
基于耳石形态的大泷六线鱼(Hexagrammos otakii)群体判别研究
2022-04-29 00:29王英俊吴莹莹宋爱环唐学玺刘洪军
海洋科学 2022年4期
孔 晓, 王英俊, 吴莹莹, 刘 童, 宋爱环, 唐学玺, 刘洪军, 4
基于耳石形态的大泷六线鱼()群体判别研究
孔 晓1, 王英俊2, 3, 吴莹莹2, 3, 刘 童2, 3, 宋爱环2, 3, 唐学玺1, 刘洪军2, 3, 4
(1. 中国海洋大学 海洋生命学院, 山东 青岛 266003; 2. 山东省海洋科学研究院, 山东 青岛 266104; 3. 青岛市海洋生物种质资源发掘与利用工程实验室, 山东 青岛 266104; 4. 青岛市水产生物品质评价与利用工程研究中心,山东 青岛 266104)
为探究邻近海域大泷六线鱼()群体间的判别问题, 共采集中国黄、渤海近岸海域7个野生群体(连云港、青岛、威海、烟台、秦皇岛、大连、丹东)和1个养殖群体(威海)的大泷六线鱼共183尾, 利用传统形态测量法和傅里叶分析对不同群体耳石形态进行多元统计分析。典型判别分析结果显示, 传统形态测量法选择8个参数值用于判别分析, 傅里叶分析法选择12个系数(elliptic Fourier descriptors, EFDs)用于判别分析, 综合判别成功率为79.2%。聚类分析显示, 威海养殖群体在最外层, 与野生群体距离较远, 与判别散点图结果一致。丹东、青岛野生群体自成一支, 与其他群体距离较远, 可被认定为独立的生态群体, 推测环境因素是导致不同群体间耳石差异的主要原因。
耳石形态; 大泷六线鱼; 群体判别; 傅里叶分析
大泷六线鱼(), 别名欧式六线鱼、六线鱼等, 属鲉形目(Scorpaeniformes)、六线鱼科(Hexagrammidae), 俗称“黄鱼”, 主要分布于中国渤海、黄海、东海, 也见于朝鲜半岛、日本及俄罗斯远东诸海[1-3]。大泷六线鱼是近海冷温性恋礁鱼类, 其肉质鲜美、营养丰富, 是我国北方重要的经济鱼种, 素有“北方石斑”之称。近年来, 由于过度捕捞和海洋生态环境污染, 大泷六线鱼资源已现衰退之势, 且存在养殖及增殖放流群体种质资源来源混杂、群体追溯研究不足等一系列问题[4]。制定大泷六线鱼种质资源保护和渔业管理条例, 需要清晰地把握其种群结构。

1 材料与方法
1.1 样品采集
本研究所用大泷六线鱼样本分别采自7个野生群体(连云港、青岛、威海、烟台、秦皇岛、大连、丹东)和1个养殖群体(威海养殖群体), 鱼体样本在实验室解冻后进行生物学测量, 测量体长(精确到1 mm)、体质量(精确到0.01 g), 并摘取耳石。样本采集地点、数量和体长等信息见表1。用超纯水清洗耳石, 除去耳石外组织和黏液, 晾干后利用电子天平(分度值0.01 mg)进行称重, 放入0.001 5 L离心管编号保存。

表1 大泷六线鱼的采样资料
1.2 耳石形态学指标
本研究中左右矢耳石形态差异不明显, 因此统一选取大泷六线鱼左侧矢耳石作为研究对象, 个别左耳石缺失时使用右耳石代替。耳石形态分析中采用OLYMPUS SZ61体视显微镜对耳石进行拍照并采集图像(图1), 使用生物图像处理软件Image- proplus6.0对耳石形态学指标进行测量。实验中使用的形态学参数包括: 耳石长(otolith length)、耳石宽(otolith width)、周长(perimeter)、面积(area)、最大半径(radius max)、最小半径(radius min)、半径比(radius radio)、平均直径(diameter mean)、耳石外切矩形长(size length)、外切矩形宽(size width) 10个形态测量指标, 3个形状指数矩形趋近率(area/box)、椭圆率(aspect)、面密度(surface density)及耳石重(otolith weight)共14个参数。传统形态参数的部分测量标准示意图如图2所示。
耳石长(otolith length)、耳石宽(otolith width)分别指耳石前端到后端的长度、背部到腹部的长度。
周长(perimeter)为耳石二维轮廓形状的总长度。
面积(area)为耳石的二维投影面积。
最大半径(radius max)、最小半径(radius min)是指通过耳石几何中心的最长半径长度和最短半径长度, 半径比(radius radio)是耳石最大半径和最小半径之比。
平均直径(diameter mean)是指过耳石几何中心的直径的平均长度。
外切矩形长(size length)、外切矩形宽(size width)分别指沿耳石长轴的最大费雷特(feret)直径和最小费雷特(feret)直径。
矩形趋近率(area/box)是指耳石面积与其外切矩形面积之比。
椭圆率(aspect)是指耳石外围椭圆长径与短径之比。
面密度(surface density)代替耳石厚度, 计算方法为耳石质量除以耳石面积。
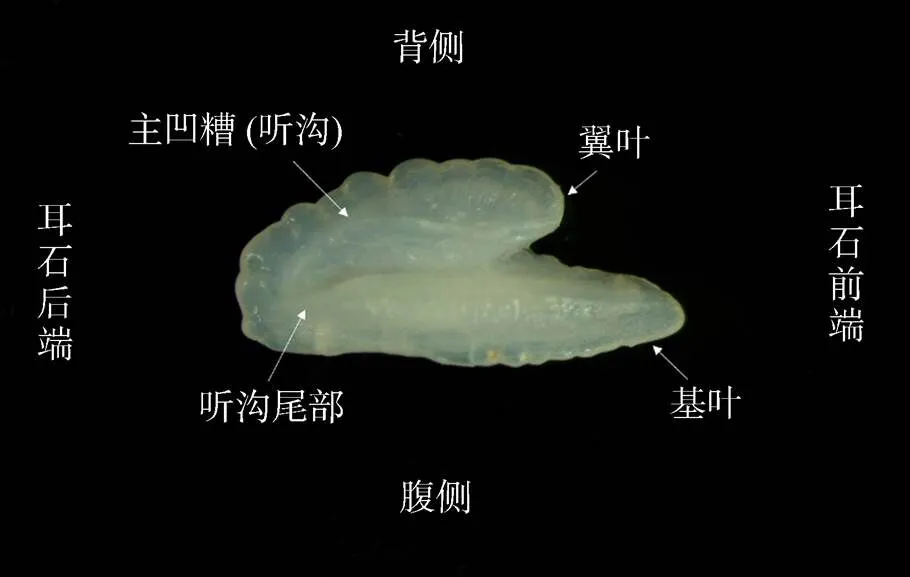
图1 大泷六线鱼左矢耳石内侧面示意图

图2 耳石测量标准的示意图
1.3 傅里叶分析
傅里叶分析对耳石的二维图像进行灰阶转换、二值化、消噪处理, 读取图像轮廓数据(图3)。通过分析耳石二维轮廓的数学表达式系数, 对耳石的轮廓曲线进行研究。使用SHAPE1.3软件包对耳石图像进行傅里叶转换, 将耳石的轮廓曲线分解成个傅里叶系数(elliptic Fourier descriptors, EFDs)[13], 得到的系数越多越接近耳石原始轮廓。一般认为, 前20个系数(1—20)可较好地描述耳石外型轮廓信息。

图3 傅里叶分析读取耳石轮廓示意图
1.4 数据分析
将形态学测量数据与提取的前20组傅里叶谐值结合进行数据分析。为提高传统形态学测量数据的有效性, 采用耳石长(otolith length)、耳石宽(otolith width)、周长(perimeter)、面积(area)、最大半径(radius max)、最小半径(radius min)、半径比(radius radio)、外切矩形长(size length)、外切矩形宽(size width)与平均直径(diameter mean)比值用于数据分析。以体长作为协变量, 对7个野生群体和1个养殖群体左矢耳石的13个形态测量参数和20个傅里叶系数进行多因素分析, 并进行正态性和方差齐性检验, 对不符合方差齐性的变量(<0.05)进行剔除, 将符合方差齐性的变量用于判别分析, 对各群体变量的平均值进行聚类分析。判别分析采用典型判别法(CDA), 聚类分析采用欧氏距离系统聚类法, 数据分析利用SPSS23.0和Excel2016软件完成。
2 结果与分析
2.1 多因素方差分析
多因素方差分析结果显示, 耳石长(otolith length)、耳石宽(otolith width)、周长(perimeter)、最大半径(radius max)、最小半径(radius min)、半径比(radius radio)、外切矩形长(size length)、外切矩形宽(size width)、矩形趋近率(area/box)、椭圆率(aspect)、面密度(surface density)、耳石重(otolith weight)显示出群体间差异(<0.05), 面积(area)在群体间未体现出差异。3、6、9、13、20在各群体间未表现出差异,2、5、8、10、15、16在各群体间差异显著(<0.05)(表2)。
2.2 判别分析
对耳石形态数据进行典型判别分析, 只有经过标准化的变量才可以用于判别分析。耳石的形态参数仅有8个指标耳石长、耳石宽、最大半径、周长、面积、外切矩形长、外切矩形宽、椭圆率被纳入判别分析, 傅里叶系数有12个指标(2、3、4、5、6、8、9、12、13、16、18、20)被纳入判别分析。总体判别成功率为79.2%, 高于单独的传统形态测量法的判别成功率65.0%和傅里叶分析的判别成功率51.4%(表3)。其中威海养殖群体判别成功率最高, 为91.7%; 秦皇岛群体判别成功率最低, 为69.2%, 连云港群体有较多的误判分类发生在烟台和威海群体之间(表3)。
从判别分析散点图中各群体的组质心可见, 威海养殖群体和其他7个野生群体区分明显, 连云港群体和威海、烟台群体之间有不同程度的混合, 与青岛、大连、丹东群体之间区分明显。青岛和大连的群体距离较近, 秦皇岛、大连、丹东群体混群程度较明显(图4)。


表3 大泷六线鱼8个群体耳石形态分析的判别结果
图4 大泷六线鱼8个群体典型判别分析散点图
Fig. 4 Scatter plot of canonical discriminant scores for 8 populations of Hexagrammos otakii
2.3 聚类分析
对7个野生群体和1个养殖群体大泷六线鱼耳石形态参数的平均值进行系统聚类分析。树状图结果显示, 秦皇岛和大连群体距离最近, 首先聚为一小类, 然后与烟台群体聚合, 形成第二小类。连云港和威海群体聚合在一起, 与第二小类聚合形成第三小类, 丹东群体、青岛群体、威海养殖群体单独形成一类(图5)。
3 讨论
3.1 耳石形态参数的确定和数据处理方法
耳石形态参数大致可以分为大小变量和轮廓变量2类。大小变量是通过线性测量软件获得耳石的表征性参数(如耳石的长、宽、周长、面积等), 耳石形状指数由上述测量参数经过数学公式转换而来或通过测量软件获取数据[14-16]。另一类轮廓变量则侧重于耳石的整体形态[17-18]。本文采用传统形态测量法和傅里叶分析相结合对不同群体大泷六线鱼进行判别, 最终传统形态测量法选取了椭圆率、外切矩形长、外切矩形宽、最大半径、耳石周长、耳石长、耳石宽、耳石面积8个测量参数用于判别分析。傅里叶分析是对于提取的轮廓系数选取了前20个(1—20)用于描述耳石外部轮廓特征[19], 并最终选取了12个系数用于判别分析。一般来说, 低谐值的傅里叶系数表征物体的一般形状, 高谐值的傅里叶系数可以表征轮廓边缘中细微的不易被捕捉的形态变化[20-21]。在本研究中, 单独使用耳石传统形态分析的判别率为65.0%, 高于单独使用傅里叶分析的判别率51.4%(表3)。以体长作为协变量对形态测量参数和20个傅里叶系数进行多重比较, 结果显示8个群体间的形态变量值存在较大差异, 而傅里叶系数3、6、9、13、20在各群体间未检测到差异, 导致单独使用傅里叶分析的判别成功率低于传统形态测量法。而使用两种方法结合进行群体判别比单独使用其中任何一种的判别成功率都要高, 说明使用形态测量法和傅里叶分析结合的方法进行群体判别的可靠性。相关研究中也选择使用两种变量结合来表征耳石形态和群体识别研究[22-24], 均取得较高的判别成功率。

图5 大泷六线鱼群体聚类结果
3.2 耳石形态差异比较与原因探讨
鱼类耳石虽体积微小但形态变化多样, 耳石边缘具齿状或不规则突起, 传统形态测量法难以解释这些细微变化。判别结果显示, 黄渤海7个野生群体和1个养殖群体大泷六线鱼耳石整体形态差异较明显, 综合判别成功率较高, 为79.2%, 高于凤鲚(70.4%)、刀鲚(68.3%)、短颌鲚(54.3%)等鲚属鱼类耳石的判别成功率[25-26], 原因在于大泷六线鱼耳石呈靴型, 基叶、翼叶区分明显, 而鲚属鱼类耳石多呈近圆形, 并且基叶、翼叶的区分度弱于大泷六线鱼, 导致大泷六线鱼耳石在形态识别上更有优势。
在威海养殖群体和野生群体的传统形态分析判别中, 有1个野生群体被误判到养殖群体, 判别成功率为97.4%。傅里叶分析未出现误判情况, 判别成功率为100%。两种方法结合, 对威海野生群体和养殖群体总的判别成功率为100%。在判别分析散点图中, 函数2可以区分养殖群体和其他野生群体。有研究表明同一种群的养殖和野生群体在不同环境下会表现出不同的耳石形状[27-28]。本研究也证实同一物种的养殖群体和野生群体的耳石形状存在差别。因此, 传统形态测量法和傅里叶分析结合的耳石形态分析方法可有效地区分养殖群体和野生群体。
综合判别结果显示, 威海养殖群体的判别成功率最高, 为91.7%。丹东、青岛、烟台野生群体的判别成功率均大于80%, 其中丹东群体判别率达到88.2%, 青岛群体为86.7%, 烟台群体为85.7%, 均较高。秦皇岛、大连、威海、连云港野生群体判别率为70%~80%, 可见丹东、青岛、烟台群体与其他野生群体混群度较低, 而其余4个野生群体间个体交流较多, 表现为多群体混栖特点。聚类分析表明威海养殖群体位于系统树的最外层, 表明养殖群体与野生群体差异较大。丹东、青岛群体自成一支, 与其他群体距离较远, 可被认定为独立的生态群体。秦皇岛和大连群体, 连云港和威海野生群体距离较近, 差异较小, 这种聚类结果与其地理分布具有一定的关系。一般来说, 群体间地理位置距离较远, 耳石的形态差异越明显。连云港和威海野生群体差异较小的原因可能是由于黄海沿岸流导致两地理群体间的个体混合。
鱼类耳石的形态与鱼类生活环境关系密切, 不同群体的耳石生长沉积过程可能会有差异。一些关于环境因素作用于鱼类耳石的研究证实, 温度、盐度、深度、摄食条件、底质类型等环境因子影响鱼体的生理活动和新陈代谢[28-29], 从而影响耳石形状[30-32]。胡发文等[33]发现, 温度、盐度等环境因子对大泷六线鱼幼鱼的呼吸及代谢等生理活动有显著影响。耳石的形成发育受到鱼体生理活动的调控, 因此环境因子在不同群体大泷六线鱼耳石形态差异方面有显著影响。较远的地理隔离是导致群体隔离的原因, 例如, 较多的误判分类发生在地理位置相近的威海和烟台群体之间, 样本重叠的原因可能是误判的群体之间有相似的环境压力, 这与其地理位置相似。因此推测耳石形状的群间变异中起主导作用的是环境因素而不是遗传差异。国内学者通过分子标记对大泷六线鱼群体进行多样性研究, 用于揭示大泷六线鱼群体间遗传结构和变异演化。李莹等[34]对6个大泷六线鱼群体线粒体控制区序列分析发现, 研究区域根据遗传距离可以被划分成山东和辽宁两个地区, 二者来源于不同的母系祖先。沈朕等[35]通过Cyt和-片段对我国近海大泷六线鱼分析显示野生群体的遗传多样性明显高于养殖群体。赵文溪等[36]通过和Cyt对黄海近海和离岸大泷六线鱼群体进行研究, 发现各群体间遗传分化不明显。可能是由于黄东海沿岸流和暖流以及大泷六线鱼增殖放流行为的影响导致不同地区基因交流机会较多, 群体间遗传差异不显著。武世雄等[37]通过微卫星分析认为不同地理群体间大泷六线鱼由于地理环境等因素导致交流减少。综上所述, 不同种属间耳石形态的差异主要由遗传因素控制, 而同一物种不同群体间耳石形态的差异主要由生活习性、生理活动和环境因子等控制, 遗传因素占次要原因。
耳石形状分析可以获得与遗传研究相补充的信息, 用于研究具有遗传同质性的多群体样本。采用多元统计分析的方法, 对中国黄、渤海近岸海域大泷六线鱼群体的耳石形态差异进行比较研究。结果表明养殖群体与野生群体差异显著, 丹东和青岛野生群体与其他群体间交流较少, 群体相对独立。研究结果为大泷六线鱼群体结构分析、种质资源保护及管理提供了技术支持和理论依据。在今后的研究中, 可结合耳石听沟形态的地标点法分析, 对耳石形态变化进行可视化图形处理[38-39], 完善耳石形态学分析方法。
[1] 中国科学院海洋研究所. 中国经济动物志(海产鱼类)[M].北京: 科学出版社, 1962: 135-137.
Institute of Oceanology, Chinese Academy of Sciences. Economic fauna of China (Marine fishes)[M]. Beijing: Science Press, 1962: 135-137.
[2] HABIB K A, JEONG D, MYOUNG J G, et al. Population genetic structure and demographic history of the fat greenling[J]. Genes & Genomics, 2011, 33(4): 413-423.
[3] CROW K D, MUNEHARA H, BERNARDI G. Sympatric speciation in a genus of marine reef fishes[J]. Molecular Ecology, 2010, 19(10): 2089-2105.
[4] 冯昭信, 韩华. 大泷六线鱼资源合理利用的研究[J]. 大连水产学院学报, 1998, 13(2): 26-30.
FENG Zhaoxin, HAN Hua. Rational utilization ofresources[J]. Journal of Dalian Fisheries University, 1998, 13(2): 26-30.
[5] 曾祥波, 张国华. 基于傅立叶耳石形态分析法对几种家鱼仔稚鱼的种类判别分析[J]. 中国水产科学, 2012, 19(6): 970-977.
ZENG Xiangbo, ZHANG Guohua. Species identification at the larval and juvenile stages for several Chinese domestic fishes by elliptical Fourier analysis of otolith form[J]. Journal of Fishery Sciences of China, 2012, 19(6): 970-977.
[6] 宋骏杰. 耳石和听沟形态分析方法及其在三种石首科鱼类群体判别中的应用[D]. 青岛: 中国科学院大学(中国科学院海洋研究所), 2018.
SONG Junjie. Otolith and sulcus morphology analyses and their applications in stock discrimination of three Sciaenids[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2018.
[7] CAMPANA S E, CASSELAM J M. Stock discrimination using otolith shape analysis[J].Canadian Journal of Fisheries & Aquatic Sciences, 1993, 50: 1062-1083.
[8] 孙诗萌, 安百胜, 蔡明城, 等. 大泷六线鱼不同群体的形态差异分析[J]. 水产学杂志, 2015, 28(4): 6-11.
SUN Shimeng, AN Baisheng, CAI Mingcheng, et al. Analysis of morphological variation of different populations of fat greenling[J]. Chinese Journal of Fisheries, 2015, 28(4): 6-11.
[9] 窦硕增, 于鑫, 曹亮. 鱼类矢耳石形态分析及其在群体识别中的应用实例研究[J]. 海洋与湖沼, 2012, 43(4): 702-712.
DOU Shuozeng, YU Xin, CAO Liang. Otolith shape analysis and its application on fish stock discrimination: a case study[J]. Oceanologia et Limnologia Sinica, 2012, 43(4): 702-712.
[10] WANG Y J, YE Z J, LIU Q. Use of otolith shape for the identification of trumpeter sillago () and silver sillago ()[J]. Journal of Ocean University of China, 2010, 9(3): 286-291.
[11] BURKE N, BROPHY D, KING P A. Otolith shape analysis: its application for discriminating between stocks of Irish Sea and Celtic Sea herring () in the Irish Sea[J]. Ices Journal of Marine Science, 2008, 65(9): 1670-1675.
[12] BOUREHAIL N, MORAT F, RAYMONDE L F, et al. Using otolith shape analysis to distinguish barracudasandfrom the Algerian coast[J]. Cybium: International Journal of Ichthyology, 2015, 39(4): 271-278.
[13] IWATA H, UKAI Y. SHAPE: a computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors[J]. Journal of Heredity, 2002, 93(5): 384-385.
[14] BURKE N, BROPHY D, KING P A. Shape analysis of otolith annuli in Atlantic herring (): a new method for tracking fish populations[J]. Fisheries Research, 2007, 91(2): 133-143.
[15] PAULO D N, ROSANGELA L, BORKO S, et al. The use of sagittal otoliths in discriminating stocks of common dolphinfish () off northeastern Brazil using multishape descriptors[J]. ICES Journal of Marine Science, 2008, 65(7): 1144-1152.
[16] LONGMORE C, FOGARTY K, NEAT F, et al. A comparison of otolith microchemistry and otolith shape analysis for the study of spatial variation in a deep-sea teleost,[J]. Environmental Biology of Fishes, 2010, 89(3/4): 591-605.
[17] 李辉华, 郭弘艺, 唐文乔, 等. 两种耳石分析法在鲚属种间和种群间识别效果的比较研究[J]. 淡水渔业, 2013, 43(1): 14-18.
LI Huihua, GUO Hongyi, TANG Wenqiao, et al. Comparative study of two otolith shape analysis for genusspecies and stocks identification[J]. Freshwater Fisheries, 2013, 43(1): 14-18.
[18] 潘晓哲, 高天翔. 基于耳石形态的鱚属鱼类鉴别[J]. 动物分类学报, 2010, 35(4): 799-805.
PAN Xiaozhe, GAO Tianxiang. Sagittal otolith shape used in the discrimination of the fishes of the genusin China[J]. Zootaxonomica Sinica, 2010, 35(4): 799-805.
[19] 王英俊. 傅里叶分析在鱼类耳石形态学中的应用研究[D]. 青岛: 中国海洋大学, 2010.
WANG Yingjun. The application of Fourier analysis in the research of otolith morphology[D]. Qingdao: Ocean University of China, 2010.
[20] 魏联, 朱国平. 基于椭圆傅里叶分析的次南极电灯鱼矢耳石形态多样性[J]. 应用生态学报, 2017, 28(9): 3078-3086.
WEI Lian, ZHU Guoping. Otolith morphological diversity ofbased on elliptical Fourier analysis[J]. Chinese Journal of Applied Ecology, 2017, 28(9): 3078-3086.
[21] 赵博. 矢耳石形态分析方法及其在石首科鱼类群体判别中应用[D]. 青岛: 中国科学院大学(中国科学院海洋研究所), 2017.
ZHAO Bo. Otolith morphological analysis and its application in stock discrimination of Sciaenidae fishes[D]. Qingdao: University of Chinese Academy of Sciences (Institute of Oceanology, CAS), 2017.
[22] GALLEY E A, WRIGHT P J, GIBB F M. Combined methods of otolith shape analysis improve identification of spawning areas of Atlantic cod[J]. Ices Journal of Marine Science, 2006, 63(9): 1710-1717.
[23] TRACEY S R, LYLE J M, DUHAMEL G. Application of elliptical Fourier analysis of otolith form as a tool for stock identification[J]. Fisheries Research, 2006, 77(2): 138-147.
[24] WANG Y J, YE Z J, LIU Q, et al. Stock discrimination of spottedtail goby () in the Yellow Sea by analysis of otolith shape[J]. Chinese Journal of Oceanology and Limnology, 2011, 29(1): 192-198.
[25] 宋超, 杨琴, 赵峰, 等. 长江口及邻近海域4个不同地理群体凤鲚矢耳石形态差异[J]. 中国水产科学, 2020, 27(10): 1125-1135.
SONG Chao, YANG Qin, ZHAO Feng, et al. Otolith Morphological variations among four geographic populations ofin the Yangtze Estuary and its adjacent waters[J]. Journal of Fishery Sciences of China, 2020, 27(10): 1125-1135.
[26] 卢明杰. 鄱阳湖水域刀鲚耳石的形态学和微化学研究[D]. 上海: 上海海洋大学, 2015.
LU Mingjie. Studies on otolith morphometry and microchemistry ofcollected from the Poyang Lake[D]. Shanghai: Shanghai Ocean University, 2015.
[27] SIMONEAU M, CASSELMAN J M, FORTIN R. Determining the effect of negative allometry (length/height relationship) on variation in otolith shape in lake trout (), using Fourier-series analysis[J]. Canadian Journal of Zoology, 2000, 78(9): 1597-1603.
[28] CARDINALE M, DOERING-ARJES P, KASTOWSKY M, et al. Effects of sex, stock, and environment on the shape of known-age Atlantic cod () otoliths[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(2): 158-167.
[29] VIGNON M T, MORAT F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish[J]. Marine Ecology Progress Series, 2010, 411(12): 231-241.
[30] FERGUSON G J, WARD T M, GILLANDERS B M. Otolith shape and elemental composition: Complementary tools for stock discrimination of mulloway () in southern Australia[J]. Fisheries Research, 2011, 110(1): 75-83.
[31] CAMPANA S E, CASSELMAN J M. Stock discrimination using otolith shape analysis[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1993, 50(5): 1062- 1083.
[32] VIGNON M. Ontogenetic trajectories of otolith shape during shift in habitat use: Interaction between otolith growth and environment[J]. Journal of Experimental Marine Biology and Ecology, 2012, 420/421(2): 26-32.
[33] 胡发文, 王晓龙, 高凤祥, 等. 温度、盐度和两种麻醉剂对大泷六线鱼幼鱼耗氧率、排氨率的影响[J]. 海洋科学, 2021, 45(1): 54-61.
HU Fawen, WANG Xiaolong, GAO Fengxiang, et al. Influence of temperature, salinity, and anesthetics on the oxygen consumption and ammonia excretion rates in fat greenling () juveniles[J]. Marine Sciences, 201, 45(1): 54-61.
[34] 李莹, 王伟, 孟凡平, 等. 利用线粒体DNA控制区部分序列分析不同地理群体大泷六线鱼遗传多样性[J]. 海洋科学, 2012, 36(8): 40-46.
LI Ying, WANG Wei, MENG Fanping, et al. Genetic diversity analysis between different stocks ofbased on analysis of mitochondrial DNA control region partial sequence[J]. Marine Sciences, 2012, 36(8): 40-46.
[35] 沈朕, 关洪斌, 郑风荣, 等. 基于和D-loop的4个大泷六线鱼群体遗传多样性分析[J]. 海洋科学进展, 2017, 35(4): 524-534.
SHEN Zhen, GUAN Hongbin, ZHENG Fengrong, et al. Analysis of genetic diversity brtween four populations ofbased on partial mtDNAand D-Loop[J]. Advances in Marine Science, 2017, 35(4): 524-534.
[36] 赵文溪, 刘莹, 于超勇, 等. 基于及基因序列的大泷六线鱼野生群体遗传多样性分析[J]. 广西科学院学报, 2019, 35(4): 308-318.
ZHAO Wenxi, LIU Ying, YU Chaoyong, et al. Analysis of genetic diversity of wild populations ofbased onandgene sequences[J]. Journal of Guangxi Academy of Sciences, 2019, 35(4): 308-318.
[37] 武世雄, 姜欣彤, 王伟, 等. 大泷六线鱼6个群体遗传多样性的微卫星分析[J]. 中国渔业质量与标准, 2018, 8(3): 52-60.
WU Shixiong, JIANG Xintong, WANG Wei, et al. Genetic diversity among 6 populations ofbased on microsatellite analysis[J]. Chinese Fishery Quality and Standards, 2018, 8(3): 52-60.
[38] 欧利国, 刘必林, 方舟. 基于椭圆傅里叶变换的鱼类矢耳石和听沟形态识别[J]. 海洋渔业, 2019, 41(4): 385-396.
OU Liguo, LIU Bilin, FANG Zhou. Identification of sagittal otolith morphology and sulcus morphology based on elliptic Fourier transform[J]. Marine Fisheries, 2019, 41(4): 385-396.
[39] GRANADOS-AMORES E, GRANADOS-AMORES J, ZAVALA-LEAL O I, et al. Geometric morphometrics in the sulcus acusticus of the sagittae otolith as tool to discriminate species of the genus(Centropomidae: Perciformes) from the southeastern Gulf of California[J]. Marine Biodiversity, 2020, 50(1): 1-7.
Stock discriminations ofbased on otolith morphology
KONG Xiao1, WANG Ying-jun2, 3, WU Ying-ying2, 3, LIU Tong2, 3, SONG Ai-huan2, 3, TANG Xue-xi1, LIU Hong-jun2, 3, 4
(1. College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China; 2. Marine Science Research Institute of Shandong Province, Qingdao 266104, China; 3. Engineering Laboratory for Exploration and Utilization of Marine Germplasm Resources of Qingdao, Qingdao 266104, China; 4. Municipal Engineering Research Center of Aquatic Biological Quality Evaluation and Application, Qingdao 266104, China)
In order to identifystocks in adjacent waters, a total of 183individuals are investigated from 7 wild populations (Lianyungang, Qingdao, Weihai, Yantai, Qinhuangdao, Dalian, Dandong) and 1 reared population (Weihai) in the coastal waters of the Yellow and Bohai Seas of China, multivariate statistical analysis of otolith morphology in different groups using traditional morphometry and Fourier analysis. Canonical discriminant analysis results show that 8 variables of traditional measurement entered in the CDA, 9 harmonics EFDs entered in CDA with Fourier analysis, and the total correct classification is 79.2%. Phylogenetic analysis shows that the Weihai cultured population cluster in the outermost layer, and has the greatest difference with the wild populations, consistent with the discriminant scatter plot results. The Qingdao and Dandong wild populations are self-contained, distance from other groups and can be regarded as independent ecological communities. It is speculated that environmental factors are the main reasons for the differences in otolith among different populations.
otolith morphology;; stock discrimination; Fourier analysis
Jun. 24, 2021
Q149
A
1000-3096(2022)04-0034-10
10.11759/hykx20210624002
2021-06-24;
2021-09-14
山东省农业良种工程项目(2019LZGC020); 山东省重点研发计划(2021LZGC027); 中海油公益基金会项目“山东省典型海岸带生物资源与生态环境保护修复研究”; 山东省农业重大应用技术创新项目(SD2019YY007)
[Agricultural Improved Seed Project of Shandong Province, No. 2019LZGC020; Key Research and Development Project of Shandong Province, No. 2021LZGC027; CNOOC Public Welfare Foundation Project; Agricultural Major Applied Technology Innovation Project of Shandong Province, No. SD2019YY007]
孔晓(1996—), 女, 山东德州人, 硕士研究生, 研究方向: 海洋生态学, E-mail: kongxiao1220@163.com; 刘洪军(1964—),通信作者, 男, 山东博兴人, 研究员, 主要从事水产增养殖、渔业资源修复与海洋生态环境调控技术研究, E-mail: HongjunL@126.com
(本文编辑: 丛培秀)
猜你喜欢
北京航空航天大学学报(2022年8期)2022-08-31
中学数学研究(广东)(2022年9期)2022-06-16
中等数学(2022年1期)2022-06-05
少儿科技(2021年12期)2021-01-20
中华养生保健(2020年7期)2020-11-16
学习周报·教与学(2019年16期)2019-10-21
老友(2018年7期)2018-08-08
北京航空航天大学学报(2018年1期)2018-04-20
养生保健指南(2017年12期)2017-12-07
科学生活(2017年9期)2017-10-16
