
红景天苷对97H细胞自噬的影响
2022-04-24 14:36:32蒋兵杨韬封龙飞王涛苏海翔
甘肃医药 2022年3期
蒋兵 杨韬 封龙飞 王涛 苏海翔,
1.甘肃中医药大学,甘肃 兰州 730000;2.甘肃省医学科学研究院/甘肃省肿瘤医院,甘肃 兰州 730050
自从Ashford和Porter[1]在1962年发现了细胞中的“自食”现象以来,自噬的研究越来越受到研究人员的关注。直到2016年,诺贝尔医学及生理学奖授予了一位日本学者Yoshinori Ohsumi,奖励他通过酵母细胞研究发现了部分自噬的机制[2],这进一步激发了人们对自噬探索的热情。研究发现,自噬其实是指细胞在受到某些刺激后,吞食损伤的细胞器,并通过膜包裹形成自噬小体,然后与溶酶体结合形成自噬溶酶体,最后通过自噬溶酶体内部的酶完成对吞噬细胞器的降解过程。自噬产生的氨基酸和脂肪酸等降解产物可以传输到细胞质中供细胞生长,残基则被排出或被滞留在细胞质中,以维持细胞内环境的稳定和细胞器的更新[3]。大量研究表明,自噬的发生与其相关自噬蛋白密切相关,其中Beclin-1基因、p62和LC3B蛋白已被证实是自噬的关键因子。Beclin-1基因主要通过与Ⅲ型PI3K形成复合体来调节其他Atg蛋白在自噬前体结构中的定位,从而起到调节自噬活性的作用[4]。p62能够结合泛素化修饰的待降解蛋白,通过与自噬膜泡分子上的LC3相互结合,将自噬降解底物招募至自噬泡中完成降解[5]。Atg7和Atg3等自噬相关分子可将LC3-I进行泛素样脂化偶联系统的加工,使LC3-I以磷脂酰乙醇胺(PtdEth,PE)共价的形式插入到自噬小体的双层膜中,使其转化为LC3-II,而LC3-I与LC3-II的转变又进一步促进了自噬小体的形成[6]。
中医药是中华民族的瑰宝,越来越多的研究表明,很多中药单体、提取物及衍生物对肿瘤细胞的自噬具有一定的影响。蒋君男等人[7]证实丹参中所含的丹参酮IIA可通过上调LC3-II/I、Beclin-1蛋白的表达,并下调p62蛋白的表达,诱导人肝癌HepG2细胞自噬。查勇等人[8]发现苦参中所含的苦参碱可通过升高WIPI1 mRNA表达水平,并上调Atg5、Beclin-1、LC3-II蛋白的表达,诱导人肝癌SMMC-7721细胞自噬。陈奇等人[9]发现紫草中所提取的紫草多糖可通过升高Atg5、Beclin-1 mRNA及LC3-II蛋白的表达诱导H22肝癌细胞自噬。
红景天又称“金根”,是景天科的一种多年生草本植物,广泛分布于严寒、干燥、缺氧、高海拔(3500~5000米)地区[10]。红景天在亚洲的运用已有悠久的历史,常被用作中草药来治疗疾病,如头痛、癔症和阿尔兹海默症等[11]。在欧洲和北美,红景天则被用作膳食补充剂来补充能量[12]。现代药理学的研究表明,红景天中的提取物主要包括苯乙醇衍生物、单萜、三萜、黄酮类化合物和酚酸[13]。而红景天苷(salidroside,Sal)是从红景天根茎中所提取得到的主要有效成分之一。大量研究表明,红景天苷具有广泛的药理学价值,如血管保护、神经保护、抗氧化、抗炎等[14,15],是一种颇具开发潜力的药物成分。越来越多的研究表明,红景天苷可影响不同类型癌细胞的增殖、迁移和凋亡。然而,红景天苷对肿瘤细胞的自噬作用及其潜在机制仍不清楚。本研究旨在探究红景天苷对高转移人肝癌细胞(MHCC97-H highly metastatic human liver cancer cells,97H)自噬的影响,为后续进一步探究其潜在机制提供理论及实验依据。
1 材料与方法
1.1 药品与试剂 红景天苷(纯度99.56%)购自美国Selleck公司。DMEM培养基购自美国Hyclone公司。胎牛血清(FBS)购自美国Gibco公司。青霉素-链霉素购自上海BasalMedia公司。MDC染色试剂盒、高效RIPA裂解液、BCA蛋白浓度检测试剂盒及SDS-PAGE凝胶制备试剂盒均购自北京索莱宝科技有限公司。AO染色试剂购自美国Chroma Technology公司。总RNA提取试剂盒和mRNA逆转录试剂盒购自北京天根生化科技有限公司。Beclin-1(1C10C4)和山羊抗兔二抗(SA00001-1)均购自美国Proteintech公司。p62(STX100685)、GAPDH(GT239)抗体购自美国GeneTex公司。LC3B(ab192890)购自英国Abcam公司。三色预染色蛋白标记物(20351ES72)购自上海YEASEM公司。
1.2 主要仪器 透射电镜(日本HITACHI公司);荧光显微镜(日本Olympus公司);多功能实时荧光定量PCR仪(美国ABI公司);电泳仪(美国Bio-Rad公司);电转仪(美国Bio-Rad公司);凝胶成像仪(美国Bio-Rad公司)。
1.3 细胞培养 人肝癌97H细胞购自上海生物科学研究所细胞中心。将97H细胞用5mL完全培养基(10%胎牛血清+1%双抗体+90%DMEM培养基)进行培养,并置于37℃、5%CO2饱和湿度培养箱中孵育。
1.4 透射电镜观察 将97H细胞用完全培养基(空白组)和80μM红景天苷处理48h,将处理后的细胞收集在10mL离心管中,用PBS清洗细胞两次。然后,用2.5%戊二醛室温固定细胞90 min,用1%四氧化锇固定细胞30min。用95%无水乙醇梯度脱水后,将细胞包埋,切片,用饱和醋酸铀酰和柠檬酸铅染色。最后,用透射电镜观察自噬囊泡、自噬小体和自噬溶酶体的形成,并进行拍照。
1.5 MDC染色和AO染色 将对数生长期的97H细胞以5×104个/mL的密度接种在无菌的盖玻片上,并置于37℃、5%CO2饱和湿度培养箱中过夜培养。次日待细胞贴壁后,以不含药物的完全培养基培育97H细胞作为空白组,以不同浓度红景天苷处理97H细胞作为实验组。处理48h后,用1Xwashingbuffer清洗细胞2次,分别用MDC和AO染色工作液室温避光染色30min,再用1 X washingbuffer清洗3次。最后,将盖玻片附着在载玻片上,并在盖玻片上滴加collection buffer。通过荧光显微镜进行观察并拍照。
1.6 qRT-PCR 用终浓度为0μM(空白组)、20μM、40μM、80μM的红景天苷分别处理97H细胞48h后,使用Trizol试剂提取总RNA,并通过逆转录二步法合成cDNA,且每组样品设置3个复孔。PCR引物由苏州金唯智生物科技有限公司设计合成。反应体系20μL,含SYBR Green PCR Master Mix 10μL,无酶水7.2μL,上游引物0.4μL,下游引物0.4μL及cDNA模板2μL。PCR扩增过程如下:95℃预变性3 min;95℃10s、退火30s、72℃30s,共40个循环。数据采用2-△△Ct方法进行分析。所用引物序列和最佳退火温度见表1。

表1 qRT-PCR引物序列和退火温度
1.7 Western blot 将对数生长期的97H细胞以1×106的密度接种于培养皿中,置于37℃、5%CO2饱和湿度培养箱中过夜培养。次日待细胞贴壁后,用浓度为0μM、20μM、40μM和80μM的红景天苷处理细胞。处理48h后,收集细胞,提取总蛋白。采用免疫印迹法检测Beclin-1、p62、LC3-I和LC3-II相关自噬蛋白的相对表达水平,并以GAPDH作为参照。
1.8 统计学分析 应用IBMSPSSStatistics22统计软件对实验结果进行统计学分析,检测数据以平均±标准差表示。组间均数比较采用单因素方差分析,P<0.05代表差异具有统计学意义。
2 结果
2.1 自噬现象观察 空白组中未发现自噬囊泡、自噬小体和自噬溶酶体,80μM红景天苷处理的细胞中发现了不同大小(300~900nm)的自噬囊泡、自噬小体和自噬溶酶体。见图1。为使自噬过程更加直观,我们根据其结果制作了相应的自噬流程图。见图2。

图1 红景天苷诱导97H细胞自噬(透射电镜,×8000)
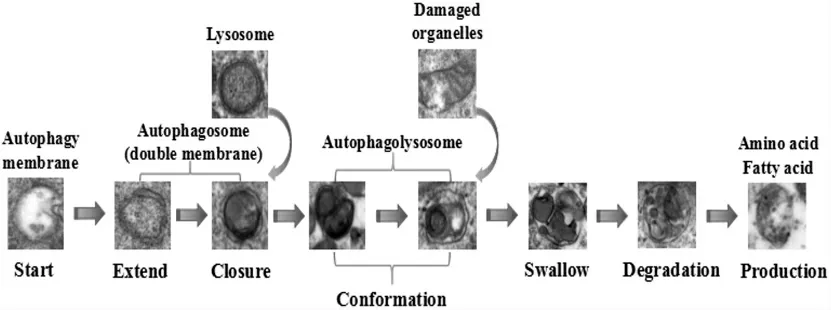
图2 红景天苷处理97H细胞后的自噬流程(透射电镜,×8000)
2.2 红景天苷诱导97H细胞自噬 与空白组相比,红景天苷处理组中与自噬小体相关的绿色荧光数量呈剂量依赖性和时间依赖性的增加,且荧光亮度也随之增强。见图3。与空白组相比,红景天苷处理组中与自噬溶酶体相关的红黄色荧光数量呈剂量依赖性和时间依赖性的增加,且亮度也随之增强。见图4。

图3 MDC染色检测红景天苷对97H细胞自噬的影响(荧光显微镜×200)

图4 AO染色检测红景天苷对97H细胞自噬的影响(荧光显微镜×200)
2.3 红景天苷对97H细胞中自噬相关基因mRNA水平的影响 qRT-PCR结果显示,与空白组相比,红景天苷处理组可上调Beclin-1的相对mRNA水平(P<0.05),同时,p62的相对mRNA水平与Beclin-1的相对mRNA水平呈负相关。见图5。进一步说明红景天苷可以诱导97H细胞的自噬。

图5 红景天苷对97H细胞中Beclin-1、p62 mRNA表达水平的影响
2.4 红景天苷对97H细胞中自噬相关蛋白表达的影响 Western blot结果显示,与空白组相比,红景天苷处理组的LC3-II与LC3-I的比例呈剂量依赖性增加(P<0.05);同时,红景天苷还能上调Beclin-1蛋白的相对表达水平,下调p62蛋白的相对表达水平(P<0.05)。见图6。

图6 红景天苷对97H细胞中LC3-I、LC3-II、Beclin-1、p62蛋白表达的影响
3 讨论
红景天不仅可被用来治疗一些疾病[11],还可以被用于膳食来补充能量[12]。红景天苷是从红景天根茎中提取得到的主要有效成分之一,越来越多的研究表明,红景天苷不仅对血管和神经起到保护作用[14,15],同时还具有抗氧化和抗炎功能[16-17]。既往研究发现,红景天苷可影响乳腺癌[18]、胃癌[19]、结肠癌[20]、膀胱癌[21]等不同类型癌细胞的增殖、迁移和凋亡,但是否有诱导这些细胞自噬的作用及其作用机制尚不清楚。本研究选用了人肝癌97H细胞作为研究对象探究红景天苷对其的自噬作用,并对机制进行了初步探讨。
自噬的形成可以概括为以下三个步骤:(1)细胞接收到自噬诱导信号后,在细胞质中产生一个类似于“脂质体”样的膜结构,该膜不断膨胀,最终形成一个由两个脂质双分子层组成的囊泡,可在透射电镜下观察到。(2)当自噬囊泡持续延伸时,细胞质中的所有成分都被细胞膜吸收,然后自噬囊泡闭合,成为一个封闭的球形自噬小体,可通过透射电镜及相应的染色方法观察到。(3)自噬小体和溶酶体结合形成自噬溶酶体,可借助透射电镜及相应的染色方法观察到。在此期间,溶酶体中所含的酶会降解自噬小体的内膜,并且两者的内容量被整合,自噬小体的含量也被降解,所形成的氨基酸、脂肪酸等降解产物被输送到细胞质中以获得细胞生长所需的能量,残基则被排出或被滞留在细胞质中,以维持细胞内环境的稳定和细胞器的更新[3]。本研究通过透射电镜观察到红景天苷可诱导97H细胞自噬的现象,通过MDC染色、AO染色检测到97H细胞中的自噬小体及自噬溶酶体在胞质中的分布。qRT-PCR和Western blot的结果表明,红景天苷诱导97H细胞自噬的机制可能与上调Beclin-1 mRNA、下调p62 mRNA、增加LC3-II与LC3-I比例、上调Beclin-1蛋白水平、下调p62蛋白水平有关。
本研究以人肝癌97H细胞作为研究对象,运用透射电镜、MDC染色、AO染色、qRT-PCR及Western blot研究了红景天苷对其的自噬作用,并对机制进行了初步探讨。研究结果为红景天苷诱导肿瘤细胞自噬的研究提供了理论和实验基础,也为后续进一步研究红景天苷通过调控自噬来发挥抗肝癌效应的相关分子机制提供了重要的实验依据。
猜你喜欢
实验室研究与探索(2021年11期)2021-01-28 22:02:14
生物化工(2021年2期)2021-01-19 21:28:13
生物化工(2020年1期)2020-02-17 17:17:58
山东化工(2020年15期)2020-02-16 01:00:12
读与写(2019年35期)2019-11-05 09:40:46
实验技术与管理(2019年6期)2019-07-12 07:38:04
现代职业教育·高职高专(2018年7期)2018-05-14 16:20:40
教育教学论坛(2017年40期)2017-10-26 11:00:54
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:43
西南军医(2015年3期)2015-04-23 07:28:32
