
星豹蛛四个羧酸酯酶基因克隆及溴氰菊酯胁迫下的表达模式
2022-05-19 06:36:02王雅丽赵萌萌张晓晨
昆虫学报 2022年4期
王雅丽, 赵 瑞, 王 美, 赵萌萌, 张晓晨, 李 锐
(山西农业大学植物保护学院, 山西太谷 030801)
昆虫体内的解毒酶系对外源有毒物质有重要的防御作用(Hemingwayetal., 2004)。羧酸酯酶(carboxylesterases, CarEs)是生物体重要的解毒酶系,广泛存在于动植物及微生物体内,能够把包含酯键的内源或外源物质分解为酸和醇(Heikinheimoetal., 1999)。羧酸酯酶具有多种生物学功能,如参与外源物质的代谢,调节生长发育,降解信息素等,是一类多功能超家族酶(Ransonetal., 2002; Kontogiannatosetal., 2011)。大部分昆虫羧酸酯酶属于B型酯酶(张建琴, 2014)。根据羧酸酯酶的催化能力和细胞定位,将其分为14个簇(Oakeshottetal., 2005),归为3个功能组,分别为与消化/解毒相关功能组(A-C簇)、与信息素/激素代谢相关功能组(D-G簇)和与神经/发育相关功能组(H-N簇)(Oakeshottetal., 2010; 张建琴, 2014)。在与神经/发育相关功能组中,乙酰胆碱酯酶具有催化能力(Johnson and Moore, 2012)。研究表明,羧酸酯酶可以降解酯类气味分子,还可以通过失活酯类信息素和寄主挥发物来调控梨小食心虫Grapholitamolesta的交配和觅食行为(Weietal., 2021)。 在家蝇Muscadomestica抗拟除虫菊酯类品系中,上调的羧酸酯酶基因参与对杀虫剂的代谢(Feng and Liu, 2020)。
星豹蛛Pardosaastrigera属蛛形纲(Arachnida)蜘蛛目(Araneae)狼蛛科(Lycosidae),是蜘蛛类捕食性天敌的优势种,星豹蛛种群数量的变化影响着害虫的发生数量(李锐等, 2014a, 2015)。化学农药广泛应用于农业生产中,在使用高浓度杀虫剂防治农林害虫的同时也严重威胁着害虫天敌的生存(张珠凤, 2001)。研究表明,高浓度除草剂胁迫后星豹蛛的活动能力显著降低(袁泽斌等, 2017)。高浓度杀虫剂胁迫后,星豹蛛捕食量显著降低,雌雄蛛交配时间延长且交配率下降(刘小荣, 2013)。目前已报道的对星豹蛛的研究主要集中在生命表的构建(燕晶晶等, 2021),化学农药对其行为(袁泽斌等, 2017)和捕食能力(李锐等, 2014b)等方面的影响,在分子研究方面关于羧酸酯酶在星豹蛛体内的作用鲜有报道。基于本课题组前期对星豹蛛的转录组测序结果,从中筛选出15个羧酸酯酶基因,主要分布在J, M和N 3个簇,还有1个基因不能很好地与其他簇聚在一起,选择其中4个基因进行克隆和序列分析,同时利用RT-qPCR技术检测其在不同组织的表达特性,分析这4个羧酸酯酶基因在溴氰菊酯不同时间及不同浓度胁迫后的表达模式,为进一步探讨羧酸酯酶基因的表达调控和对溴氰菊酯的解毒代谢机制研究提供理论基础。
1 材料与方法
1.1 供试蜘蛛
于山西省太谷区山西农业大学农学院试验基地采集星豹蛛雄性亚成蛛,待其生长发育为成蛛后作为溴氰菊酯胁迫试验材料。本实验室饲养星豹蛛幼蛛,待其发育为成蛛作为基因克隆及不同组织表达所需试验材料。星豹蛛单头置于含有湿棉球的指形管中(内直径1.5 cm,高8 cm),在人工气候箱(温度29±1℃,相对湿度75%±5%,光周期16L∶8D)中饲养。以黑腹果蝇Drosophilamelanogaster和黄粉虫Tenebriomolitor为主要食物,每2 d喂1次,定期更换指形管。
1.2 主要试剂
RNA提取所用的柱式动物组织总RNA抽提纯化试剂盒和胶回收所用的SanPrep柱式 DNA胶回收试剂盒购自生工生物工程(上海)股份有限公司;基因扩增所使用的TransStart®FastPfuDNA Polymerase和基因克隆所使用的pEASY®-Blunt Zero Cloning Kit购自北京全式金生物技术有限公司;荧光定量所使用的Talent qPCR PreMix (SYBR Green)购自天根生化(北京)科技有限公司;反转录试剂HiScript®Ⅱ 1st Strand cDNA Synthesis Kit购自南京诺唯赞生物科技股份有限公司;25 g/L溴氰菊酯(deltamethrin)乳油购自拜耳作物科学(中国)有限公司。
1.3 基因克隆和生物信息学分析
从本实验室建立的星豹蛛转录组数据库(未发表)中筛选羧酸酯酶基因序列,以此为模板序列设计引物(表1),引物由生工生物工程(上海)股份有限公司合成。选择星豹蛛雌成蛛3头提取总RNA,检测RNA纯度和浓度合格后,使用反转录试剂盒合成cDNA作为PCR扩增模板。PCR反应体系(50 μL): cDNA 2 μL, 上下游引物 (10 μmol/L)各1 μL, 5×TransStart®FastPfuBuffer 10 μL, dNTPs(2.5 mmol/L) 4 μL,TransStart®FastPfuDNA Polymerase 1 μL, Nuclease-free Water 31 μL。PCR反应条件: 95℃ 1 min; 95℃ 20 s, x℃ 20 s, 72℃ 1 min, 40个循环; 72℃ 5 min。琼脂糖凝胶电泳检测PCR产物,将目的条带进行割胶回收,回收产物连接转化,进行菌落PCR验证,送生工生物工程(上海)股份有限公司测序。
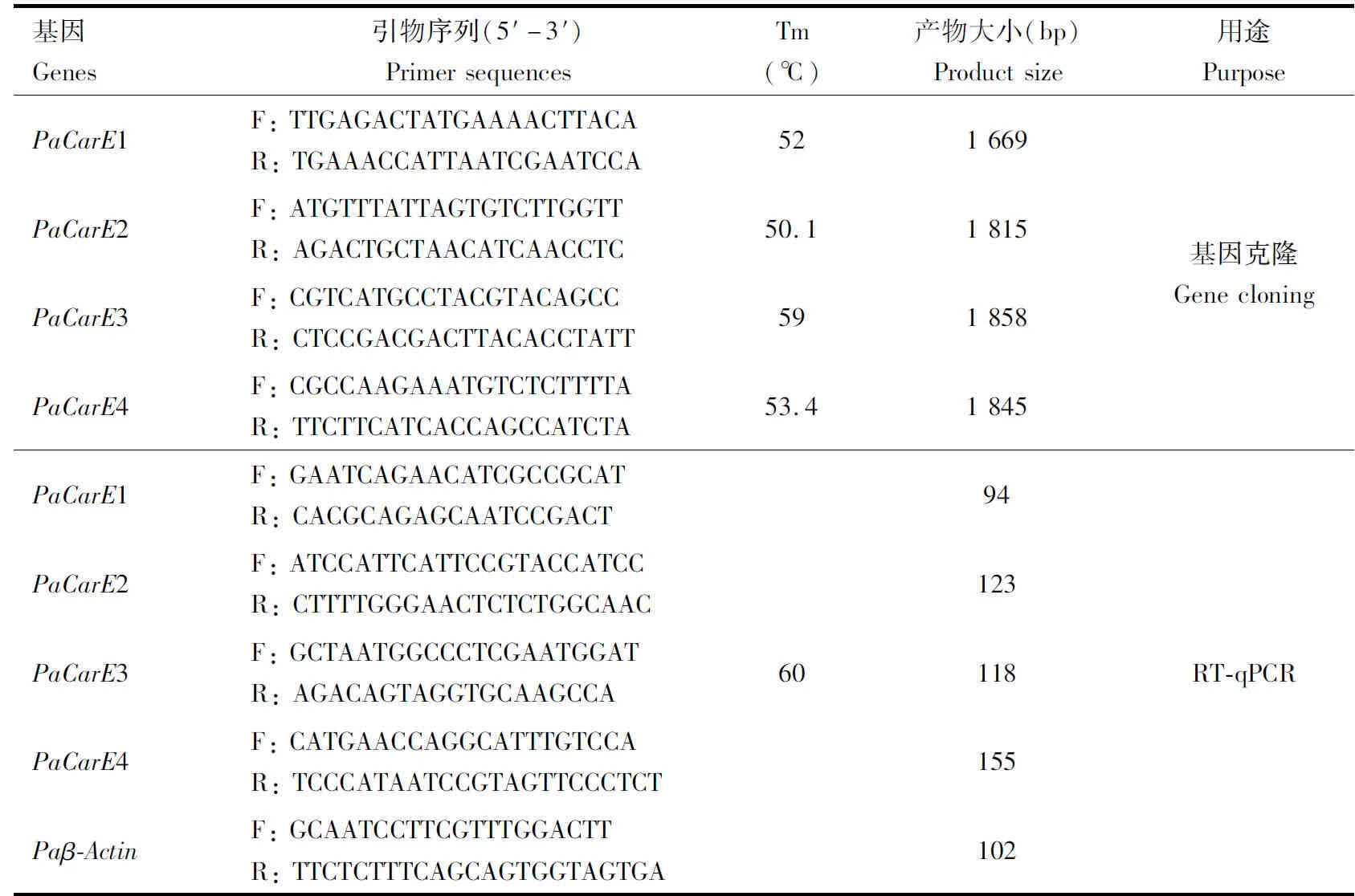
表1 引物信息Table 1 Primer information
使用在线软件ExPasy-ProtParam Tool预测蛋白的基本理化性质,包括每个羧酸酯酶基因的分子量和等电点等;使用在线软件SignalP5.0预测蛋白信号肽;使用在线软件NetNGlyc 1.0 Server预测N-糖基化位点;在NCBI保守域数据库(Conserved Domains Database, CDD)中分析羧酸酯酶基因编码氨基酸的保守序列;使用DNAMAN软件完成多序列比对,使用MEGA 7.0 软件以邻接法构建系统发育树,重复1 000次。
1.4 羧酸酯酶基因在星豹蛛成蛛不同组织中的表达量测定
选取雌雄成蛛各6头,冰上迅速解剖其头胸部、腹部和足,将相同组织混在一起提取总RNA并反转录合成cDNA(同1.3节),每处理重复3次。以星豹蛛β-Actin(李锐等, 2015)作为内参基因,羧酸酯酶基因RT-qPCR引物(表1)由生工生物工程(上海)股份有限公司合成。 RT-qPCR反应体系(20 μL): 2×Talent qPCR PreMix 10 μL, 上下游引物(10 μmol/L)各0.6 μL, cDNA 1 μL, 50×ROX Reference Dye 0.4 μL, ddH2O 7.4 μL。RT-qPCR反应程序: 95℃ 3 min; 95℃ 5 s, 60℃15 s, 40个循环; 95℃ 15 s, 55℃ 30 s, 95℃ 30 s。每样品测定3次技术重复。
1.5 溴氰菊酯胁迫后羧酸酯酶基因的表达量测定
参照李萨丽(2017)对星豹蛛成蛛的生物测定结果,选择LC10(5.151 mg/L),LC30(8.619 mg/L)和LC50(12.311 mg/L)浓度溴氰菊酯,参考池仕运等(2009)采用药膜法处理星豹蛛成蛛。用丙酮将溴氰菊酯稀释为以上3个浓度,进行浓度效应试验,对照组使用丙酮处理,每个浓度处理13~23头,12 h后收集存活的星豹蛛。选择LC30浓度(8.619 mg/L)的溴氰菊酯进行农药时间效应试验,对照组使用丙酮处理,对照组处理约70头,试验组处理约100头,分别于处理后2, 4, 8, 12, 24和48 h收集存活的星豹蛛。每个处理取样3头,重复3次,提取总RNA并反转录合成cDNA(同1.3节),进行RT-qPCR,反应体系和反应程序同1.4节。
1.6 数据分析
基因相对表达量采用2-ΔΔCt法(Pfaffl, 2001)计算。使用WPS 2019进行数据计算,使用SPSS 25.0软件中的Duncan氏新复极差法和独立样本T检验进行差异显著性分析。
2 结果
2.1 羧酸酯酶基因克隆及序列分析
克隆获得星豹蛛4个羧酸酯酶基因PaCarE1-4(GenBank登录号分别为MZ643212, MZ643214, MZ643215和 MZ643216) 的全长cDNA序列,开放阅读框(ORF)分别长1 653, 1 803, 1 827和1 818 bp,分别编码550, 600, 608和605个氨基酸,编码蛋白预测分子量分别为62.29, 67.77, 68.39和68.14 kD。PaCarE1-3等电点(pI)偏酸性,分别为5.37, 5.75和5.52,与其他已报道昆虫CarEs的等电点(Mackertetal., 2008)一致,只有PaCarE4的等电点偏碱性,为8.39。对N-糖基化位点及信号肽位置进行预测,只有PaCarE4没有信号肽。4个PaCarEs氨基酸保守基序分析结果显示,氨基酸序列均包含羧酸酯酶的保守结构域,丝氨酸活性中心F[GR]Gx(4)[LIVM]x[LIV]-xGxS[STAG]G和二硫键形成位点[EDA][DG]CL[YTF][LIVT][DNS][LIV][LIVFYW]x[PQR](“[ ]”代表不同物种在该位点为其中任意一个氨基酸);对于RF位点,PaCarE4的苯丙氨酸F变为色氨酸W;对于DQ位点,PaCarE1变为DM;PaCarE2和PaCarE4催化三联体(S-E/D-H)基序发生氨基酸的替换,PaCarE1和PaCarE3具有保守的催化三联体(图1)。

图1 推导的星豹蛛4个羧酸酯酶氨基酸序列比对Fig.1 Alignment of deduced amino acid sequences of four PaCarEs of Pardosa astrigera下划线为羧酸酯酶保守结构域,氨基酸序列保守结构用长方形和三角形标出。Conserved domains of carboxylesterase are underlined, and the conserved structure of amino acid sequence is marked by rectangle and triangle.
系统发育树(图2)及氨基酸序列比对结果表明,PaCarE1与PaCarE3被划分在与神经/发育相关功能组J簇,PaCarE1与拟环纹豹蛛Pardosapseudoannulataacetylcholinesterase type 2(GenBank登录号: AHB20143.1)的氨基酸序列一致性为71.87%,星豹蛛PaCarE3与拟环纹豹蛛AChE1 variant 1(GenBank登录号: ANQ45781.1)的氨基酸序列一致性为99.18%;PaCarE4被划分在与神经/发育相关功能组N簇,与隆头蛛StegodyphusmimosarumNeurotactin(GenBank登录号: KFM76584.1)的氨基酸序列一致性为52.22%;PaCarE2单独为一簇,与斑络新妇蛛Nephilapilipesvenom carboxylesterase-6(GenBank登录号: GFT66653.1)的氨基酸序列一致性为45.67%。

图2 邻接法构建的基于氨基酸序列的星豹蛛4个PaCarEs与其他昆虫种CarEs的系统发育树(1 000次重复)Fig.2 Phylogenetic tree of four PaCarEs from Pardosa astrigera and CarEs from other insect species usingneighbour-joining method based on amino acid sequence (1 000 replicates)F: 非鳞翅目保幼激素酯酶Nonlepidopteran juvenile hormone esterases; J: 乙酰胆碱酯酶Acetylcholinesterases; M: 胶质接触蛋白Gliotactins; N: 神经趋化蛋白Neurotactins (Oakeshott et al., 2010; 张建琴, 2014).
2.2 羧酸酯酶基因在星豹蛛成蛛不同组织的表达模式
PaCarE1-4在星豹蛛成蛛不同组织中的表达量差异显著(P<0.05)(图3)。PaCarE1在腹部中表达量最高,在头胸部及足中的表达量最低;对于同种组织中的表达量,PaCarE1在雄蛛腹部中的表达量显著高于在雌蛛中的(P<0.05),在其余组织中的表达量在雌雄间无显著差异(P>0.05)。PaCarE2在腹部中表达量最高,在头胸部及在足中的表达量较低;对于同种组织中的表达量,PaCarE2在雄蛛腹部中的表达量显著高于在雌蛛中的(P<0.05),在雌蛛头胸部及足中的表达量显著高于在雄蛛中的(P<0.05)。PaCarE3在头胸部中表达量最高,在足中次之,在腹部中最低;对于同种组织中的表达量,PaCarE3在雌蛛头胸部中的表达量显著高于在雄蛛中的(P<0.05),在其余组织中的表达量在雌雄间无显著差异(P>0.05)。PaCarE4在头胸部中表达量最高,在足中较低,在腹部中最低;对于同种组织中的表达量,PaCarE4在雄蛛头胸部中的表达量显著高于雌蛛中的(P<0.05),在其余组织中的表达量在雌雄间无显著差异(P>0.05)。
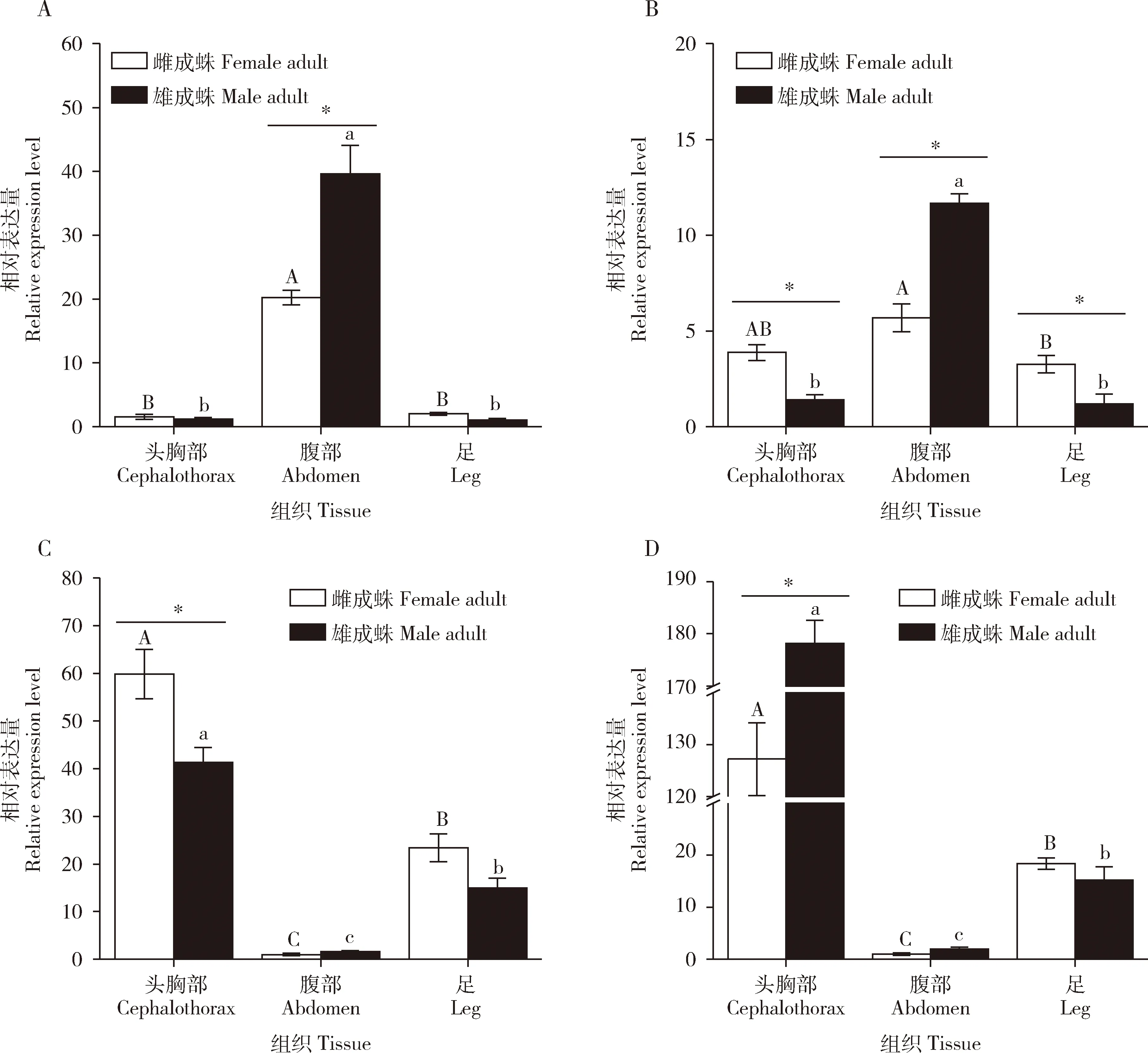
图3 星豹蛛成蛛不同组织中4个羧酸酯酶基因的表达模式Fig.3 Expression profiles of four carboxylesterase genes in different tissues of Pardosa astrigera adultsA: PaCarE1; B: PaCarE2;C: PaCarE3; D: PaCarE4. 图中数据为平均值±标准误;柱上不同大小写字母分别表示雌性和雄性成蛛不同组织间表达量差异显著(P<0.05, Duncan氏新复极差法),星号表示同一组织不同性别间表达量差异显著(P<0.05, T检验)。Data in the figure are mean±SE.Different uppercase and lowercase letters above bars indicate significant difference in the gene expression level among different tissues of female and male adults, respectively (P<0.05, Duncan’s multiple range test), while the asterisk indicates significant difference in the gene expression level in the same tissue between different genders (P<0.05, T-test).
2.3 不同浓度溴氰菊酯对星豹蛛羧酸酯酶基因表达水平的影响
与对照组相比,LC10, LC30和LC50浓度溴氰菊酯处理星豹蛛雄成蛛12 h后,PaCarE3和PaCarE4的相对表达量无显著差异(P>0.05);LC10浓度溴氰菊酯胁迫12 h后,PaCarE2的相对表达量显著升高(P<0.05),为对照的1.26倍;LC30浓度溴氰菊酯胁迫12 h后,PaCarE1和PaCarE2的相对表达量极显著升高(P<0.01),分别为对照的2.76和2.07倍;LC50浓度溴氰菊酯胁迫12 h后PaCarE2的相对表达量极显著降低(P<0.01)(图4)。
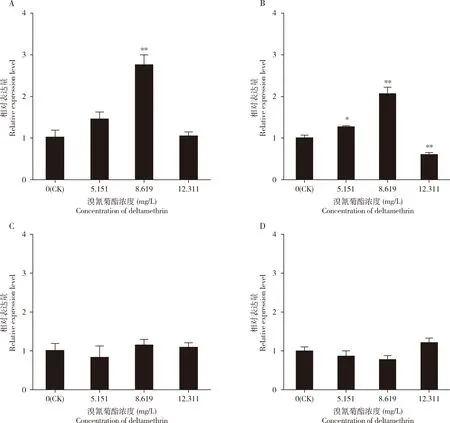
图4 不同浓度溴氰菊酯胁迫12 h后星豹蛛雄成蛛中羧酸酯酶基因的相对表达量Fig.4 Relative expression levels of carboxylesterase genes in male adults of Pardosa astrigeraafter exposed to different concentrations of deltamethrin for 12 hA: PaCarE1; B: PaCarE2;C: PaCarE3; D: PaCarE4. CK: 丙酮Acetone.参照李萨丽(2017)对星豹蛛成蛛的生物测定结果,选择LC10(5.151 mg/L), LC30(8.619 mg/L)和LC50(12.311 mg/L)浓度溴氰菊酯进行测定。图中数据为平均值±标准误;柱上星号表示与对照比较差异显著(*P<0.05; **P<0.01)(T检验)。LC10(5.151 mg/L), LC30(8.619 mg/L) and LC50(12.311 mg/L) of deltamethrin were used according to the bioassay results against P. astrigera adults in Li (2017).Data in the figure are mean±SE.Asterisk above bars indicate significant difference compared with the control (*P<0.05; **P<0.01)(T-test).
2.4 溴氰菊酯胁迫不同时间后对星豹蛛羧酸酯酶基因表达水平的影响
LC30浓度溴氰菊酯胁迫星豹蛛雄成蛛2, 4, 8, 12, 24和48 h后,PaCarE4表达量与对照组均无显著差异(P>0.05),PaCarE1-3的表达量短期内被诱导表达(图5)。溴氰菊酯胁迫2和8 h时,PaCarE1被诱导表达,表达量分别是对照的1.45和1.57倍(P<0.05);12 h时,PaCarE1和PaCarE2的表达量分别是对照的2.76和2.07倍(P<0.05);24 h时,PaCarE1的表达被显著抑制(P<0.05),PaCarE3的表达被显著诱导(P<0.05);48 h时,PaCarE3的表达被显著抑制(P<0.05)。

图5 LC30浓度(8.619 mg/L)溴氰菊酯胁迫不同时间后星豹蛛雄成蛛中羧酸酯酶基因的相对表达量Fig.5 Relative expression levels of carboxylesterase genes in male adults of Pardosa astrigera after exposedto LC30 (8.619 mg/L) of deltamethrin for different timeA: PaCarE1; B: PaCarE2; C: PaCarE3; D: PaCarE4. 图中数据为平均值±标准误;柱上不同字母表示差异显著(P<0.05, Duncan氏新复极差法)。Data in the figure are mean±SE.Different letters above bars indicate significant difference (P<0.05, Duncan’s multiple range test).
3 讨论
本研究基于星豹蛛转录组数据库克隆得到4个羧酸酯酶基因PaCarE1,PaCarE2,PaCarE3和PaCarE4,其氨基酸序列均包含羧酸酯酶的保守结构域,丝氨酸活性中心F[GR]Gx(4)[LIVM]x[LIV]-xGxS[STAG]G和二硫键形成位点[EDA][DG]CL[YTF][LIVT][DNS][LIV][LIVFYW]x[PQR](“[ ]”代表不同物种在该位点为其中任意一个氨基酸),包含RF,N端保守的半胱氨酸(C),组成氧化洞的氨基酸(GG),DQ,亲核臂(GxSxG)及催化三联体(S-E/D-H)(图1),其作用是维持蛋白高级结构的完整性和蛋白催化活性的稳定性(Cygleretal., 1993),其中PaCarE2和PaCarE4基因推导的氨基酸序列中催化三联体存在氨基酸的替换,催化三联体的改变意味着它们可能会获得一些新的功能(Yuetal., 2009),这两个基因催化活性是否改变需要进一步探究。张建琴等(2014a)研究表明,飞蝗Locustamigratoria羧酸酯酶基因LmCesF1编码序列中催化三联体发生氨基酸的替换,将该基因干扰后,飞蝗对西维因的敏感性显著提高,表明其可能参与对西维因的解毒代谢。PaCarE2在不同组织的表达模式与PaCarE1的相似,PaCarE1属于J簇乙酰胆碱酯酶,能水解昆虫中枢神经系统突触上的神经递质乙酰胆碱(Toutant, 1989),PaCarE2与venom carboxylesterase-6有较高的相似性,关于venom carboxylesterase-6的研究较少,该酶属于羧酸酯酶的一类,在红光雄蜂Bombusignitus中通过降解血液甘油三酯从而参与毒液的分解(Dengetal., 2021)。PaCarE2在星豹蛛体内的作用需要进一步的功能特性分析。PaCarE1-4都具有N-糖基化位点,N-糖基化可能与酶活性的空间结构或蛋白质结构稳定性有关(Wheelocketal., 2005),因此推测这4个羧酸酯酶具有稳定的蛋白结构或催化效率。
基因在生物体中组织表达的特异性与其生物学功能相关。郭艳琼等(2017)研究了莲草直胸跳甲AgasicleshygrophilaAhCesB1在不同组织中的表达情况,结果表明AhCesB1基因在除去头部及中肠的残体中高表达。葛娉婷(2012)研究表明在飞蝗脂肪体高表达的LmCarE26基因参与对溴氰菊酯的代谢。本研究分析了羧酸酯酶基因在星豹蛛成蛛不同组织中的表达情况,结果表明4个羧酸酯酶基因在被测组织中均存在,并且在不同组织中表达量存在差异(图3)。PaCarE1和PaCarE2在雌雄成蛛腹部中表达量最高,PaCarE3和PaCarE4在雌雄成蛛头胸部中表达量最高;徐西霞(2019)研究表明,拟环纹豹蛛羧酸酯酶基因Ppace1在脑中高表达,Ppace2在肠道中高表达,Ppace5在神经组织(脑、触肢和足)中高表达,与本研究中羧酸酯酶基因在多个组织中分布的结果一致。
利用昆虫代谢酶的可诱导性,可以推测其是否参与对化学杀虫剂的代谢过程(Poupardinetal., 2008)。研究表明朱砂叶螨Tetranychuscinnabarinus体内4个羧酸酯酶基因可以被甲氰菊酯诱导,对能够被诱导的4个基因进行RNAi,发现沉默其中任何一个均能提高朱砂叶螨对甲氰菊酯的敏感性,说明这4个基因与朱砂叶螨对甲氰菊酯的代谢密切相关(Weietal., 2019)。为探讨星豹蛛羧酸酯酶基因是否参与星豹蛛对溴氰菊酯的代谢过程,本研究分析了溴氰菊酯胁迫后星豹蛛羧酸酯酶基因的表达情况,结果表明: LC10, LC30和LC503个浓度的溴氰菊酯胁迫12 h后,PaCarE1和PaCarE2在LC30浓度溴氰菊酯胁迫下被显著诱导;LC50浓度溴氰菊酯胁迫下,PaCarE2表达被抑制,PaCarE3-4的表达与对照相比并无显著变化(图4),这可能由于羧酸酯酶对溴氰菊酯的降解存在着浓度效应,具体的原因需要进一步探究;LC30浓度溴氰菊酯胁迫不同时间后,PaCarE1和PaCarE2在短时间胁迫后能被诱导表达,溴氰菊酯的胁迫对PaCarE4表达量没有影响(图5),这与张建琴等(2014b)研究飞蝗羧酸酯酶基因在溴氰菊酯不同浓度及不同时间处理后,不同基因对溴氰菊酯响应不同结果一致。
星豹蛛头胸部的神经系统较多,并且头胸部中分布着消化道的前肠和中肠,主要由咽、吸吮胃和盲管组成,能把液态食物吸入体内(张征田等, 2010; 龚忱和王智, 2011);腹部分布后肠和马氏管等组织,昆虫肠道能分泌解毒酶对外源物质进行降解,马氏管是排泄器官,参与对外源物质的代谢(Yangetal., 2007),因此推测不同的羧酸酯酶在不同的组织中均可能发挥代谢作用。综合羧酸酯酶基因在星豹蛛不同组织及溴氰菊酯胁迫后的表达情况,推测PaCarE1,PaCarE2和PaCarE3可能参与星豹蛛对溴氰菊酯的代谢。化学农药处理浓度的大小及处理时间的长短都会对基因诱导造成一定影响,所以要阐明一个基因在化学农药代谢中的作用,我们需要利用RNA干扰技术沉默单个基因进行间接验证或将基因体外表达同时给予代谢底物直接证明基因功能。
本研究对星豹蛛4个羧酸酯酶基因进行克隆,并测定其在星豹蛛成蛛不同组织及在溴氰菊酯胁迫后的表达情况,推测其在星豹蛛体内可能发挥的作用,为进一步羧酸酯酶基因功能验证提供了基础资料。
猜你喜欢
云南化工(2021年10期)2021-12-21 07:33:28
原子与分子物理学报(2021年1期)2021-03-29 07:28:26
建材发展导向(2021年24期)2021-02-12 02:00:02
沈阳大学学报(自然科学版)(2020年1期)2020-02-18 10:12:54
淡水渔业(2019年1期)2019-01-22 06:25:36
天然产物研究与开发(2018年8期)2018-09-10 05:48:24
天然产物研究与开发(2018年4期)2018-05-07 06:47:45
中国塑料(2014年1期)2014-10-17 02:46:34
西华大学学报(自然科学版)(2014年3期)2014-09-04 00:43:38
建筑材料学报(2014年2期)2014-03-11 17:07:33
