
不同种源地喜马拉雅紫茉莉根际土壤理化性质及细菌群落结构组成分析
2022-05-14 02:52张二豪赵润东禄亚洲尹秀蔡皓兰小中
山东农业科学 2022年4期
张二豪,赵润东,禄亚洲,尹秀,蔡皓,兰小中
(西藏农牧学院食品科学学院,西藏 林芝 860000)
喜马拉雅紫茉莉(Mirabilis himalaica)又名山紫茉莉,属于多年生草本植物,主要分布在青藏高原[1];以根入药,具有治疗肾炎、肾结石、腰疼、关节痛和子宫癌的功效,是西藏一级濒危藏药材之一[2,3]。
根际土壤作为植物根系及根系分泌物的接触区域,微生物与植物生长发育、抗逆境、生态适应性及系统进化关系密切[4]。药用植物品质不仅与品种、生长环境等因素有关,而且与根际微生物密切相关。根际微生物可直接作用于植物,通过影响植物生长发育及次生代谢,进而影响药用植物生物量和药用成分含量[5-7]。有研究表明根际益生菌能够提高贯叶连翘药用成分的含量[6];枯草芽孢杆菌、梨状芽孢杆菌和金黄色固氮菌能提高党参和黄花蒿的生物量,促进其药用成分的积累[8,9]。因此,研究药用植物根际土壤细菌群落结构组成,对分离筛选与药材品质相关微生物及人工种植具有重要意义。
喜马拉雅紫茉莉作为西藏传统藏药材,相关研究主要集中在其化学成分及药理学分析[10-13],而关于根际土壤微生物群落组成的研究尚未见报道。本试验采用高通量测序技术分析西藏5个种源地喜马拉雅紫茉莉根际土壤细菌群落结构组成差异,揭示其根际细菌的核心菌群结构,并分析环境因子与细菌群落结构组成的相关性,以期为根际微生物与喜马拉雅紫茉莉药材品质间的关系研究和人工种植提供理论依据。
1 材料与方法
1.1 供试材料
2年生喜马拉雅紫茉莉根际土壤样品于2019年9月分别在林芝市巴宜区、林芝市工布江达县、山南市扎囊县、林芝市朗县和山南市桑日县采集(表1)。田间根际土壤样品采集采用抖根法[14],在每个种源地随机选取20株长势良好的喜马拉雅紫茉莉并采集根际土壤,混合均匀,每个种源地3组重复。将采集的根际土壤立即装入无菌袋中,低温运输至实验室,-80℃保存备用。
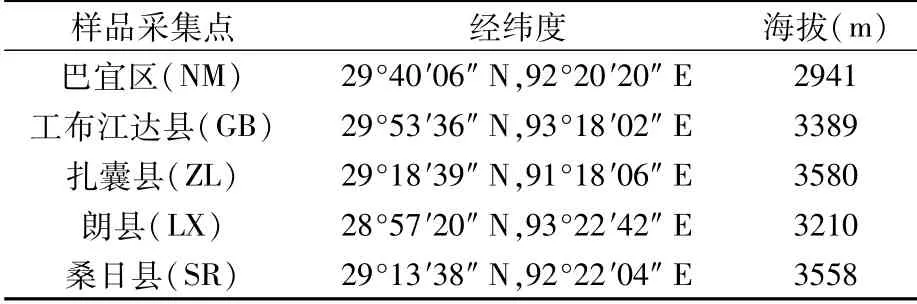
表1 样品采集点信息
1.2 试验方法
1.2.1 不同种源地喜马拉雅紫茉莉根际土壤样品理化性质分析 土壤理化性质测定采用常规检测方法[15],测定指标主要包括pH值、总磷(TP)、总钾(TK)、总氮(TN)、速效氮(AN)、速效磷(AP)、有机质含量(SOM)和电导率(EC)。土壤pH值检测采用电位法,总磷和速效磷检测采用钼锑抗比色法,总钾和速效钾检测采用火焰光度计法,总氮检测采用凯氏定氮法,速效氮检测采用碱解扩散法,电导率测定采用电极法,有机质测定采用重铬酸钾水合热法。
1.2.2 不同种源地喜马拉雅紫茉莉根际土壤样品总DNA提取 利用Mag-Bind Soil DNA Kit(Omega)提取15个根际土壤样品总DNA,琼脂糖凝胶电泳检测DNA质量并用NanoDrop ND-2000紫外分光光度计检测DNA的浓度和纯度。
1.2.3 PCR扩增 利用338F(5′-ACTCCTAC GGGAGGCAGCA-3′)和806R(5′-GGACTACHV GGGT WTCTAAT-3′)引物扩增细菌16S rRNA V3-V4可变区,引物由上海美吉生物医药科技有限公司合成。反应体系(20μL):10×PCR Buffer 2.0μL,rTaq酶(5.0 U/μL)0.20μL,正反向引物各1μL,dNTPs(5 mmol/L)1μL,DNA模板1μL,ddH2O 13.80μL。PCR反应程序:95℃5 min;95℃30 s,55℃30 s,72℃45 s,共35个循环;72℃10 min,4℃保存。琼脂糖凝胶电泳检测PCR反应结果并使用胶回收试剂盒回收PCR产物,送上海美吉生物医药科技有限公司进行测序。
1.3 数据处理与统计
利用QIIME(Version 1.9.1)软件对高通量测序原始数据进行过滤除杂,以除去低质量序列、错配序列、引物序列和过长(≥500 bp)或过短序列(≤200 bp),从而获得高质量的Clean data,根据Reads间的重叠关系将Reads拼接成Tags。在97%相似度下,利用软件USEARCH(v7.0.1090)对拼接好的Tags进行OTU聚类[16];利用Uparse(v7.0.1090)和Usearch(v7.0)软件对OTU序列进行聚类和统计;利用Mothur(v1.30.2)软件计算Alpha多样性,包括Chao1指数、Shannon指数、Simpson指数和Shannoneven指数[17];利用Qiime(v1.9.1)软件生成各分类学水平丰富度和Beta多样性[18]。
土壤理化性质测定数据利用Microsoft Excel 2019进行统计处理,使用SPSS 20.0软件进行方差检验分析。
2 结果与分析
2.1 不同种源地喜马拉雅紫茉莉土壤理化性质
由表2可知,不同种源地土壤样品中ZL的pH值为8.437,呈碱性,NM的pH值为6.620,呈酸性,其它3个种源地土壤样品呈中性;LX样品中总磷、总钾和总氮含量最高,ZL的含量最少;土壤电导率以GB最大,其次是LX,NM最小;NM速效磷含量最高,GB含量最低;有机质含量以LX最高,NM含量最低。以上结果表明,不同种源地喜马拉雅紫茉莉土壤理化性质差异较大。
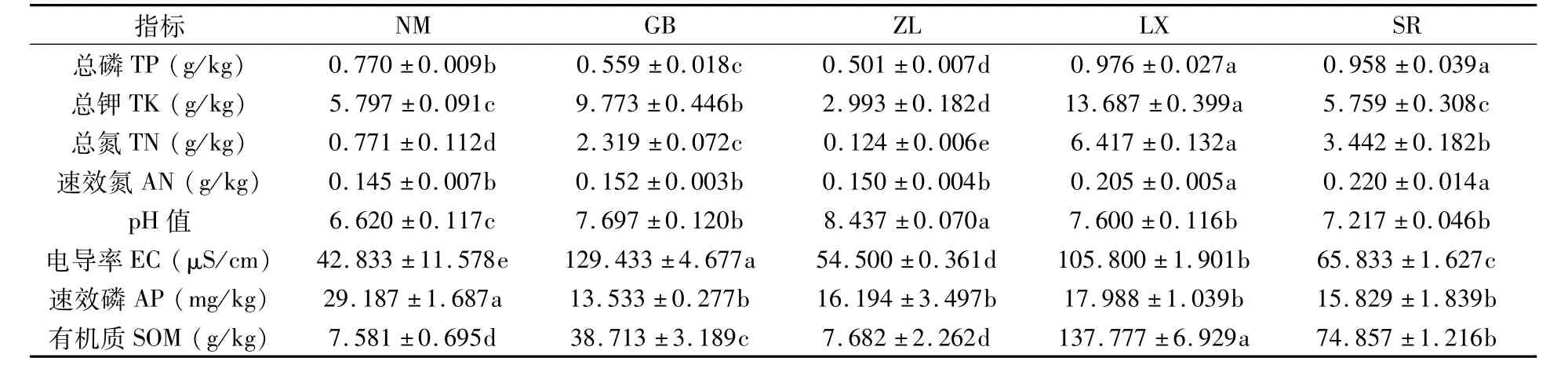
表2 不同种源地喜马拉雅紫茉莉根际土壤理化性质
2.2 不同种源地根际土壤细菌高通量测序结果及多样性分析
高通量测序结果表明,5个种源地15个根际土壤样品中共获得237120条有效序列,隶属36门85纲221目384科708属;在97%相似度水平下,对各样品进行OTU聚类,15个样品共产生2435个OTUs,NM、GB、ZL、LX和SR分别产生473、487、531、415和529个OTUs。Alpha多样性分析结果表明,不同种源地根际土壤细菌微生物多样性不同。SR和GB样品中细菌群落丰度较高,NM样品丰度指数最低;GB样品细菌群落均匀度和多样性指数最高,ZL样品最低。覆盖度代表每个样本的测序深度,5个种源地细菌群落测序覆盖度均大于97%,说明测序数据能够基本反映不同种源地细菌群落结构组成。
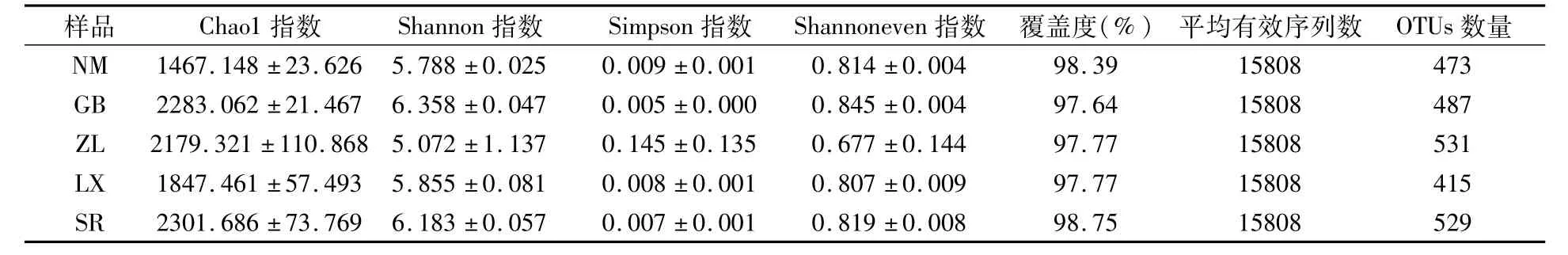
表3 不同种源地喜马拉雅紫茉莉根际土壤细菌群落Alpha多样性
2.3 不同种源地根际土壤细菌在门、属分类水平上的群落组成
2435 个OTUs隶属36门708属,相对丰度较高的细菌有8个门,NM、GB、ZL、LX和SR样品中的优势门均为放线菌门(Actinobacteria)和变形菌门(Proteobacteria),其相对丰度分别为29.97%~57.89%和17.54%~40.39%(图1A),不同根际土壤样品细菌优势门相对丰度差异较大。在属分类水平下(图1B),相对丰度≤1.00%或未被聚类的细菌群落相对丰度占32.02%~41.10%,在相对丰度≥1.00%条件下,NM、GB、ZL、LX和SR样品中分别有9、9、10、12、9个属,其优势属分别为Norank_f_norank_o_Gaiellales(5.52%)、Norank_f_norank_c_Subgroup_6(6.76%)、Methylocaldum(22.11%)、Blastococcus(5.40%)和Norank_f_norank_c_Subgroup_6(8.52%)。
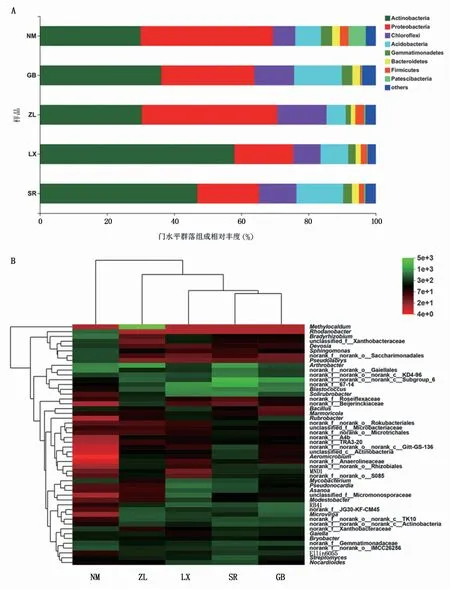
图1 不同种源地根际土壤细菌在门、属分类水平的群落组成
2.4 不同种源地根际土壤细菌核心菌群
不同种源地喜马拉雅紫茉莉根际土壤细菌在属分类水平Venn图分析结果(图2)表明,5个种源地根际土壤细菌共鉴定到708属,其中ZL样本中鉴定到的细菌属最多,LX样本中最少。各样本中共同拥有295个属,NM、GB、ZL、LX和SR样品中特有菌属分别为80、11、43、2和18。根际土壤中至少在3个种源地中共存的核心菌属有483个,占根际土壤细菌种类的68.22%。
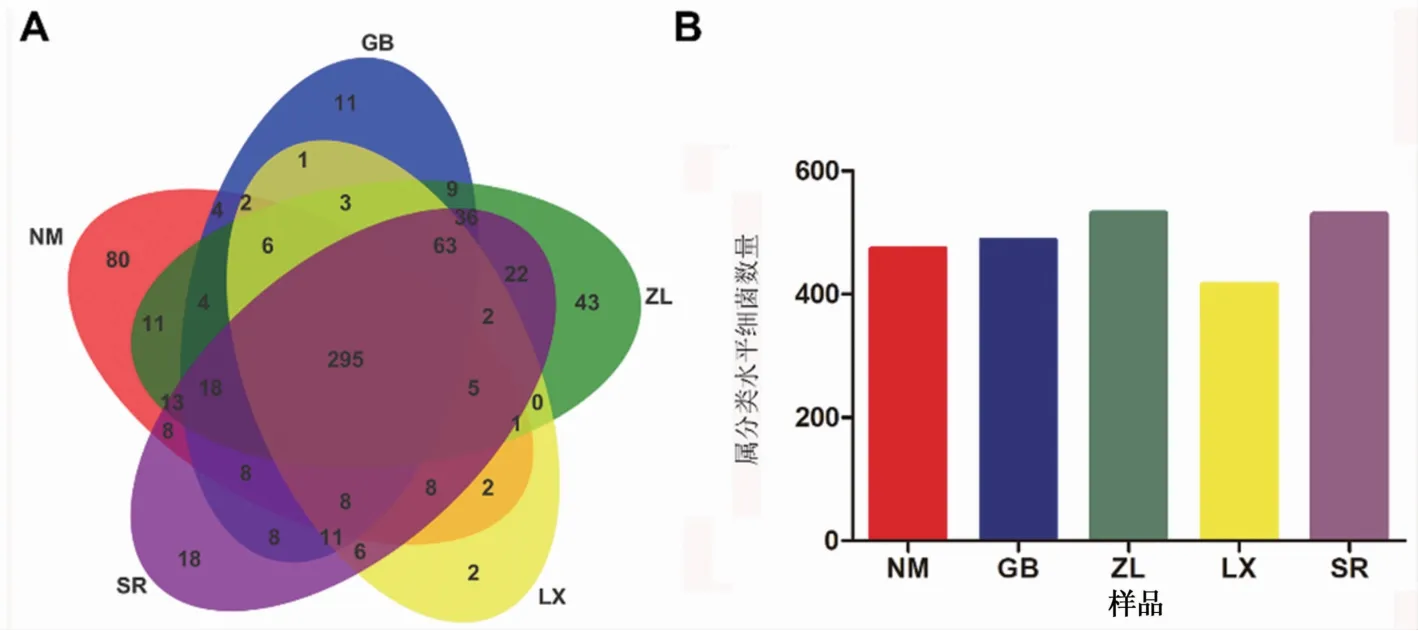
图2 不同种源地根际土壤细菌在属分类水平上Venn图和数量
2.5 不同种源地喜马拉雅紫茉莉根际土壤细菌群落比较
基于R语言对不同种源地喜马拉雅紫茉莉根际土壤细菌群落进行主坐标分析(PCoA,principal co-ordinates analysis),结果如图3所示,主坐标分析1和主坐标分析2的贡献率分别为45.77%和19.87%,累计65.64%,这是差异的主要影响因素。5个种源地根际土壤分成3大组,其中GB、SR和LX样品聚在一起,说明野生喜马拉雅紫茉莉根际土壤细菌群落结构组成相似,而人工种植的NM和ZL样品各呈一组,说明环境因子是造成根际土壤细菌群落结构差异的关键因素。
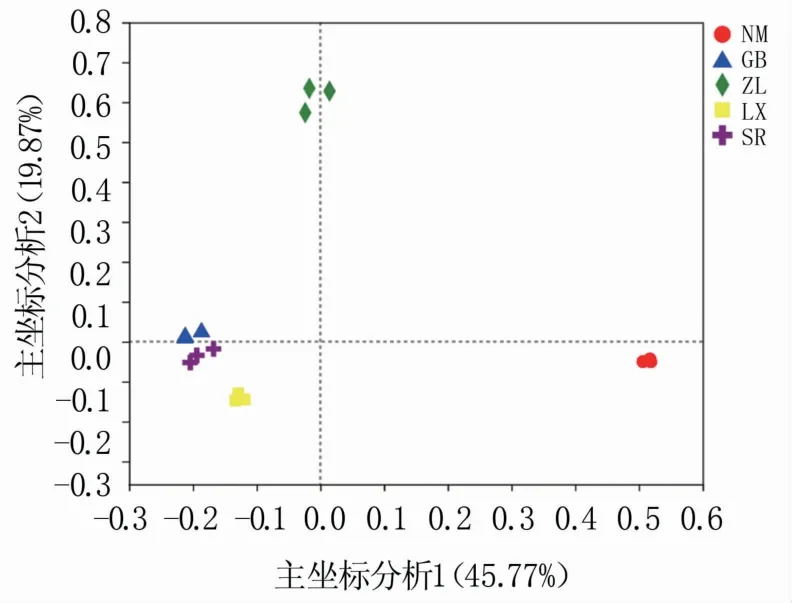
图3 细菌群落主坐标分析
2.6 土壤理化性质对不同种源地根际土壤细菌群落的影响
利用5个不同种源地喜马拉雅紫茉莉根际土壤的pH值、总磷(TP)、总钾(TK)、总氮(TN)、速效氮(AN)、速效磷(AP)、土壤电导率(EC)和有机质(SOM)等环境因子对样品OTUs聚类结果进行Spearman和RDA(Redundancy analysis)分析,结果(图4A)表明,TP、TN、AN和SOM与放线菌门(Actinobacteria)丰度显著正相关(P<0.05),TK和EC与厚壁菌门(Firmicutes)丰度显著负相关(P<0.05),pH和EC与Patescibacteria丰度显著负相关(P<0.05),TK、TN和SOM与变形菌门(Proteobacteria)丰度显著负相关(P<0.05),而土壤理化因子与酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)、浮霉菌门(Planctomycetes)和Rokubacteria丰度相关性均不显著。RDA1和RDA2的解释度分别为40.09%和32.08%,合计72.17%(图4B),说明pH、TP、TK、TN、AN、AP、EC和SOM是影响根际土壤细菌群落结构组成的关键因素。
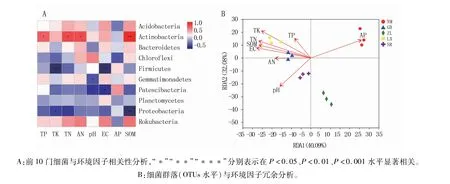
图4 环境因子对不同种源地根际土壤细菌群落结构的影响
2.7 不同种源地喜马拉雅紫茉莉根际土壤细菌群落功能预测
为分析不同种源地喜马拉雅紫茉莉根际土壤细菌群落功能,本试验采用PICRUSt软件对细菌COG功能进行预测。结果如图5所示,共获得24类COG功能,主要参与物质运输、代谢、信号转导和膜转运。与KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库比对分析发现,土壤细菌主要参与氨基酸代谢、其他次生代谢物的生物合成、碳水化合物代谢、能量代谢和辅酶因子及维生素代谢等,各样品中参与各功能信息的物种丰度差异较大(表4),其中SR样品中参与各种代谢通路的物种丰度均最高。
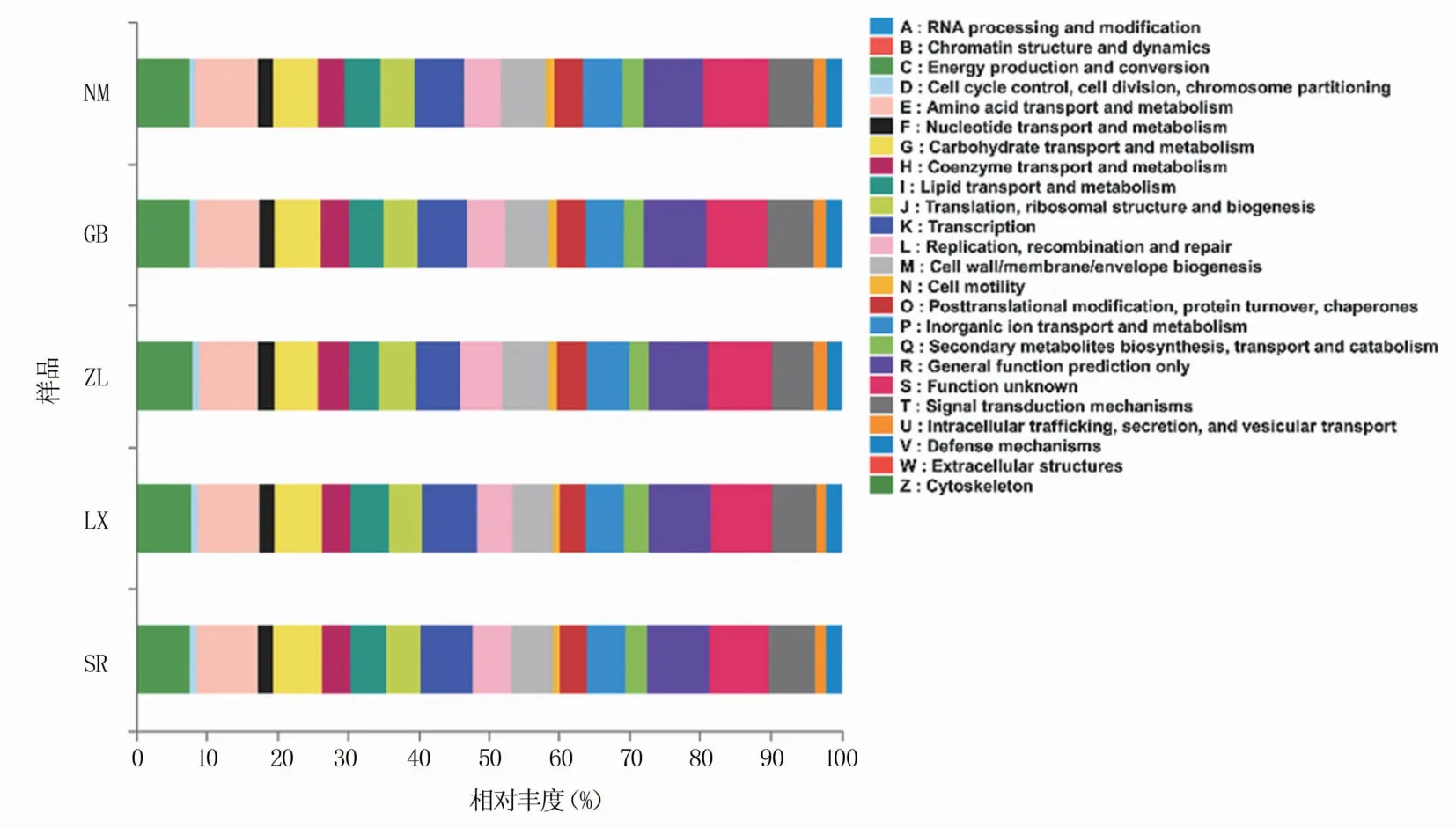
图5 不同种源地根际土壤细菌COG功能预测
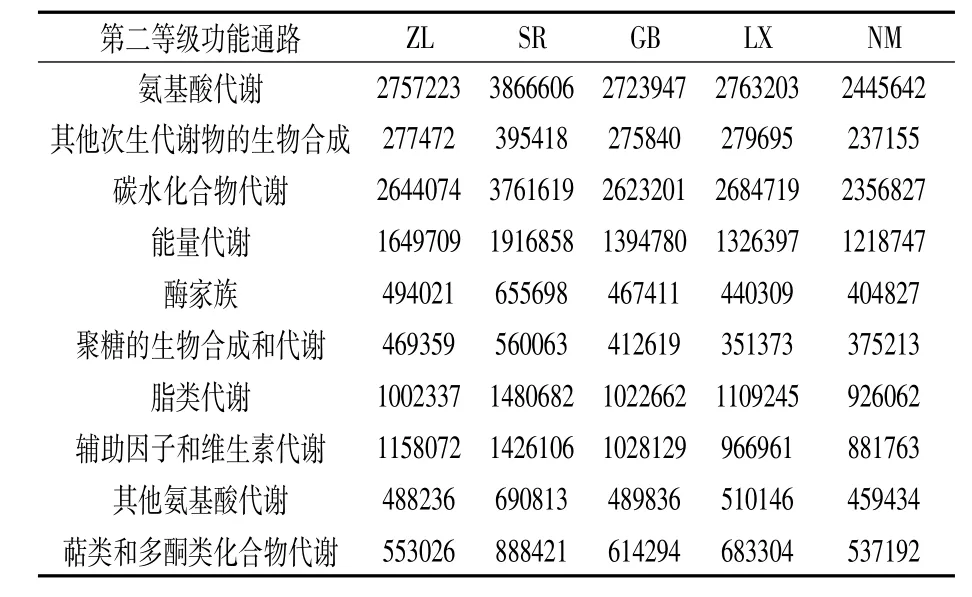
表4 不同种源地根际土壤中细菌群落的KEGG代谢通路丰度
3 讨论与结论
喜马拉雅紫茉莉作为一种传统藏药材,具有生肌、排石、利尿等功效且地上部分可作为动物饲料,有着广泛具的应用前景。根际土壤作为植物根系及根系分泌物的接触区域,其微生物对促进植物养分吸收、生长及生理过程有重要影响[19]。因此本试验通过高通量测序技术对西藏5个种源地喜马拉雅紫茉莉根际土壤细菌群落结构及多样性进行研究并对其土壤理化性质进行分析。5个种源地中GB样品细菌多样性指数最高,NM和ZL样品中最低,总体而言,野生喜马拉雅紫茉莉根际土壤细菌多样性指数均高于人工种植。有研究表明,使用化肥和农药可导致细菌多样性降低[20],NM和ZL为人工种植喜马拉雅紫茉莉,其根际细菌多样性低可能与使用化肥和农药有关。
有研究表明,在门分类水平下,土壤微生物优势类群基本相似,主要集中在10个门,如变形菌门和放线菌门等[21]。对宁夏枸杞土壤细菌群落结构的研究表明其优势类群是变形菌门、放线菌门和拟杆菌门[22];孙晓等[23]研究表明药用植物锁阳根际土壤微生物的优势类群是变形菌门和放线菌门,西洋参根区土壤细菌标志类群为变形菌门和酸杆菌门[24]。本试验结果表明,变形菌门和放线菌门是不同种源地喜马拉雅紫茉莉根际土壤细菌优势类群,同时还发现不同种源地根际土壤均存在厚壁菌门。研究表明变形菌门和厚壁菌门有助于提高药用植物的产量和质量[19]。
野生喜马拉雅紫茉莉主要分布在干热河谷地带,其生境以沙土为主。研究表明,根际微生物多样性受气候、地理位置及人类生产活动等因素影响[25,26]。Gong等[27]研究表明环境条件对红树林土壤微生物群落多样性至关重要;尚天翠等[28]在研究不同生态环境野生樱桃李土壤微生物时发现,养分显著影响土壤微生物数量。本研究发现,野生喜马拉雅紫茉莉(GB、LX和SR)根际细菌群落结构组成相似,与人工栽培喜马拉雅紫茉莉(ZL和NM)根际细菌群落结构组成差异较大,且ZL和NM样品细菌群落结构组成差异较大,这可能与生态环境(NM地区属热带湿润和半湿润气候类型,ZL地区属高原温带半干旱季风气候类型)和土壤理化性质差异有关。
本研究发现假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、鞘氨醇单胞菌属(Sphingomonas)、根瘤菌属(Bradyrhizobium)、节杆菌属(Arthrobacter)和链霉菌属(Streptomyces)是不同根际土壤的核心微生物类群。假单胞菌属和芽孢杆菌属是植物根际土壤微生物的重要组成部分,具有促进植物生长和抑制病害的作用[29];有研究表明芽孢杆菌属具有提高植物抗旱能力[30],这可能是喜马拉雅紫茉莉具有较强抗旱能力的原因。鞘氨醇单胞菌属能够利用多种有机化合物,使番茄植株在贫瘠土壤中能够较好生存[31];能够增加大豆IAA含量,促进大豆生长和抗渗透胁迫[32];同时能够抵御植物根部病害,如增强莴苣抗根腐的能力[33]。鞘氨醇单胞菌属作为不同种源地喜马拉雅紫茉莉根际土壤的核心微生物,可能有助于喜马拉雅紫茉莉在贫瘠的土壤中生长。根瘤菌属具有固定氮源和促进植物生长的作用;节杆菌属和链霉菌属具有代谢多种物质的能力,广泛分布于土壤中。节杆菌属能够增强谷子抗旱能力[34]。链霉菌属能够减轻干旱对玉米生长的影响,提高产量[35]。由此可见,不同种源地喜马拉雅紫茉莉根际土壤核心微生物类群可促进植物对营养物质的吸收、增强抗旱能力、抵御病原菌的感染,从而改善根际土壤微生物环境,促进喜马拉雅紫茉莉的生长发育。
药材品质不仅与环境因素有关,还与土壤微生物密切相关。采用PICRUSt进行基因功能预测,结果表明,不同种源地喜马拉雅紫茉莉根际土壤细菌参与代谢的物种相对丰度较高。次生代谢产物如黄酮类、醌类化合物和糖脂类化合物是喜马拉雅紫茉莉的主要药用成分[36,37],其根际土壤细菌基因中富含氨基酸、酮类、次级代谢产物、碳代谢、多糖合成和脂质代谢等与植物药用成分合成相关的通路,这些通路可能有助于其药用成分的合成、积累和运输。因此研究喜马拉雅紫茉莉根际土壤细菌群落结构组成,对获取参与药用成分合成相关基因信息、提高药用植物品质具有重要意义。
本试验表明不同种源地喜马拉雅紫茉莉根际土壤细菌群落结构存在一定的共性和差异,核心微生物类群可增强植物对营养物质的吸收,提高抗旱能力和抵御植物病原菌感染,同时还发现了参与植物药用成分合成相关的基因信息,为挖掘根际土壤微生物对药用植物抗逆境、抗病害及促进药用成分合成、积累等方面的功能及了解根际微生物与喜马拉雅紫茉莉药材品质间的关系提供了理论依据。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
中国农学通报(2022年14期)2022-06-01
江西农业学报(2022年1期)2022-03-16
土壤学报(2022年1期)2022-03-08
中国农学通报(2022年2期)2022-03-07
中国畜牧杂志(2022年1期)2022-01-20
科普童话·百科探秘(2020年9期)2020-09-06
湖北农业科学(2019年22期)2019-12-23
好日子(2019年4期)2019-05-11
艺术评论(2013年12期)2013-11-10
