
碱基切除修复基因OGG1对紫外线诱导晶状体上皮细胞损伤的保护作用△
2022-05-19 02:34李鹏飞罗家伟康丽华张国伟茅馨木吴安然张文怡管怀进
眼科新进展 2022年4期
李鹏飞 罗家伟 康丽华 张国伟 茅馨木 吴安然 张文怡 管怀进
迄今为止,年龄相关性白内障(ARC)的发病机制仍未完全清楚。研究证实,紫外线(UVB)辐射所造成的氧化损伤与ARC的发生发展紧密相关[1]。这种氧化损伤首先累及晶状体上皮细胞(LEC),造成细胞内DNA和细胞膜质子泵受损,从而激活细胞凋亡途径。由于晶状体本身存在一定的损伤修复机制,只有细胞内损伤性DNA累积到一定程度时,才会引起LEC凋亡[2]。因此,DNA的损伤修复能力对LEC的存活至关重要。我们前期研究已证实,8-羟基鸟嘌呤DNA糖苷酶1(OGG1)基因能通过碱基切除修复(BER)通路特异性去除DNA双链中的8-氧鸟嘌呤(8-oxoG),参与受损DNA的修复过程[3]。现有的研究结果显示,OGG1基因的突变与老化和各种退行性疾病的发生密切相关,也可能参与ARC的发生发展过程[4-6]。本研究以人LEC(SRA01/04细胞株)为研究对象,采用UVB照射细胞模拟氧化损伤环境,探讨OGG1对UVB诱导损伤的LEC的保护作用。
1 材料与方法
1.1 材料SRA01/04细胞株购自中国科学院上海生命科学研究所。细胞培养实验用品包括:DMEM培养液、胎牛血清、胰蛋白酶和细胞培养板(均购自美国Gibco公司)。手持UVB检测灯购自上海光豪分析仪器公司;逆转录试剂盒购自美国Thermo Scientific公司,Trizol试剂盒购自美国Invitrogen公司,引物购自中国生工生物工程股份有限公司;靶向OGG1设计的siRNAs购自中国锐博生物科技有限公司;CCK8试剂盒购自日本同仁化学研究所;GAPDH抗体、辣根过氧化物酶标记的山羊抗兔和山羊抗鼠IgG二抗均购自中国ABclonal公司,BAX、BCL-2和15A3抗体均购自英国Abcam公司。
1.2 细胞培养和UVB处理将冻存的SRA01/04细胞复苏后,加入完全培养基(含体积分数10%胎牛血清和10 g·L-1青链霉素的DMEM)进行常规培养。将培养瓶置于37 ℃含体积分数5% CO2细胞培养箱中继续培养。待细胞融合度达80%时,用Hanks液清洗细胞2次,加入适量的无菌PBS,置于紫外灯下30 cm处进行照射。将细胞分为UVB组和对照组,UVB照射时间分别为0 min(对照组)、10 min、15 min、30 min、45 min。照射后,加入完全培养基,继续培养24 h后进行后续实验。
1.3 实时荧光定量-PCR检测根据Trizol试剂盒说明书的操作步骤提取本实验的总RNA。按照说明书设定逆转录反应体系和反应时间。以GAPDH为内参,比较各组细胞目的基因和相应内参的Ct值,采用2-△△Ct分析目的基因表达情况,实验均重复3次。OGG1引物序列:上游为5’-TCCCGGTCTTCCTGATTAGC-3’,下游为5’-TACAAGAACTGGAGCACCGT-3’;GAPDH引物序列:上游为5’-TGAAGGTCGGAGTCAACGGATTTGGT-3’,下游为5’-CATGTGGGCCATGAGGTCCACCAC-3’。
1.4 细胞转染及分组利用siRNA敲降技术针对靶基因OGG1设计3个siRNA(分别为siOGG1#1、siOGG1#2和siOGG1#3)。同时,为了验证出敲降效率最高的siOGG1,依据转染情况将细胞分为siOGG1#1组、siOGG1#2组、siOGG1#3组、siNC组和空白对照组(不转染),实时荧光定量-PCR检测各组细胞中OGG1 mRNA的相对表达量。后续为了验证OGG1的表达下降对氧化损伤环境下细胞的作用,将细胞分为空白对照组、UVB组、UVB+siNC组和UVB+siOGG1#2组,观察各组细胞凋亡相关蛋白的表达变化。具体实验方法如下:首先,将细胞均匀接种至6孔板内,待细胞密度合适后,每孔中加入Lipofectamine 3000和siRNA充分混匀,室温孵育15 min。之后,放入培养箱中孵育细胞6 h后,将细胞培养基更换为含有血清的全培养基,并置于培养箱中继续培养48 h,采用UVB照射细胞,照射后继续培养24 h,提取蛋白,免疫印迹实验检测各组细胞中OGG1蛋白表情况。
1.5 免疫荧光法检测细胞DNA氧化损伤转染siOGG1#2后经UVB照射,观察细胞内DNA氧化损伤程度的变化情况,检测方法参考前期研究[7]。将细胞接种至细胞培养板,根据实验设计分为UVB+ siNC组和UVB+siOGG1#2组,之后对细胞进行siRNAs靶向干预和UVB处理。孵育24 h后,用PBS清洗细胞。随后加入40 g·L-1多聚甲醛,室温固定30 min。配制30 g·L-1牛血清白蛋白+体积分数0.5%Triton X-100溶液,室温封闭2 h。之后,根据抗体15A3(染色反应底物是8-oxoG,它是OGG1的修复作用靶点)说明书的比例用10 g·L-1牛血清白蛋白配制一抗反应液,置于4 ℃冰箱内孵育过夜。次日,洗涤后避光,加入配置好的荧光二抗,室温下孵育2 h。加Hoechst(110 000)避光孵育8 min染细胞核。最后,采用荧光显微镜避光拍摄。
1.6 CCK8实验检测细胞活力将细胞接种于96孔板,每孔10×103个细胞,置于培养箱培养24 h;根据实验需要将细胞分为空白对照组、UVB组、UVB+siNC组以及UVB+siOGG1#2组,每组设置6个平行孔,弃培养基,向每孔加入100 μL完全培养基和10 μL CCK8溶液;继续置于培养箱孵育1~2 h(操作过程注意避光);使用酶标仪测定样品在波长450 nm处的吸光度。
1.7 免疫印迹实验检测蛋白表达从培养箱中取出已处理过的细胞,加入蛋白裂解液(CM/RIPA,1/100),冰上裂解,提取上清;BCA法进行蛋白定量后,进行凝胶电泳(80 V转120 V)分离蛋白;采用湿转法转膜(250 mA,2 h)将靶蛋白转移至PVDF膜上;采用5 g·L-1脱脂牛奶,在室温下封闭2 h;参照目的抗体说明书将稀释好的目的一抗加入PVDF膜中,置于4 ℃冰箱进行孵育过夜。次日,用TBST漂洗,之后加入辣根过氧化物酶标记的山羊抗兔(二抗)进行室温孵育2 h。洗膜后加入显影剂,置于暗室内曝光显影。以GAPDH或α-tubline为内参,计算目的蛋白的相对表达量。
1.8 统计学方法采用SPSS 22.0统计学软件进行统计学分析。多组间比较采用单因素方差分析,两样本间比较采用t检验。检验水准:α=0.05。
2 结果
2.1 UVB组和对照组细胞中OGG1的表达变化
2.1.1 UVB组和对照组细胞中OGG1 mRNA表达变化实时荧光定量-PCR检测结果显示:与对照组(UVB照射时间为0 min)相比,随着UVB照射时间的延长,细胞中OGG1 mRNA相对表达量先上升后下降,差异均有统计学意义(均为P<0.001);UVB照射时间为10 min时,OGG1 mRNA的相对表达量最高(图1)。
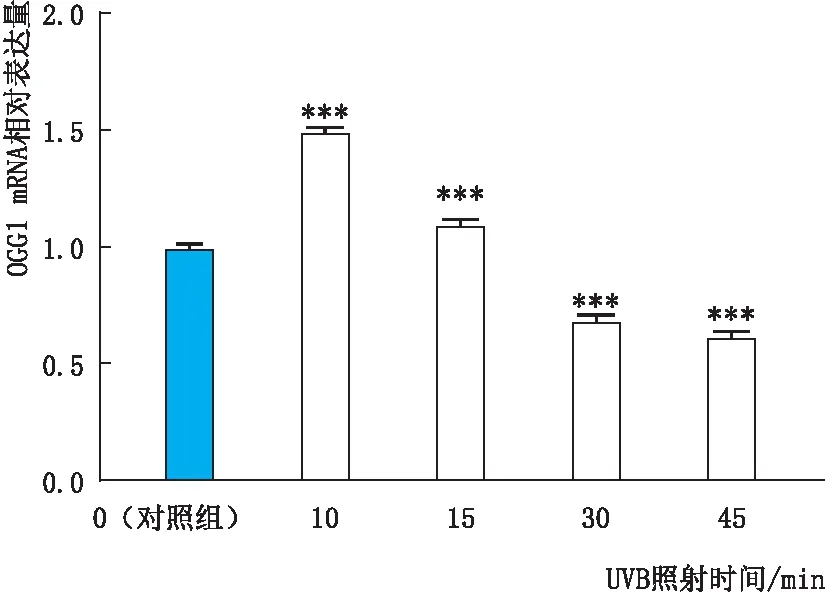
图1 UVB组和对照组细胞中OGG1 mRNA相对表达量 与对照组相比,***P<0.001。
2.1.2 UVB组和对照组细胞中OGG1蛋白表达变化免疫印迹实验检测结果显示:UVB照射10 min时,OGG1蛋白相对表达量明显较其他照射时间组升高,且与对照组(UVB照射时间为0 min)相比升高显著(均为P<0.001);随着UVB照射时间的延长,细胞内OGG1蛋白相对表达量呈时间依赖性下降(图2)。因此,本实验细胞氧化损伤模型采用UVB照射 10 min处理细胞。
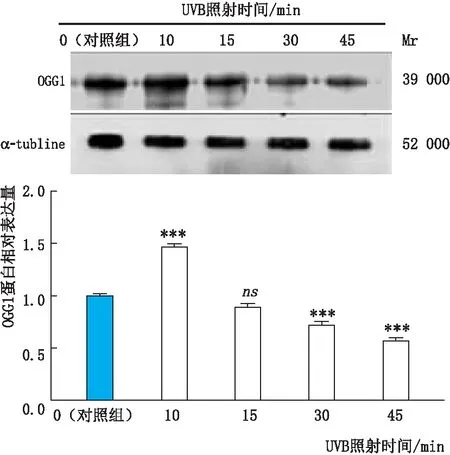
图2 UVB组和对照组细胞中OGG1 蛋白相对表达量 与对照组相比,***P<0.001,ns示差异无统计学意义。
2.2 OGG1敲降模型验证实时荧光定量-PCR检测结果显示:与siNC组相比,靶向设计的转染siOGG1组的三个亚组中细胞OGG1 mRNA相对表达量均显著降低(均为P<0.001),siOGG1#2组降低最为明显(图3)。此外,免疫印迹实验检测结果也证实:与siNC组相比,转染siOGG1#2后,OGG1蛋白水平的表达也显著降低。因此,选取siOGG1#2进行后续实验的OGG1敲降模型的制作。
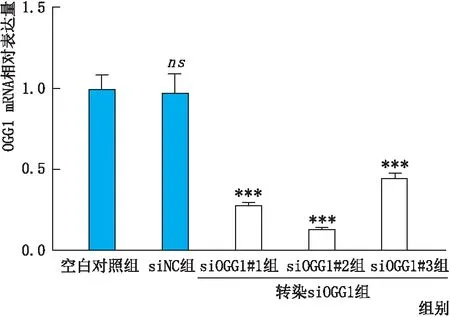
图3 转染后各组细胞OGG1 mRNA相对表达量 与空白对照组相比,ns示差异无统计学意义;与siNC组相比,***P<0.001。
2.3 转染后经UVB照射各组细胞的状态变化
2.3.1 转染后经UVB照射各组细胞DNA氧化损伤程度变化免疫荧光染色结果显示:与UVB+ siNC组相比,UVB+ siOGG1#2组15A3(染色反应底物是8-oxoG,它是OGG1的修复作用靶点)染色阳性细胞明显增多(图4)。
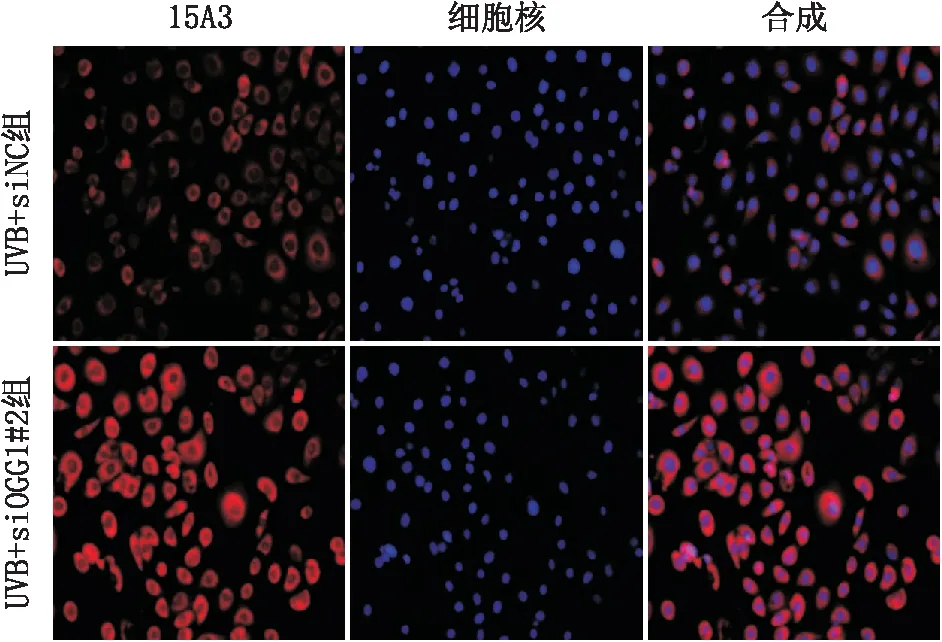
图4 免疫荧光染色结果显示经UVB照射后UVB+ siNC组和UVB+ siOGG1#2组15A3染色阳性细胞变化
2.3.2 转染后经UVB照射各组细胞活力变化CCK8实验结果显示:与UVB+siNC组和空白对照组相比,UVB+siOGG1#2组细胞活力均明显降低,差异均有统计学意义(均为P<0.01)(图5)。
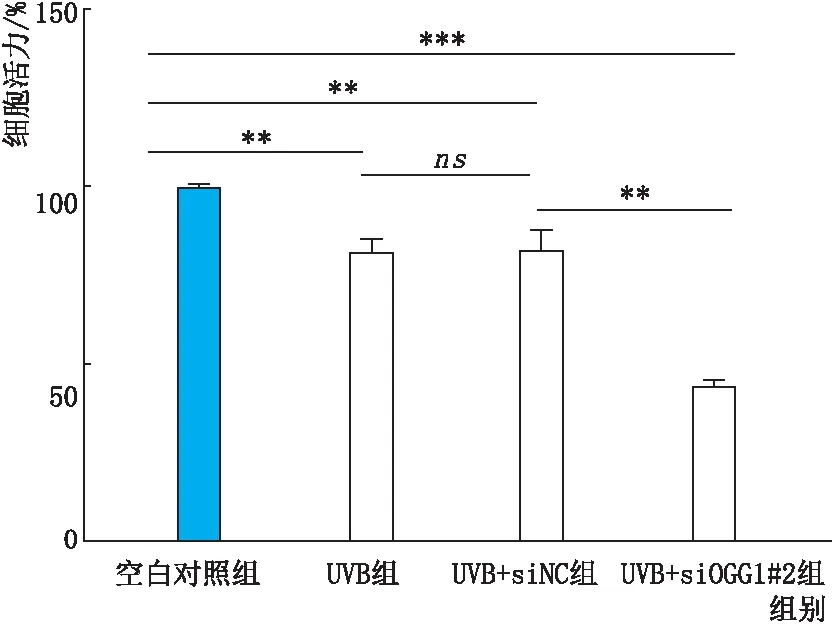
图5 转染后经UVB照射各组细胞活力变化 ns示差异无统计学意义,**P<0.01,***P<0.001。
2.3.3 转染后经UVB照射各组细胞凋亡相关蛋白BAX和BCL-2表达情况免疫印迹实验检测结果显示:与UVB+siNC组相比,转染后UVB+siOGG1#2组细胞促进凋亡的蛋白BAX表达明显增加,而抑制凋亡的蛋白BCL-2表达则明显下降(均为P<0.001)(图6)。
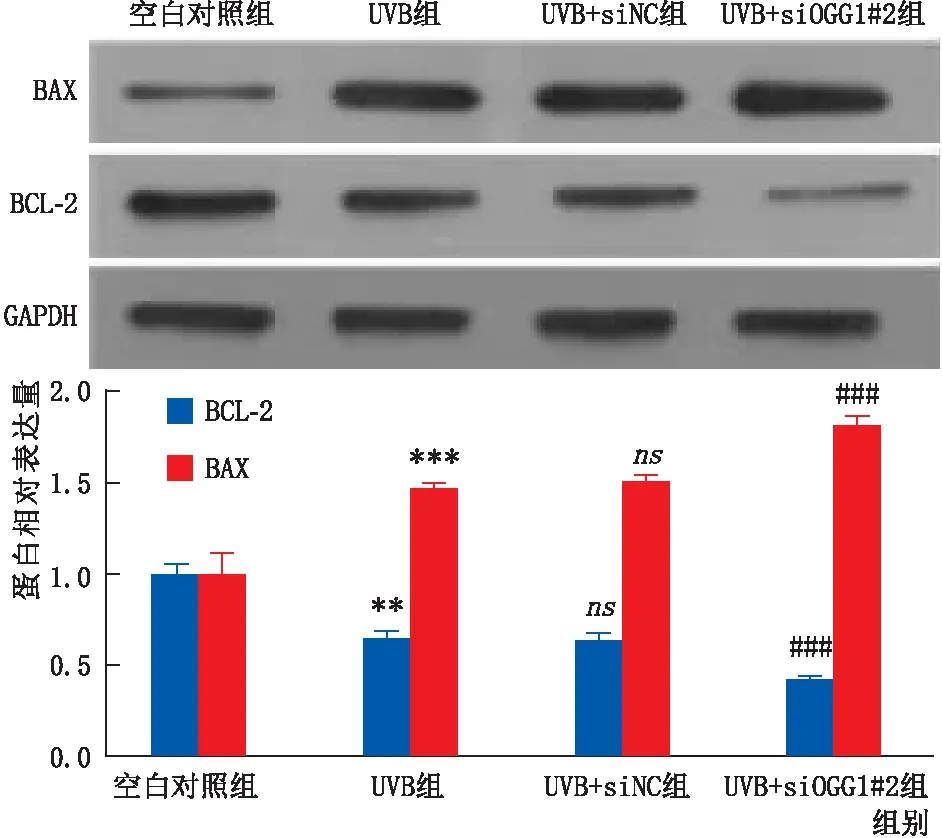
图6 转染后经UVB照射各组细胞凋亡相关蛋白BAX和BCL-2表达情况 ns示与 UVB组相比差异无统计学意义; 与空白对照组相比,**P<0.01,***P<0.001; 与UVB+ siNC组相比,###P<0.001。
3 讨论
ARC是白内障的主要类型之一,其病因目前仍不明确[8]。现阶段通过手术摘出混浊的晶状体仍是治疗ARC的主要方法。因此,探明ARC的病因及发病机制对其防治具有重要意义[8]。分子生物学研究表明,自由基和氧化应激导致的LEC内大分子物质(如DNA、RNA、脂质和蛋白质等)异常和细胞凋亡是ARC发病的病理基础[9]。研究发现,引起晶状体氧化损伤与应激的自由基来源有两类,一类是由外源性UVB等对晶状体组织长期刺激损伤所产生[10];另一类则是由内源性的线粒体氧化呼吸链损伤所致[11]。本研究利用UVB处理人LEC,模拟ARC发生的氧化微环境,建立体外人LEC 氧化损伤的细胞模型,旨在寻找能有效抑制UVB所引起的细胞内DNA损伤的药物,减少LEC的凋亡。
LEC内DNA发生氧化损伤后,机体会自发启动DNA氧化损伤修复系统进行防御。研究发现,DNA氧化损伤修复主要有5种通路,包括BER、直接修复、核苷酸切除修复、错配修复和双链断裂修复通路[12]。上述任何一个DNA修复过程受到抑制均有可能导致细胞活力下降和凋亡发生。其中,BER通路是主要的DNA损伤修复通路[12-13],该通路利用DNA糖苷酶识别并切除受损的核酸位点,并在DNA双链上形成去嘧啶位点(AP位点);然后AP核酸内切酶在AP位点的5’端或者3’端切断DNA链产生裂口;最终通过DNA聚合酶和DNA连接酶合成新的片段并填补缺口,形成DNA新链[13-14]。此通路能否有效完成,主要取决于氧化损伤修复基因能否及时有效地修复受到破坏的DNA。而OGG1是BER通路中发挥修复功能的关键酶,其修复机制主要是靶向识别和切除受损DNA双链中氧化损伤的产物8-oxoG,从而修复其损伤,起到保护细胞的作用[15]。多项研究均已证实,OGG1基因的多态性与ARC易感性相关,有助于鉴定出ARC的高风险人群[16-17]。我们前期研究[18-19]发现,在三种轻度早期ARC患者样本中,OGG1基因和蛋白表达均显著升高,并且与8-oxoG的表达呈正相关。此外,OGG1基因的表达受其CpG岛和组蛋白区表观遗传修饰的动态调节。本研究中,我们首先以时间梯度采用UVB照射来诱导人LEC以构建细胞氧化损伤模型,模拟ARC中的氧化环境。结果发现,UVB照射10 min时,细胞中OGG1 mRNA和蛋白表达水平均显著升高。敲降OGG1表达之后,LEC内DNA氧化损伤标志物8-oxoG显著增加,说明OGG1参与LEC内受损DNA的修复过程。
研究发现,OGG1基因的失活或缺失能够抑制癌细胞的增殖,从而引起癌细胞凋亡,如乳腺癌、前列腺癌、肺磷癌等[20-21]。也有研究表明,OGG1可能是阿尔茨海默病和ARC等多种衰老性和退行性疾病潜在的生物标志物[4,22-24]。近年来,许多研究探讨了OGG1在ARC患者中的表达和功能机制。在敲除OGG1基因的斑马鱼氧化损伤模型中,研究表明,OGG1功能的丧失会加速ARC的发展进程[25]。同时,OGG1在过氧化氢诱导的氧化损伤模型中,对细胞的凋亡起调控作用,而针对UVB诱导的氧化损伤模型目前尚未见相关报道。本研究在UVB诱导的细胞氧化损伤模型中初步探讨OGG1功能机制,设计靶向OGG1的siRNA,发现OGG1修复能力的减弱会明显抑制LEC的细胞活力,从而导致细胞凋亡发生。
综上所述,本研究结果证实了OGG1基因在UVB诱导的细胞氧化损伤模型中通过BER通路参与受损DNA的修复过程,调控LEC内受损DNA的修复和细胞凋亡过程。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
中国现代医生(2022年21期)2022-08-22
现代仪器与医疗(2021年6期)2022-01-18
中国药学药品知识仓库(2021年11期)2021-11-25
昆明医科大学学报(2021年1期)2021-02-07
健康护理(2021年12期)2021-01-14
中小学德育(2020年11期)2020-03-18
学校教育研究(2018年27期)2018-05-14
教育界·上旬(2016年12期)2017-05-25
山东青年(2014年10期)2014-11-24
