张树林, 张达娟, 等. 天津近海鱼类群落结构及功能群组成初步研究[J]. 海洋科学, 2019, 43(9): 78-87.
[17] SC/T 9110-2007 建设项目对海洋生物资源影响评价技术规程[S].
[18] PINKAS L, OLIPHAMT M S, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. California Department of Fish and Game Fish Bulletin, 1971, 152: 1-105.
[19] SHANNON C E, WEAVER W. A Mathematical Theory of Communication[J]. Addison-Wesley Pub, 1981, 97(3): 29-125.
[20] ULANOWICZ R E. Information theory in ecology[J]. Computers & Chemistry, 2001, 25(4): 393-399.
[21] PIELOU E C. Species-diversity and pattern-diversity in the study of ecological succession[J]. Journal of Theoretical Biology, 1966, 10(2): 370-383.
[22] CLARKE K R, WARWICK R M. Change in marine communities: an approach to statistical analysis and interpretation[M]. [S. l.]: Plymouth marine laboratory, Natural environment research council, 1994.
[23] 周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报(自然科学版), 2003, 33(1): 58-64.
[24] 樊紫薇, 蒋日进, 张洪亮, 等. 岱衢洋产卵场鱼卵、仔稚鱼群落结构及其与环境因子的关系[J]. 水产学报, 2020, 44(4): 606-620.
[25] CLARKE K R, WARWICK R M. Change in marine communities: an approach to statistical analysis and interpretation[M]. s. l. : Plymouth marine laboratory, Natural environment research council, 1994.
[26] 天津市海岸带和海涂资源综合调查组. 天津市海岸带和海涂资源综合调查报告第八篇: 海洋生物[M]. 北京: 海洋出版社, 1991.
[27] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ. α多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 31-239.
[28] 王泽斌. 围填海工程对天津近海鱼类资源的影响及评估[D]. 天津: 天津农学院, 2019.
[29] 张静, 邱俊文, 陈春亮, 等. 广东大鹏湾北部海域春秋季鱼类群落结构及其与环境因子关系[J]. 广东海洋大学学报, 2020, 40(6): 43-52.
[30] 于南京, 俞存根, 许永久, 等. 舟山群岛外海域春秋季鱼类群落结构及生物多样性[J]. 水产学报, 2021, 45(8): 1374-1383.
[31] 万瑞景, 姜言伟. 渤、黄海硬骨鱼类鱼卵与仔稚鱼种类组成及其生物学特征[J]. 上海水产大学学报, 2000, 9(4): 290-297.
[32] BREIMAN L. Random forests[J]. Machine Learning, 2001, 45(1): 5-32.
[33] 刘前, 王学锋, 吕少梁, 等. 湛江港海域游泳动物群落结构及多样性分析[J]. 广东海洋大学学报, 2021, 41(2): 103-110.
[34] 俞松立, 王咏雪, 韩晓凤, 等. 三门湾海域渔业生物群落结构及营养级变化特征[J]. 大连海洋大学学报, 2021, 36(3): 478-487.
Fish Community Structure and Diversity in Coastal Area of Tianjin in 2020
LIANG Peng-fei, ZHANG Shu-lin, ZHANG Da-juan, LI Qi, WANG Ze-bin, DAI Wei, BI Xiang-dong
(,/,300392,)
【】To investigate the fish stock in the Tianjin coastal waters, and explore the changes of fish species composition, community structure and diversity in this waters. 【】The bottom trawl was used to investigate the fish resources of 6 voyages off the coast of Tianjin in April, from June to October 2020. Based on the analysis of species composition, quantitative distribution, cluster analysis, RDA analysis, ABC curve analysis and other statistical methods were used to explore the current status of fish community structure in Tianjin coastal waters.【】A total of 21 species of fishes were identified, belonging to 4 orders,11 families and 17 genera. And their ecological type was dominated by warm temperate and shallow bottom fish on the continental shelf. The annual dominant species were mainly small fish, including,,andThe average annual abundance and biomass of fish were 588.48 ind/net·h and 4.79 kg/net·h. In relative weight of the cash, the,andaccounted for 37.83%, 20.84% and 11.84% respectively. The average Shannon-Weiner biodiversity index, Margalef abundance index and Pielou evenness index of fish community were 1.93, 1.81 and 0.53 respectively. The cluster showed the fish community were divided into three groups A, B and C at 59% similarity level, among which theandcontributed most to the intergroup dissimilarity. RDA analysis showed that temperature change was an important factor in the change of community structure; The ABC curve showed that the abundance curve was located above the biomass curve (= -0.038 < 0), indicating that the fish community was greatly affected by external disturbance.
fish stock; community structure; dominant species; Tianjin coastal waters; ABC curve
梁鹏飞,张树林,张达娟,等. 2020年天津近岸海域鱼类群落结构及多样性[J]. 广东海洋大学学报,2022,42(3): 18-24.
S932.4
A
1673-9159(2022)03-0018-07
10.3969/j.issn.1673-9159.2022.03.003
2022-01-22
国家自然科学基金项目(32172978);天津市自然科学基金项目(19JCYBJC30000);天津市淡水养殖产业技术体系创新团队-养殖水环境调控岗位(ITTFRS2021000-009);中央引导地方(天津)科技发展专项(21ZYCGSN00500);天津市教委科研计划项目(2020ZD06,2021KJ110);天津市高等学校创新团队基金项目“天津市现代水产生态健康养殖创新团队”(TD13-5089);甘肃省科技计划项目民生科技专项(21CX6NP223)
梁鹏飞(1996―),男,硕士研究生,主要从事海洋生态调查研究。Email:lpengfei96@163.com
张达娟(1981―),女,实验师,博士,研究方向为养殖水环境调控与修复。E-mail:dajuanzhang@163.com
(责任编辑:刘庆颖)
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
农业工程学报(2022年10期)2022-08-22
中国音乐学(2022年2期)2022-08-10
现代园艺(2022年6期)2022-04-08
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
