
The phycocyanin-chlorophyll-protein complexes isolated from Chroomonas placoidea*
2022-04-07 09:09MinCHENQinLIYuxuanWANGJingWANGKunZHANG
Min CHEN, Qin LI, Yuxuan WANG, Jing WANG, Kun ZHANG
Institute of Life Science, Yantai University, Yantai 264005, China
Abstract An active photosystem (PS) Ⅱ particle and two light-harvesting complexes, as well as their subcomplexes that have not been reported previously, were isolated from a cryptophyte Chroomonas placoidea by Triton X-100 sucrose density gradient centrifugation. The f luorescence spectra revealed that there were effi cient energy couplings between phycocyanin (PC645) and chlorophyll (Chl) within both zones Ⅲ andⅣ of the gradient, which were designated respectively as light-harvesting complex and PSⅡ particles whose size was 15-20 nm according to negative staining in electron microscopy. When the two complexes were further resolved into sub-complexes, the energy coupling was retained in the core PSⅡ complex (named as zone Ⅳ-2 of the sucrose gradient), which contained almost no outer antenna pigment Chl c. Sodium dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE) showed that the PC645 components appeared in Chl-containing protein complexes were mainly the β subunit with molecular weight of 20 kDa. These results demonstrate that PC645 in this cryptophyte was structurally but preferentially combined with the lightharvesting complex and PSⅡ core. The excitation energy absorbed by PC645 could be directly transferred to Chl a (especially the long wavelength of Chl a) in the PSⅡ reaction center or via the Chl a/ c-protein complex. The β subunit corresponded to the terminal f luorescence emission and might play an important role in transmitting energy from PC645 to the Chl-protein complex. The results will help in elucidating the architecture and function of the energy transfer system comprising phycobiliproteins and Chl-protein complexes in cryptophytes.
Keyword: Chroomonas placoidea; phycocyanin-chlorophyll-protein complex; photosystem Ⅱ; lightharvesting complex (LHC); phycocyanin (PC645); f luorescence spectra
1 INTRODUCTION
Oxygen-evolving photosynthetic organisms have evolutionarily conserved reaction centers, which generally contain chlorophylla(Chla)-protein complexes and are varied in their types of lightharvesting components, which are Chla/b-protein complex in green plants, or Chla/c-protein complex in chromophytic plants and phycobiliproteins (PBPs)in blue and red plants. Cryptophytic algae are chromophytic plants. They are thought to be a unique lineage that emerged from red algae after a secondary endosymbiosis event. Unusually, they possess two sets of co-existed light-harvesting systems in their chloroplast. Some cryptophytes use phycobiliproteins,whose maximal absorption is around 500-650 nm, as their primary light-harvesting complex (LHC).Cryptophytic PBPs are partially inherited from red algae. During the evolution, the phycobilisomes(PBS) in red algae, which are disc-shaped or half disc-shaped multimeric PBPs complexes anchored on the external surface of thylakoid membranes, became reduced and allophycocyanin disappeared. The remaining PBPs, either phycoerythrin (PE) or phycocyanin (PC), depending on the algal species,transferred to the luminal side of the thylakoids (Gantt et al., 1971; Spear-Bernstein and Miller, 1987;Ludwig and Gibbs, 1989), and has evolved into the only type of light-harvesting antenna that locatedwithin the thylakoidal lumen found up to now.Although the number of genes encoding PBP subunits diff ers in diff erent species (Kieselbach et al., 2018),the PBPs in cryptophytes are reported as consisting primarily of α, α′, and β subunits. From crystal structure analysis, PE545 and PC645 were proposed to form a quaternary structure of ( α β) ( α′ β)heterodimers (Morisset et al., 1984; Wilk et al., 1999;van der Weij-De Wit et al., 2006). The β subunit of cryptophytic PBPs, encoded by the chloroplast genome, has been shown to be inherited from red algae (Douglas and Penny, 1999), while the evolutionary origin of the nuclear-encoded α subunits remain still unknown (Douglas and Penny, 1999;Kieselbach et al., 2018; Greenwold et al., 2019;Tomazic et al., 2020).
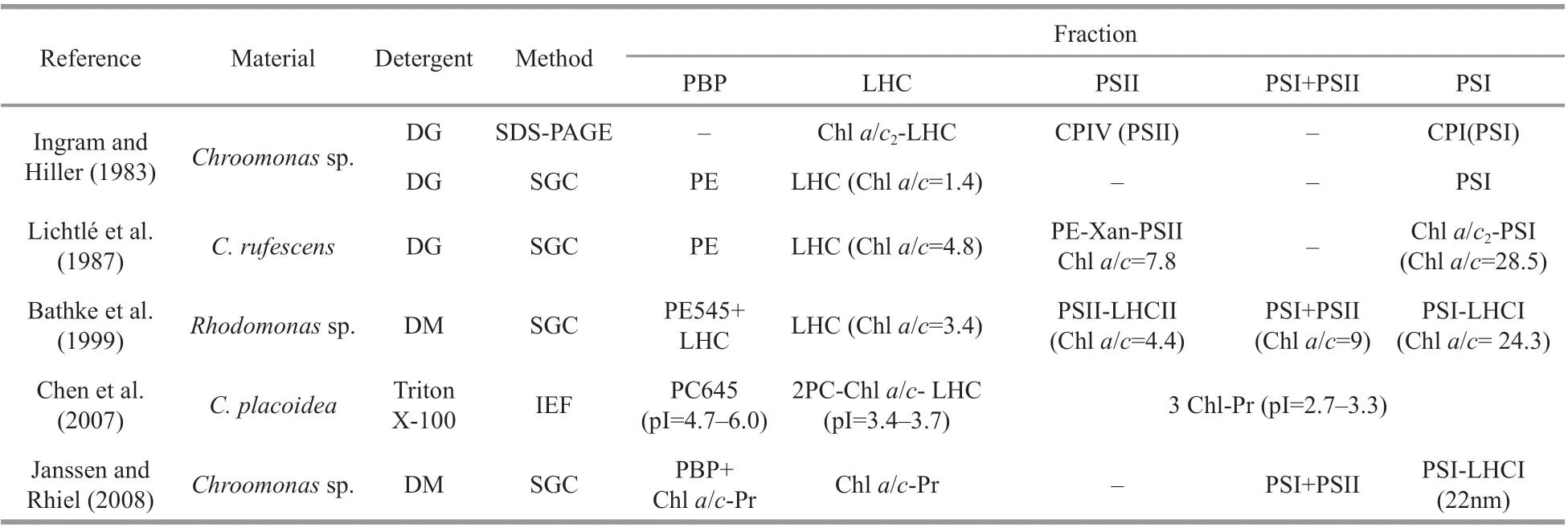
Table 1 Pigment-protein complexes isolated from cryptophytes
Furthermore, as chromophytic algae, cryptophytes contain another type of light-harvesting system in addition to soluble PBPs, namely the insoluble Chla/c-protein complex. The Chla/c- protein complex is located predominantly in the stacked regions of thylakoid membranes, and probably mediates the excitation transfer from PE to photosystem (PS) Ⅱ, as determined by steady-state and time-resolved f luorescence measurements in whole cells (Lichtlé et al., 1980, 1987; Ingram and Hiller, 1983; Cheregi et al., 2015). Previous works focused on isolated cryptophytic PBPs, including their crystal structure(Becker et al., 1998; Wilk et al., 1999; Doust et al.,2004; van der Weij-De Wit et al., 2006; Harrop et al.,2014), biological and spectroscopic characteristics(MacColl et al., 1995, 1998; Li and Chen, 2013), and energy transfer (Collini et al., 2010; Novoderezhkin et al., 2010; Arpin et al., 2015). In contrast, the Chlcontaining complexes are poorly understood.Generally, 3-6 pigment-protein complexes could be resolved from cryptophytic algae thylakoids. In addition, the results varied from each other under diff erent solubilization and separation conditions(Table 1). Most of them were Chl-protein complexes,only a LHC-PSⅡ fraction (Lichtlé et al., 1987) and a Chla/c2were found to combine with cryptophytic PBP (Chen et al., 2007). Actually, the arrangement of cryptophytic PBPs in the thylakoid lumen, as well as the excitation energy transfer pathways between PE or PC to LHC and the reaction centers, which are Chlcontaining complexes embedded in the membrane,remain uncertain. It was reported that isolated PE,such as antenna PE545, showed diff erent features of energy transfer compared with those in intact cells(van der Weij-De Wit et al., 2006; Cheregi et al.,2015). It was concluded that spectroscopic experiments on isolated components could not necessarily predict their precise function in intact cells (van der Weij-De Wit et al., 2006). In the present study, active PC-LHC proteins and PC-PSⅡcomplexes, as well as their sub-complexes were resolved from a PC645-containing cryptophyteChroomonasplacoidea, and provided a direct view of the composition in sub-complex level, energy transfer and arrangement of PBPs and Chl-protein complexes in this cryptophytic alga.
2 MATERIAL AND METHOD
2.1 Isolation and preparation of thylakoid and PC samples
Thylakoids ofChroomonasplacoideawere prepared as described previously (Chen et al., 2007)and stored at -70 ° C in 50-mmol/L Tricine-NaOH buff er (pH 8.0) containing 20% glycerol. Chlorophyll concentration and Chla/cratio of thylakoids and isolated zones were determined following the procedure of Jeff rey and Humphrey (1975). The blue supernatant containing PC was treated with 70%ammonium sulfate overnight and centrifuged at 36 000×gfor 10 min to deplete thylakoids and Chlcontaining components. The supernatant was then precipitated with 100% ammonium sulfate. The PC sediment was collected by centrifuging at 2 000×gfor 20 min, dialyzed against 0.5-mol/L phosphate buff er(pH 7.5) overnight, and stored at 4 ° C in darkness.
2.2 Sucrose gradient centrifugation of pigmentprotein complexes
Before sucrose gradient centrifugation, stored thylakoids were thawed and washed twice with 7.5-mmol/L EDTA (pH 8.0) containing 50-mmol/L sorbitol and centrifuged at 36 000×gfor 10 min. The thylakoid sediments were re-suspended in ultrapure water at 0.8-mg/mL Chl and adjusted to pH 8.0 using(CH3)4NOH. Then, Triton X-100 was added to a f inal concentration of 0.7%. The mixture was stirred at 4 ° C for 30 min, then centrifuged at 30 000×gfor 10 min. Aliquots (0.5-1.0 mL) of the supernatant were immediately loaded onto a 5-mL SW-41 centrifuge tube containing a discontinuous sucrose gradient comprising 1.5 mol/L (1.0 mL), 0.8 mol/L(0.5 mL), 0.6 mol/L (0.5 mL), 0.5 mol/L (1 mL),0.4 mol/L (1 mL), and 0.3 mol/L (0.5 mL), each containing 0.02% Triton X-100. The gradients were centrifuged in a Hitachi-55P-72 SW-41 centrifuge at 220 000×gfor 14-16 h.
2.3 Secondary sucrose gradient centrifugation of pigment-protein sub-complex
Zones containing pigments were collected, pooled and diluted in 50-mmol/L Tricine- NaOH buff er (pH 8.0), then centrifuged and concentrated at 38 000×gfor 7 h in 10-kDa ultra-f iltration tubes to deplete sucrose. The concentrates were adjusted to 0.5-mg/mL Chl with (CH3)4NOH (pH 8.0), and 20% Triton X-100 was added to a f inal concentration of 0.5%.The mixtures were incubated at 4 ° C for 10 min in darkness, then subjected to a secondary step of centrifugation in 12-mL tubes containing discontinuous sucrose gradients comprising 1.5 mol/L(2.5 mL), 1.0 mol/L (1.0 mL), 0.8 mol/L (1.0 mL),0.6 mol/L (1.0 mL), 0.5 mol/L (2.0 mL), 0.4 mol/L(1.0 mL), and 0.3 mol/L (1.0 mL) under the same centrifugal conditions described above.
2.4 Measurement of photo-oxidative activity
For assay of PSI activity, dichlorophenol indophenol (DCPIPH2) oxidation was performed on fractions collected from sucrose gradients as described by the Teaching and Research Group of Plant Physiology, Department of Biology, East China Normal University (1980). Measurement of PSⅡactivity was carried out as described by Samuelsson and Prezélin (1985).
2.5 Negative staining
Portions of 5 μL of zone Ⅳ from the sucrose gradient were applied at 0.03 mg/mL or 1.5 mg/mL to the surface of hydro-treated and carbon-coated copper grids then stained for 1 min. The grids were washed with ddH2O after removing the drops, and then dyed with 5-μL 2% uranyl acetate for 1 min. Samples were detected at 200 kV by transmission electron microscopy (Talos F200C), and images were recorded by camera (Ceta) with a magnif ication of 73 000×g.
2.6 Absorption and f luorescence spectra
Thylakoids and zones collected from sucrose gradients were diluted with 50-mmol/L Tricine-NaOH (pH 8.0). Absorption spectra were recorded on an UV-3400 spectrophotometer, and f luorescence spectra at room temperature or 77 K (-196 ° C) were recorded with a Carian Cary Eclipse f luorimeter.Glycerol (50%) was added to samples before low temperature spectrum determinations.
2.7 Polypeptide analysis by SDS-PAGE
Thylakoids, PC samples, as well as pigmentprotein complexes and sub-complexes isolated by sucrose gradient centrifugation were precipitated with 20% trichloroacetic acid in microcentrifuge tubes,and centrifuged at 25 000×gfor 10 min. The sediments(except PC samples) were washed twice with 80%acetone/alcohol to deplete chlorophylls and carotenoids. All samples were incubated in sample treating buff er (0.1-mol/L Tris-HCl buff er, pH 6.8,containing 4% SDS, 10% 2-mercaptoethanol, 20%glycerol, and 0.005% bromophenol blue) for 3 h at 37 ° C. Thylakoids were centrifuged at 30 000×gfor 10 min before SDS-PAGE (sodium dodecyl sulfatepolyacrylamide gel electrophoresis). Electrophoresis was carried out according to Laemmli (1970), and the gels were f ixed in 0.2-mol/L iminazole for 10min and stained with 0.2-mol/L ZnSO4, which enhances the f luorescent intensity of the PC subunit. Gels were observed and photographed under ultra-violet light in a UVP (Ultra-Violet Products)-Biospectrum imaging system, washed with 7% (v/v) acetic acid to deplete ZnSO4,and photographed under natural light after restaining by Coomassie brilliant blue G-25.
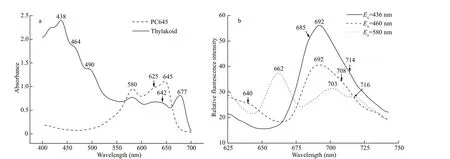
Fig.1 The absorption spectra of thylakoid and PC645 at room temperature (a) and f luorescence emission spectra at 77 K (b)

Fig.2 Triton X-100 sucrose gradient centrifugation of PC645 (a) and thylakoids (b)
3 RESULT
3.1 Thylakoids of C. placoidea
The absorption spectrum of well-washedC.placoideathylakoids showed absorption maxima corresponding to Chla(438 and 677 nm), Chlc(464 nm for the blue region; the red region absorption was covered by PC645), carotenoids (490 nm), and PC645 (580- and 645-nm typical absorption peaks and 625-nm shoulder) (Fig.1a). Low-temperature f luorescence emission spectra showed that exciting Chlaat 436 nm and Chlcat 460 nm led to PSⅡ-specif ic emission peaks with maximum at 692 nm coupled with a shoulder at 685 nm (Fig.1b). The main contributor to the two maxima is most likely the inner antenna of PSⅡ, as in higher plants and blue algae. In the emission spectrum with excitation at 580 nm, the 685-nm peak disappeared and the 692-nm peak diminished, while the maximum emission migrated to 703 nm. The 662-nm individual f luorescent emission indicated the presence of PC645, and the 716-nm shoulder represented the activity of PSI. This result was unusual, and suggested that the PC645 might energetically connect to PSI in the thylakoids ofC.placoidea, unlike the PE545 in cells ofGuillardiatheta, primarily connected to PSⅡ (Kieselbach et al.,2018).
3.2 Isolation of pigment-protein complexes
Centrifugation of Triton X-100-solubilized thylakoids (Chla/cratio was 4.04) on discontinuous sucrose density gradients gave f ive distinct fractions,named as zones I-V from top to bottom (Fig.2b).Zone I, located at the top of the tube, was orange and contained no proteins in SDS-PAGE (data not shown).It was characterized as a free pigment band.
The dark green fraction zone Ⅱ was at 0.3-0.4-mol/L sucrose gradient. It contained nearly 50% of the total chlorophyll, including Chlc(absorption at 460-465 and 628 nm) and Chla(436 and 672 nm) (Chla/cratio=2.91), as well as carotenoids (490-495 nm)(Fig.3a), and showed neither PSI nor PSⅡ oxidative activity. The excitation spectrum of zone Ⅱ showed the excitation peaks of Chla(437 and 672 nm)(Fig.3b). However, Chlccontributed less to the 700-nm f luorescence emission, compared with zones Ⅲand Ⅳ. The 464-466-nm f luorescence in the blue region, associated with Chlc, was weak (Fig.3b). The maximum emission of zone Ⅱ was at 679-680 nm when the fraction was excited at 436, 460, or 580 nm(Fig.4a). These results suggest that zone Ⅱ might be a light-harvesting Chla/c-protein complex that maintains a certain extent of energy transfer among the pigments.
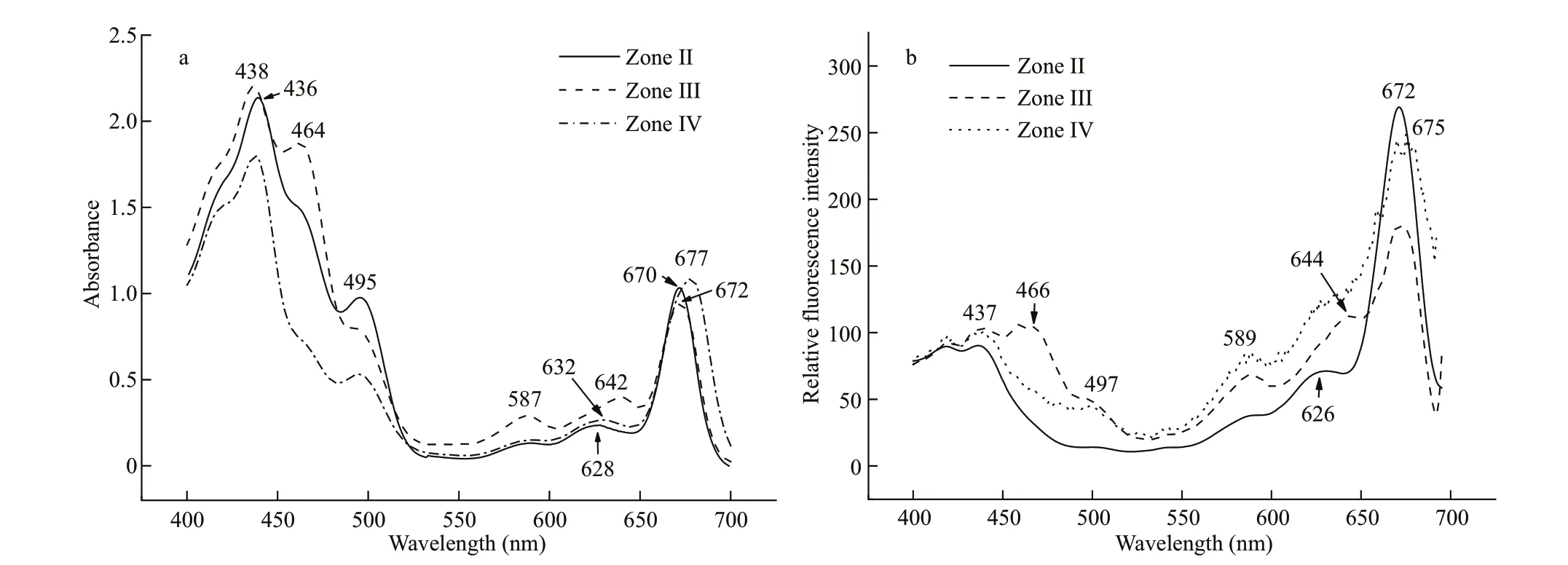
Fig.3 The absorption (a) and f luorescence excitation spectra (b) of pigment-protein complexes isolated by gradient centrifugation at room temperature ( E m=700 nm)
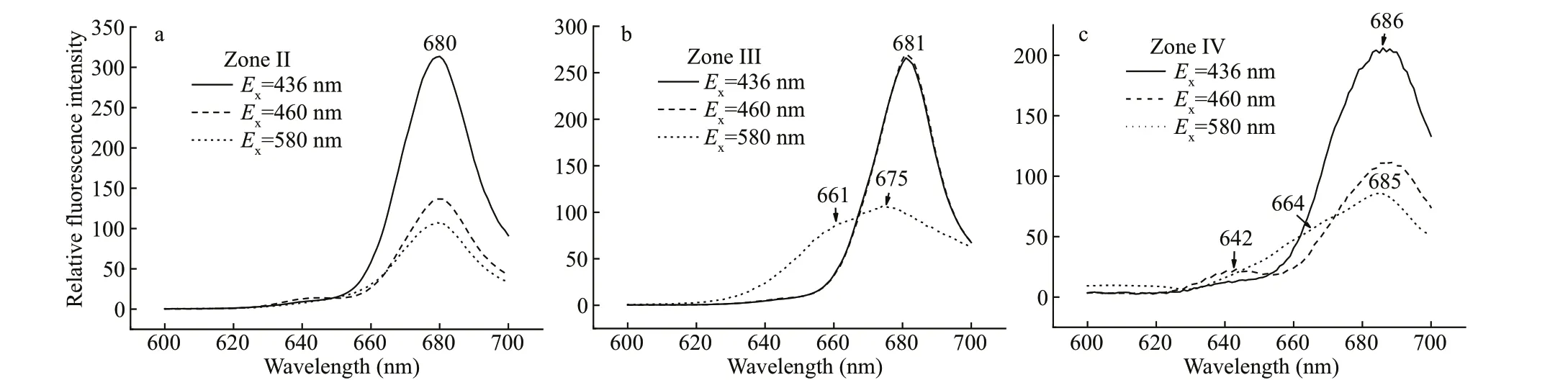
Fig.4 Fluorescence emission spectra of zones isolated by sucrose gradient centrifugation determined at room temperature
The yellow-green band zone Ⅲ at 0.5-mol/L sucrose had no photo-chemical activity. It had two small absorption peaks at 587 and 642 nm that probably corresponded to PC645 in addition to major peaks for Chla(438 and 672 nm) and relative higher amounts of Chlc(464 nm) and carotenoids (495 nm)compared to the other isolated zones (Fig.3a) (Chla/cratio=1.47). The f luorescence emission spectra of zone Ⅲ were unusual. The dominant f luorescence emission excited by either 436- or 460-nm excitation light was at 680-681 nm, similar to that for zone Ⅱ(Fig.4b). However, the emission spectrum excited by 580-nm light showed a peak at 675 nm (684 nm at 77 K, Fig.5a) coupled with a shoulder around 661 nm(minimized shoulder at 660-665 nm at 77 K). The excitation spectra at 700 nm also showed a f luorescence contribution of PC at 589 nm and 644 nm(Fig.3b). Accordingly, zone Ⅲ might be a PC-Chla/c-protein light-harvesting complex, although some free PC components might co-migrate with it (Fig.2a).
Zone Ⅳ was a green band located at 0.6-mol/L sucrose gradient. The absorption spectra showed the dominant absorptions were long wavelength of Chla(438 nm and 676-678 nm) and carotenoids (495-497 nm) (Fig.3a). A small amount of Chlc(464 nm)and trace of PC component also appeared (Chla/cratio=6.62); the latter absorbed mainly around 645 nm and made the Chlcabsorption band in the red region migrate from 625-630 to 632-635 nm. When excited by 580-nm light, zone Ⅳ showed a f luorescent emission coupling of 662-665 nm from PC645 and a peak at 685 nm from Chlaat room temperature(Fig.4c), and 663-668 nm coupled with 680-682 nm at 77 K (Fig.5b), indicating energy transfer from PC to Chlain this complex. This result was conf irmed by the excitation spectra (Fig.3b): the excitation peaks at 589 and 640-644 nm were attributed to PC.Furthermore, zone Ⅳ showed obvious PSⅡ photooxidative activity. These features allowed us to designate zone Ⅳ as an active PC-Chla/c-protein PSⅡ complex. This particle was round in shape with a diameter size about 15-20 nm, according to negative staining and transmission electron microscopy measurement (Fig.6a & b).

Fig.6 Negative staining of zone IV observed by cryogenic transmission electron microscopy
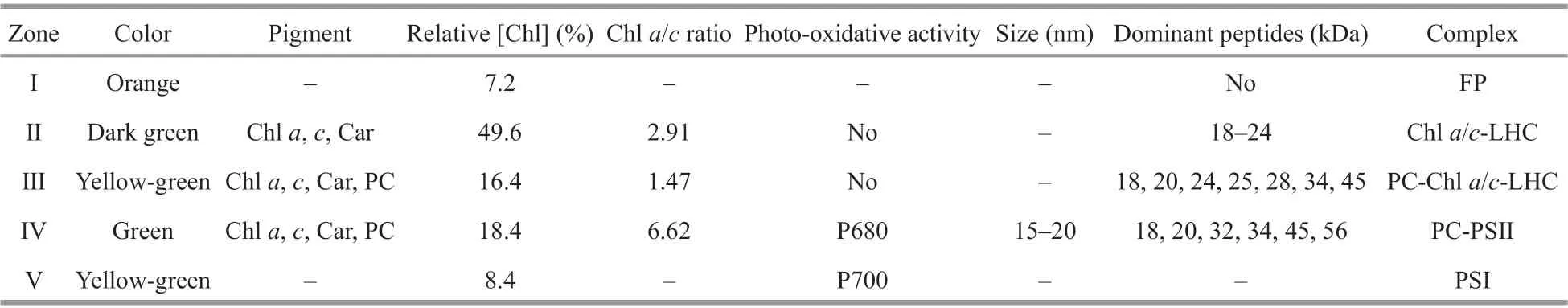
Table 2 The pigment-protein complexes isolated by the f irst step of sucrose gradient centrifugation
Zone V (yellow-green), which appeared at 1.0 mol/L in the sucrose gradient, showed P700 oxidative activity was therefore characterized as an active PSI particle. The results of isolated zones after the f irst steps of sucrose gradient centrifugation are summarized in Table 2.
3.3 Sub-complex isolated by secondary Triton X-100 sucrose gradient centrifugation
Light-harvesting complexes (zones Ⅱ, Ⅲ) and PSⅡparticle (zone Ⅳ) were pooled and subjected to secondary Triton X-100 sucrose gradient centrifugation. Zone Ⅱ-1 migrated at the same position(Fig.7a) and showed similar absorbance and f luorescence features to zone Ⅱ (results not shown).Zone Ⅲ created a new yellow-green band at 0.3 mol/L in the gradient, named as zone Ⅲ-1, while zone Ⅲ-2 appeared at a similar gradient position as zone Ⅲ in the f irst centrifugation (Fig.7b). Zone Ⅲ-2 contained an even higher amount of Chlc(460 nm) and carotenoids (495 nm) than did zone Ⅲ (Fig.8a), while zone Ⅲ-1 was lower. The 642-nm f luorescence emission peak in zone Ⅲ-1 produced by Chlcindicated the partial interruption of energy transfer from Chlcto Chla(Fig.9a). The PC component (585 and 640 nm) remained in zone Ⅲ-2 (Fig.8a), which contributed 665-nm f luorescence emission shoulder except 680-nm f luorescence of Chlaunder excitation by 580-nm light, and 589- and 642-nm f luorescence under excitation at 700-nm emission light (Fig.9b).This result was conf irmed by low temperature f luorescence emission spectra (Fig.10a).

Fig.7 Secondary sucrose gradient centrifugation of zone Ⅱ(a), zone Ⅲ (b), and zone Ⅳ (c) treated with 0.5%Triton X-100 before centrifugation
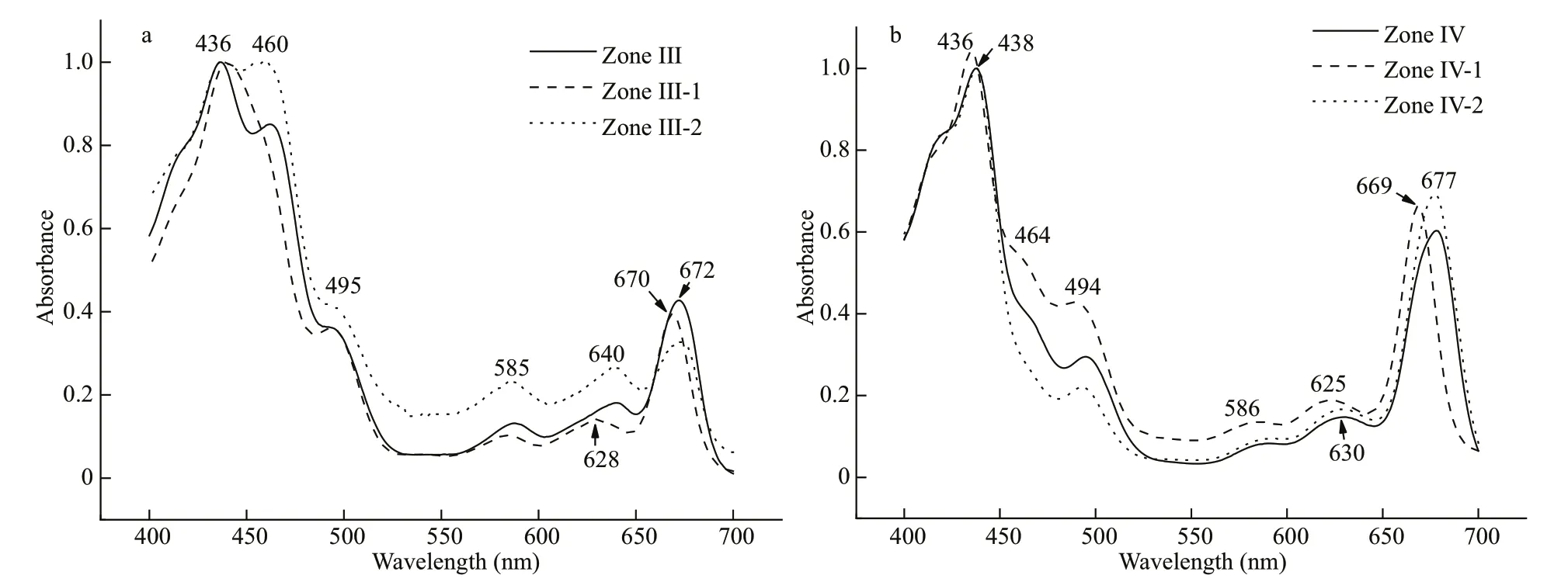
Fig.8 Absorption spectra of zones isolated by secondary sucrose gradient centrifugation after further treatment with 0.5%Triton X-100
Zone Ⅳ also created two bands, named zoneⅣ-1, located at 0.3 mol/L in the gradient, and zoneⅣ-2 at 0.6 mol/L (Fig.7c). Zone Ⅳ-1, containing a small amount of light-harvesting complex stripped from PSⅡ core particle (zone Ⅳ), showed a small amount of Chlc(464 and 625 nm) and carotenoids(494 nm) (Fig.8b). The f luorescence contribution of Chlcand PC were not obvious to 700 nm (Fig.9c).The green band zone Ⅳ-2, containing mainly Chla(438 and 677 nm), small amount of carotenoids(494 nm) and only trace of Chlc(464 nm), showed obvious PSⅡ photo-oxidative activity, and was characterized as a PSⅡ core complex. Signif icantly,zone Ⅳ-2 retained a 660-664-nm and 685-688-nm coupling of f luorescence emission under 580-nm excitation at room temperature (Fig.9d) and 644 nm with 673 nm coupling peak at 77 K (Fig.10b).Therefore, zone Ⅳ-2 appears to be a core complex of PSⅡ still tightly associated with PC components;this f inding has not been reported before in cryptophytic algae.
3.4 Fluorescence dynamics during storage
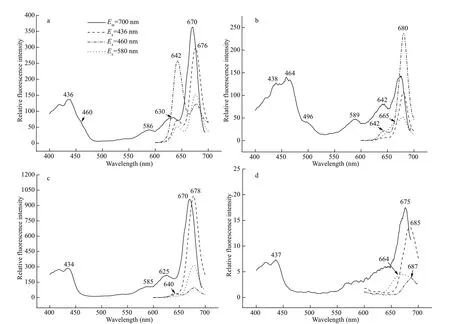
Fig.9 Room temperature f luorescence spectra of sub-complexes isolated by secondary sucrose gradient centrifugation
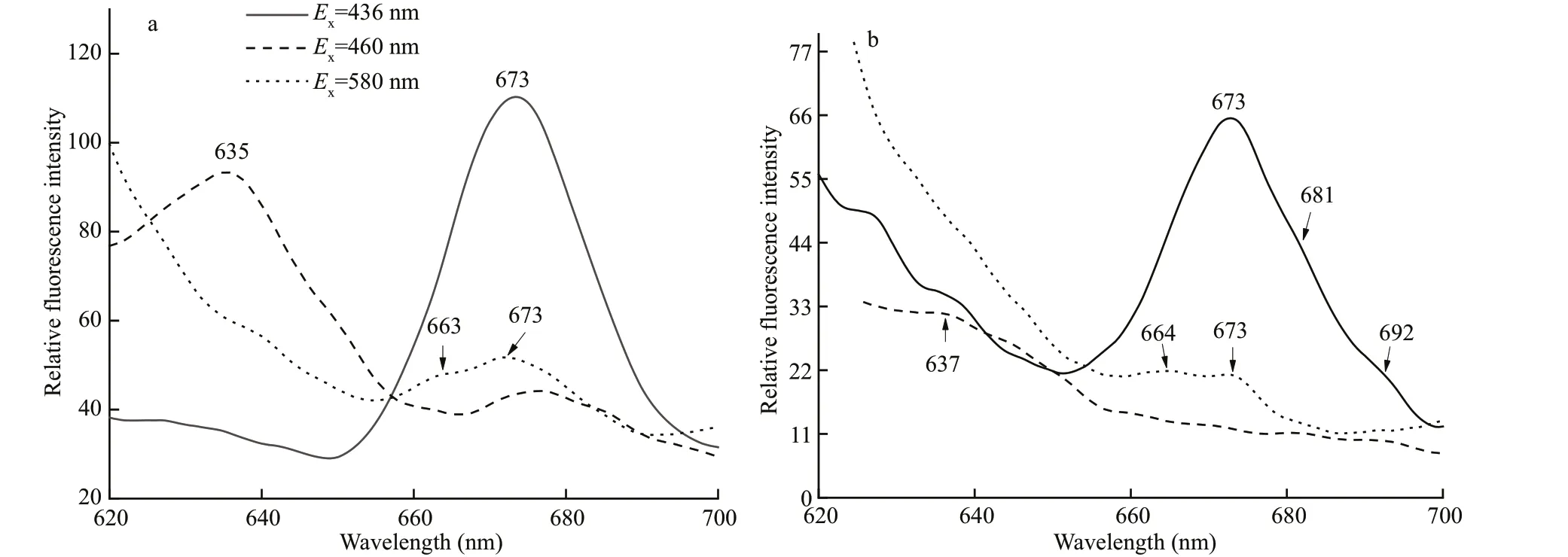
Fig.10 Fluorescence emission spectra of sub-complexes determined at 77 K
Light-harvesting sub-complex zone Ⅲ-2 and PSⅡcomplex zone Ⅳ were collected and stored at room temperature (15-25 ° C) for 7 days. The f luorescence emission spectra when excited with 580-nm light showed that the 680-nm or 685-nm peak emitted by Chlabecame lower during storage, while the 660-nm f luorescence of PC became higher (Fig.11a & b). This result conf irmed that the PC components were combined non-covalently with chlorophyll-protein complexes and were able to transfer excitation energy effi ciently to Chlain both light-harvesting and PSⅡcore complexes. Storage can cause the decomposition of PC from Chl-containing protein complexes, as well as the interruption of energy transfer between PC645 bilins and Chla.
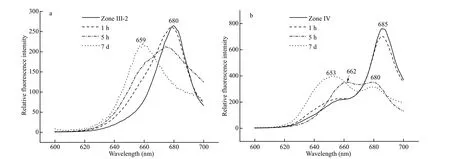
Fig.11 Eff ect of storage on f luorescence of zone Ⅲ-2 (a) and sub-complex zone Ⅳ (b) at room temperature ( E x=580 nm)
3.5 Polypeptide analysis by SDS-PAGE
Zones enriched in PSⅡ and light-harvesting complexes isolated by two steps of sucrose gradient centrifugation and PC samples were subjected to SDS-PAGE. The polypeptide patterns of PC samples revealed the presence of three main polypeptides of relative molecular masses (Mr) approximately 20 and 10 kDa, which were reported as the β and α subunits of PC645 (Lichtlé et al., 1987), and an 8-kDa subunit named as α′ (Fig.12a & b). All three subunits showed pink f luorescence under ultra-violet light after staining by ZnSO4(Fig.12b). Furthermore, some peptides at the 30-40-kDa region also showed pink f luorescence and were considered as α β and β β aggregates(Fig.12b).
The small LHC zone Ⅱ contained mainly peptides of 18-24 kDa (Fig.12) that were considered typical LHC proteins as previously identif ied by the Western blotting (Janssen and Rhiel, 2008). Zone Ⅲ, a largerMrlight-harvesting fraction, comprised a multitude of peptides ranging from under 5 kDa up to 65 kDa,dominated by peptides of 18, 24, 25, 28, 34, and 45 kDa. The sub-complex zone Ⅲ-2 showed a similar composition to zone Ⅲ, but zone Ⅲ-1 was found to consist of peptides of 40-65 kDa, as was sub-complex zone Ⅳ-1. Zone Ⅳ, which was characterized as PSⅡparticles, consisted of three prominent groups of peptides of around 18-28, 30-36, and 40-60 kDa,although some peptides with higherMrwere also detected. Zone Ⅳ-2 contained mostly peptides of 30-66 kDa, which were designated as PSⅡ peptides(Ingram and Hiller, 1983; Lichtlé et al., 1987; Bathke et al., 1999; Janssen and Rhiel, 2008), and two smallerMrpeptides of 16 and 20 kDa. The 18-28-kDa LHC proteins of zone Ⅳ were diminished in zone Ⅳ-2 because of the further resolution and separation. ZoneⅣ-2 was characterized as a core PSⅡ complex with a small amount of associated LHC remaining.Signif icantly, zones Ⅲ, Ⅳ, Ⅲ-2, and Ⅳ-2 also contained an extra peptide of 20 kDa, and showed the same migration position and pink f luorescence under ultra-violet as the β subunit in PC (Fig.12b).Sometimes the 20-kDa subunits aggregated as a β β complex and showed another less intense f luorescent band at around 40 kDa (Fig.12). In contrast, the α subunits were not noticeable in these Chl-containing complexes, when stained with either ZnSO4or silver(data not shown).
4 DISCUSSION
Cryptophytes are unique organisms that contain both PBPs and chlorophyll-protein complexes as their light-harvesting structures. Investigations on the photosystems of cryptophytes have focused on isolated PBPs rather than on Chl-containing proteins.Fractions enriched in PSI, PSⅡ, and Chla/c2-protein complexes have been reported following either gel electrophoresis or sucrose density centrifugation(Ingram and Hiller, 1983; Lichtlé et al., 1987; Janssen and Rhiel, 2008). However, the potential subcomplexes of these fractions have not been reported.
The latest model of cryptophytic algal photosystems suggested that the cryptophytic PBPs pack densely and freely inside the lumen as individual heterodimers displaying no relation to each other or to the thylakoid membrane (Doust et al., 2004; Mirkovic et al., 2017).An active PE-PSⅡ complex fromCryptomonasrufescensand a novel PC-Chla/c2-protein complex inC.placoideawere reported by Lichtlé et al. (1987)and Chen et al. (2007), respectively. Here we provide evidence for two kinds of PC-PSⅡ complexes as well as two PC-LHC complexes. Fluorescent spectra and a dynamic study during storage conf irmed that the PC645 was combined non-covalently with chlorophyll-protein complexes in these isolated complexes. Effi cient energy transfer was observed from PC645 to Chlain both PSⅡ particles and LHCs.These results demonstrate that PC645 inC.placoideais structurally but not preferentially combined with a LHC and the PSⅡ reaction center. The excitation energy absorbed by PC645 can be directly transferred to Chlain the PSⅡ reaction center or via a Chla/cprotein complex. The present work provides a series of PC-chlorophyll-protein complexes which should serve as useful materials for further investigation of structural architecture, energy transfer, and the molecular arrangement of photosynthetic systems in cryptophytic algae.
From previous peptide analysis and western blotting (Janssen and Rhiel, 2008), the proteins of 18-22 kDa, which we found as the dominant peptides in zones Ⅱ, Ⅲ, and Ⅲ-2, are considered typical LHC proteins of cryptophytic algae. The peptides of 30-34 kDa enriched in zone Ⅳ and sub-complex zoneⅣ-2, which are characterized as PSⅡ particles and reaction cores, respectively, are considered the core peptides D1/D2 of PSⅡ. In the previous report, the proteins at 45-55 kDa were designated as inner antenna peptides of PSⅡ; if the PSⅡ in cryptophytes has a similar inner antenna structure to that in higher plants and green algae, it has not yet been well def ined.Accordingly, in the present work, zone Ⅱ (containing 18-22-kDa peptides) was considered as a lowMrLHC associated peripherally with the PSⅡ core. ZoneⅢ (an intact LHC with some PSⅡ components still attached), was divided into two parts (Fig.7): the inner antenna zone Ⅲ-1 containing inner peptides and less Chlc, and zone Ⅲ-2 containing unusually high amounts of auxiliary pigments Chlcand PC. ZoneⅢ-2 was proposed as a large-size LHC located at the outer part of PSⅡ. Zone Ⅳ is a larger PSⅡ particle with a diameter of about 15-20 nm, similar in size to the PSⅡ dimer of blue algae (Zhao et al., 2020), but smaller than the PSⅡ-LHCⅡ particle of spinach that was reported as 26 nm× 14 nm×11 nm (Wei et al.,2016) because of the attachment of two tightly associated LHCⅡ trimers along the long axis.Therefore, zone Ⅳ was considered as a PSⅡ particle surrounded by a certain amount of antenna LHC.Zone Ⅳ-1, derived directly from zone Ⅳ that contained active PSⅡ particles, was considered as the inner antenna of PSⅡ that might be located closer to the reaction core complex than are the proteins of zone Ⅲ-1. The other part of zone Ⅳ, named zoneⅣ-2, was a typical core complex of PSⅡ with traces of LHC peptides and PC subunits still attached. The proposed relationship of these isolated complexes and sub-complexes is summarized in Fig.13.
Each PC645 heterodimer contained three diff erent types of bilins (Wedemayer et al., 1996; Collini et al.,2010; Overkamp et al., 2014): one 15,16-dihydrobiliverdin (DBV) and two phycocyaninbilins (PCB) in each β subunit, and one mesobiliverdin (MBV) in each α subunit. MBV exhibits a strong red absorption at 697 nm and had been considered as the terminal emitter bilin in the PC645 heterodimer (Doust et al., 2006). But ultrafast spectral analysis elucidated that the shoulder at 625-633 nm and the red most peak at 640-645 nm were assigned to four PCBs. The PCB β 82 was assumed to be the f inal emitter and responsible for the characteristic f luorescence emission of PC645 at 660-665 nm (Mirkovic et al., 2007, 2017; Marin et al., 2011). In our case, two small f luorescent α subunits withMrof 8 kDa and 10 kDa were detected in addition to the 20-kDa β subunit in our purif ied PC645 sample as described in previous reports.However, in our isolated PC-Chl-protein complexes,only β subunits rather than α subunits could be detected. These Chl-containing complexes were obtained from well-washed thylakoids, a process that might cause the loss of small α subunits. When the excitation wavelength was 580 nm, these complexes exhibited the 670-685-nm f luorescence emitted by Chla, coupled with 660-665-nm emission from the f inal emitting bilins of PC645. Therefore, the demonstration of 20-kDa β subunits in these PC-Chlprotein complexes provides direct evidence supporting the view that the β subunit corresponds to the terminal emission of PC645. These results also imply that PC645 might attach to the insoluble thylakoid components with a specif ic form and angle. The β subunit might play an important role in the combination of PC645 with Chl-containing complexes, although the α subunits also contribute importantly to interactions in the quaternary structure of PC645 and energy transfer between PC645 and chlorophylls (van der Weij-De Wit et al., 2006).Further investigation on the precise structure and energy transfer of these novel PC-Chl-protein complexes will help in elucidating the characteristics and function of these subunits in the structural relationship between PBPs and Chl-protein complexes in cryptophytes.
5 DATA AVAILABILITY STATEMENT
The datasets generated and/or analyzed in the current study are available from the corresponding author upon reasonable request.
 Journal of Oceanology and Limnology2022年2期
Journal of Oceanology and Limnology2022年2期
- Journal of Oceanology and Limnology的其它文章
- Identif ication of Antarctic minke and killer whales with passive acoustic monitoring in Prydz Bay, Antarctica*
- Eff ects of dissolved oxygen and nutrients from the Kuroshio on hypoxia off the Changjiang River estuary*
- Methane in the Yellow Sea and East China Sea: dynamics,distribution, and production*
- Longitudinal genetic analysis of growth-related traits in red swamp crayf ish Procambarus clarkii (Girard)*
- Early life migration and population discrimination of the small yellow croaker Larimichthys polyactis from the Yellow Sea: inferences from otolith Sr/Ca ratios*
- A new oil spill detection algorithm based on Dempster-Shafer evidence theory*