
10种月见草属植物染色体核型分析
2022-04-02 06:10孙浩男李明阳刘冬云
西北林学院学报 2022年2期
孙浩男,郑 妍,李明阳,刘冬云,王 鑫,田 琳
(河北农业大学 园林与旅游学院,河北 保定 071000)
柳叶菜科(Onagraceae)月见草属(OenotheraL.)植物多为一、二年生或多年生草本,原产美洲温带至亚热带地区,全属约150多种,我国各省均有栽培引种,部分沦为逸生。月见草属植物适应性强,部分种类如长果月见草(O.marcrocarpa)、美丽月见草(O.speciosa)等具有花大色香、花量丰富、花期长的特点,观赏价值极高,是出色的地被植物,常用于园林绿化。在我国东北地区,常大面积栽培夜来香(O.biennes)作为油用植物,经过常年的选育,已从野生种及野生转栽培种中筛选出一批含油量较高的品系,其种子油中富含的γ-亚麻酸具有一定抗炎、降血脂的功效,具有一定药用价值。除此,月见草属植物也具蜜源、酿酒、纤维和香精等用途[1-3]。
核型参数是植物分类、鉴定的重要指标之一,通过核型分析,能从细胞学方面研究植物的遗传多样性[4]。目前,核型分析已成功在菊花[5]、百合[6]和蝴蝶兰[7]等观赏花卉上得到应用。从现有报道来看,我国月见草属植物资源的质量参差不齐,更是存在同种异名的现象[8],极大地影响了我国月见草属植物进一步的研究和应用。目前,月见草属的植物虽然已有细胞学标记方面的研究,但其主要集中在其染色体环、染色体连接现象,及其染色体的异位突变性[8-10]等,而有关染色体核型的极少,更是缺乏同属间物种染色体核型的比较,截至目前,国内仅有夜来香和小花山桃草(O.parviflora)2个种的核型进行了报道[2,11-12]。如今,随着月见草属植物在我国不断归化,对我国常见的月见草属植物进行细胞学方面的研究,并明确其染色体形态及核型特征具有十分重要的意义。因此,采用常规压片法,对国内常见的10个月见草属植物的染色体数目及核型等参数进行了研究,旨在为其资源鉴定、利用和细胞学研究等方面提供更多理论基础。
1 材料与方法
1.1 材料
试验材料为10种月见草属植物的成熟种子,A-H采自河北绿地花卉景观研创中心引种圃,I和J分别引自英国Thompson&Morgan和Plant Word Seeds园艺公司,其他信息详见表1。
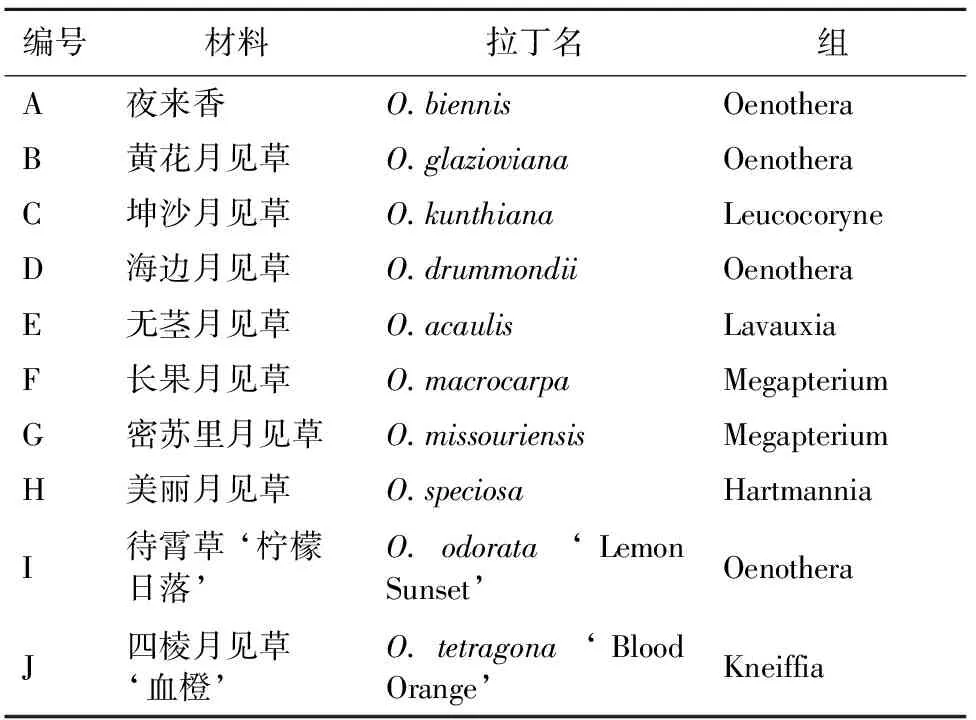
表1 供试月见草材料的基本信息
1.2 方法
使用垫有润湿滤纸的干净培养皿在光照培养箱中对种子进行催芽生根,其余方法参考孔令月等[4]的研究方法,并进行改良:切取根尖先端1~3 cm置于冰水混合物中预处理24 h;随后使用卡诺氏固定液浸泡20~24 h;用蒸馏水冲洗3~5次;使用1 mol/L的稀盐酸在60 ℃水浴锅中进行解离,解离时间依据根尖大小进行调整;再次用蒸馏水冲洗3~5次,吸取多余水分后置于载玻片上,用镊子挑取根尖先端1~5 mm,捣碎后滴入2~3滴改良卡宝品红染色10~30 min;盖上盖玻片,使用带有橡皮头的铅笔轻敲促使细胞分散,至根尖整体呈云雾状;使用Leica DM4000荧光显微镜观察制片,对染色体分散良好且处于中期分裂相的细胞进行观察拍照。
核型数据测量和处理等参考戴小红等[13]的研究,观察和统计至少30个有丝分裂中期相细胞进行染色体计数,参考李懋学等[14-15]对5个分散良好且数目清晰的中期分裂相进行核型分析,染色体类型按照A.Levan等[16]的标准,核型类型分析参照G.L.Stebbins等[17]的标准进行,核型不对称系数按照Arano[18]的方法计算。参考孙桂芳等[19]的方法,以平均臂比和核型不对称系数制作核型坐标图。参考秦丹等[20]的方法制作染色体核型聚类图。
2 结果与分析
2.1 染色体数目与倍性
根据图1和表2中的染色体形态与染色体参数进行同源染色体配对,其中,H细胞中的28条染色体可配成7对,为四倍体,其余9种月见草的14条染色体均可配成7对,为二倍体,染色体基数均为x=7。
2.2 染色体核型分析
由表2和表3可知,10种月见草的染色体组成类型单一,仅含有m和sm 2种类型,A和E全部由m型染色体组成,其余由m型和sm型2种染色体组成。染色体相对长度相差不大,分别为A(12.99%~15.67%)、B(11.79%~20.01%)、C(12.07%~17.65%)、D(11.17%~20.21%)、E(12.96%~16.03%)、F(11.77%~16.16%)、G(11.99%~18.09%)、H(12.98%~15.33%)、I(12.28%~16.77%)、J(12.22%~16.19%)。除A外,在其余9种月见草中发现随体,均处于染色体的短臂上,随体数量上存在一定差别,4个(C、G、I、J)为1条,2个(D、H)为2条,3个(B、F、G)为3条,随体所在的染色体序号也存在差别,分别为B(4号,6号,7号)、C(7号)、D(4号,7号)、E(5号)、F(4号,5号,6号)、G(1号,4号,7号)、H(1号,7号)、I(4号)、J(7号)。在H中发现1条B染色体,其余材料中则未发现B染色体。核型类型存在3种,A和E为1A型,I为3A型,其余均为2A型。核型不对称系数介于56.79%~65.43%,整体较为对称,由小到大可排序为:A、E、C、J、H、D、B、G、F、I。平均臂比为1.33~1.92,最长染色体/最短染色体值介于1.21~1.81,臂比大于2的染色体占染色体总数的百分比为0%~57.14%。
2.3 染色体进化趋势及聚类分析
依据核型坐标图(图2)可知,I处于坐标图的最右上角,最为进化,而A和E处于左下角,较为原始。根据聚类结果,在遗传距离为1.0时,10种月见草被划分为4类:A、C、E为一类;D、H、J为一类,F、G、I为一类;而B为单独一类。当遗传距离为2.0时,B与F、G、I聚为一类;当遗传距离为3.0时,10种月见草明显划分为2大类,其中B、F、G、I为一类,其余6种月见草聚为一类;当遗传距离为25.0时,所有月见草聚为一类。sect.Oenothera的4个材料(在坐标图与聚类图中的位置较为分散,平均臂比和核型不对称系数上A 有关核型试验细胞的最佳解离时间,因植物物种本身和取材部位等而不同[4-5,19-20]。本试验解离部位为月见草种子萌发后的根尖部位,在实际操作中发现,不同种类的月见草在最佳解离时间上存在一定差异。如坤沙月见草和美丽月见草等一些种子极小的材料(直径1 mm),解离时间过长会导致根尖部位难以分辨甚至解离过度,将其解离时间保持在4~5 min,得到的压片细胞分散情况极好;而无茎月见草(直径2 mm)和长果月见草(直径3~5 mm)等在6~8 min较为合适,这与周俊英[11]的研究方法不同。 月见草属染色体多倍化发生较少,但18个组中有10个组存在这个现象[21]。从现有报道来看,月见草属染色体基数为x=7,月见草属植物的染色体存在2n=14,28,42,56[22]。本试验有9种材料的染色体为2n=2x=14,H美丽月见草为2n=4x=28,与L.L.Wolin等[23]报道的(2n=2x=14,2n=4x=28,2n=6x=42)部分一致,未发现其他倍性的多倍体、非整倍体及混倍体。B染色体对于增强植物的抗逆性有一定作用[24-25],美丽月见草具有适应性强,生长旺盛的特点,大量地应用到了我国城市绿化中,B染色体可能增强了其对环境的强适应性,但在加拿大一枝黄花(Solidagocanadensis)[26]、薇甘菊(Mikaniamicrantha)[27]的报道中,B染色体的出现频率也与强入侵性相关。 一般认为,核型的进化趋势为染色体的对称程度和倍性的提高,依据核型不对称系数可初步分析物种间的进化趋势[28]。在柳叶菜族(Tribeonagreae)中,地烟花属(Gayophytum)和月见草属的所有7对染色体是在减数分裂中交换臂形成,并作为一个单位进行分离,这可能是造成月见草属植物核型较为对称的主要原因[29]。从先前报道过的月见草属的夜来香和小花山桃草[12]来看,两者核型均为1A型,属于极为原始的类型。而本试验的10种月见草材料核型也多为1A型和2A型,A夜来香的核型与周俊英[11]所得出的1A型一致,核型公式也相同,但在核型不对称系数等核型参数及随体方面存在一定差距。在核型方面上看,美丽月见草的2A型虽为原始,但染色体倍性方面较为进化。月见草属植物具有极强的染色体的异位突变性,永久易位杂交品系的同源重组和染色体的自由分离会受到抑制而不易产生后代[8],而待宵草 ‘柠檬日落’的核型为3A型,其在品种选育中可能发生了其他类型染色体变异。 根据POWO(plants of the world online)上最新登录的国际月见草属植物名单,密苏里月见草已修订并为长果月见草。本试验中,F长果月见草和G密苏里月见草在核型类型、核型公式、平均臂比及不对称系数等核型参数上均相差无几,结合核型散点图和聚类分析图的结果,两者在细胞学方面具有极为紧密的亲缘关系。我国月见草属植物种类众多,据不完全统计,截至2006年,我国引入的月见草约20种[3],而后十几年内名录并无更新。此次供试的月见草属植物种类有限,但在各核型参数上存在差别,染色体的异位突变性可能是该属植物核型多样性的主要原因,染色体整体上较为对称,核型属于较为原始的类型。除夜来香外,其余9种月见草的核型均为国内首次报道。这些数据从细胞学标记方面为我国月见草属植物的鉴定和分类提供了一定理论基础。但是不同产地的材料也可能存在不同的核型特征,这需要收集更广泛的资源来进行对比探讨。3 结论与讨论
猜你喜欢
云南医药(2021年6期)2022-01-08
雪豆月读·低年级(2021年9期)2021-09-13
川北医学院学报(2021年6期)2021-07-13
小星星·阅读100分(低年级)(2019年8期)2019-09-16
作文·初中版(2019年8期)2019-09-09
中国实用医药(2016年36期)2017-06-20
诗潮(2016年5期)2016-05-14
创新作文(5-6年级)(2015年9期)2015-10-21
湖北农业科学(2015年17期)2015-10-09
汽车维护与修理(2015年2期)2015-02-28
