
基于生态酶化学计量研究小陇山不同植被类型土壤有机碳对微生物磷限制的影响
2022-04-02 05:53杨海裕郑子龙刘小林李惠萍高鹏程
西北林学院学报 2022年2期
杨海裕,郑子龙*,刘小林,李惠萍,高鹏程
(1.甘肃省小陇山林业科学研究所,甘肃 天水 741020;2.甘肃小陇山森林生态系统国家定位观测研究站,甘肃 天水 741020;3.甘肃省次生林培育重点实验室,甘肃 天水 741020;4.西北农林科技大学 资源环境学院,陕西 杨陵 712100)
土壤是森林生态系统的重要组成部分,起着为地上植被提供营养以及协调水、肥、气、热,保障植被生存生长起着重要作用[1-2]。森林土壤由固、液、气三相构成的复杂物质体系,土壤物理、化学以及生物学特性可以决定森林土壤质量[3-4]。土壤酶在森林土壤中扮演十分重要的角色,土壤微生物通过细胞胞外酶降解复杂的有机化合物来获取其自身所需的各种营养[5-6],因此土壤酶活性成为评价土壤肥力的一个重要指标[7-8]。但是通过对单种酶的分析测定仅仅能揭示样品之间绝对活性水平的差异,几乎不能为有关微生物群落整体行为或营养状况提供信息。相反,与C、P以及C和N循环相关酶的相对活性可以揭示C、N和P获取相对资源分配的差异[9]。生态酶化学计量学代表了微生物能够吸取养分的能力,因此可以用作评估生态系统能量流、微生物对养分的需求以及土壤养分供应的重要指标[10-11]。生态酶化学计量分析已用于确定半干旱地区农田生态系统以及高山和沙漠盆地生态系统中的C、N、P限制[12-13],但较少采用生态酶化学计量法来研究森林生态系统中微生物的代谢特征。
目前对小陇山林区植被类型的研究多集中在土壤理化性质和土壤肥力方面,而对微生物的代谢特征等生物学性质研究较少。本试验在甘肃省小陇山森林生态系统国家定位观测研究站(以下简称:小陇山生态站)沙坝监测区选取锐齿栎纯林与锐齿栎混交林进行生态酶化学计量分析研究,旨在探讨不同植被类型对土壤理化性质及土壤酶活性的影响,探究土壤中理化性质的改变对微生物代谢限制的影响,评估各植被类型中微生物生长的C、N、P限制。
1 材料与方法
1.1 试验区概况
试验地位于甘肃省小陇山森林生态系统国家定位观测研究站沙坝监测区,地处天水市秦州区娘娘坝镇境内(105°54′E,34°07′N)。海拔1 550~2 100 m,平均坡度36°~40°,土壤是以山地棕壤与山地褐土为主。属于大陆性季风气候,为暖温带湿润区,年平均气温7.2 ℃,最高气温30.3 ℃,最低气温-22.4 ℃;年均降水量757 mm;平均相对湿度78%;平均年日照时数1 553 h;平均年蒸发量1 012.2 mm;≥10 ℃有效积温2 480 ℃;初霜期10月16日,终霜期5月4日,无霜期154 d。
1.2 试验设计
在小陇山生态站沙坝监测区选择锐齿栎纯林、锐齿栎混交林2种森林类型,设置4块面积为20 m×20 m固定观测样地。
1.3 样品采集和参数计算
2020年8月在样地选取8个样点,清除表层枯枝落叶及表层浮土后,从土壤剖面的0~10、10~20、20~30、30~40、40~60 cm处每层采集3个土壤样品。在每个样地随机采取8个土样,并充分混匀成1个样品。去除根、落叶、杂物和枯枝后,将每个混匀样品分为2部分以备将来分析。一部分通过2.0 mm的筛子,并保存在4 ℃下,用于在1周内分析细胞外酶的活性。另一部分风干用来分析其理化性质。土壤酶活性采用微孔板荧光法测定与C、N、P循环相关的5种酶,其中与碳循环相关的酶是β-1,4-葡萄糖苷酶(BG)与β-D-纤维二糖水解酶(CBH);与N循环相关的酶是β-1,4-N-乙酰氨基葡糖苷酶(NAG)与L-亮氨酸氨基肽酶(LAP);与P循环相关的为酸性磷酸酶(AP)。
微生物代谢限制的定量:微生物代谢限制是通过计算所有未转化酶活性数据的矢量长度和角度来量化。C限制用矢量长度表示,计算方法为x2和y2之和的开平方,其中x表示(BG+CBH)/(BG+CBH+AP);y代表(BG+CBH)/(BG+CBH+NAG+LAP)。矢量角(α)表示微生物代谢受N或P限制,即用原点到点(x,y)延长线的反正切函数表示。
α=degrees(atan2(x,y))
1.4 统计分析
使用Microsoft Excel 2016 软件进行数据处理;使用R v.3.6.1软件包执行相关性分析,使用IBM SPSS Statistics 21.0进行其他统计分析;使用Amos v.24构建结构方程模型(SEM)。
2 结果与分析
2.1 不同植被类型下的土壤理化性质和养分化学计量比
通过对2种植被类型不同土层深度土壤养分测定结果分析可以看出,不同植被类型和土层深度对土壤养分含量影响很大(P<0.05,表2)。其中有机C与全N含量随土层深度的增加而降低,而锐齿栎混交林的平均SOC和TN含量分别比锐齿栎纯林高21.1%和17.3%。在0~20 cm土层,锐齿栎混交林的TP含量比锐齿栎纯林处理高16.3%,但在10~40 cm土层的平均TP含量比锐齿栎纯林低36.9%。各植被类型之间的营养元素化学计量存在显著差异(P<0.05)。在0~10 cm土层,锐齿栎纯林的C∶P(120.8±4.6)与N∶P(12.3±0.9)最高,分别为120.8和4.6。锐齿栎混交林的C∶P随土层深度的增加而减小;各植被类型的N∶P随土壤深度增加呈下降的趋势。在锐齿栎混交林下,各土层的C∶P与N∶P比率均比锐齿栎纯林高51.0%与42.9%。
2.2 不同植被类型下的土壤酶活性和生态酶化学计量比
通过对2种植被类型土壤中与C、N、P循环相关酶的活性及其化学计量比进行分析(P<0.05,表3),可以看出,2种植被类型与C、N、P循环相关酶活性均随土层深度增加不断降低;锐齿栎纯林的各土层平均BG+CBH与AP含量较锐齿栎混交林分别提高了16.6%和32.6%,最大值分别为(1 141.6±78.1)nmol·g-1·h-1和(1 417.7±87.7)nmol·g-1·h-1,而锐齿栎混交林各土层的平均NAG+LAP含量较锐齿栎纯林提高了10.3%。锐齿栎纯林与锐齿栎混交林的(BG+CBH)∶(NAG+LAP)随土层深度的增加呈先增加后降低的趋势,在20~30 cm土层达到最高,最大值分别为2.2和1.6。锐齿栎纯林与锐齿栎混交林的(BG+CBH)∶AP随土层深度的增加呈下降的趋势,锐齿栎混交林的平均(BG+CBH)∶AP较锐齿栎纯林提高了14.4%;锐齿栎纯林与锐齿栎混交林的(NAG+LAP)∶AP随土层深度的增加不断下降,最大值分别为0.6和0.8,且锐齿栎混交林的平均各土层(NAG+LAP)∶AP比率显著高于锐齿栎纯林0.5倍。
2.3 不同植被类型土壤细胞外酶化学计量学的载体特征
如图1A所示,生态酶的化学计量学载体特征在不同植被类型和土层之间有所不同。所有采样数据点均在(1∶1)直线上方,从而表明土壤微生物群落受到较强的P限制。不同的植被类型和土层对微生物的代谢特性具有强烈的交互作用(P<0.05,图1B和图1C)。
除10~20 cm,锐齿栎纯林各土层的矢量长度高于锐齿栎混交林(图1B),锐齿栎纯林各土层的矢量长度较锐齿栎混交林提高了5.4%;且锐齿栎纯林各土层的矢量角度均显著高于锐齿栎混交林(P<0.05,图1C)。但是,微生物的碳限制(矢量长度)与微生物的磷限制(矢量角度)没有显著的线性关系(图1D)。线性回归分析还表明,土微生物C限制随pH的升高而降低(P<0.05,图2)。土壤微生物P限制随着容重的升高而增加,但随着有机质、全N、速效N、速效P、C∶P、N∶P、pH和土壤含水量的升高而降低(P<0.05,图3和图4)。
SEM分析确定了土壤物理特性,总有效养分含量和养分比对微生物C和P限制的直接和间接影响。SEM分析微生物养分限制与土壤理化特性之间的关系(长度,卡方=6.380,P=0.896;角度,卡方=8.549,P=0.741)。pH与土壤含水量和速效N呈负相关,与土壤容重和全N呈正相关;土壤容重与有机质、全N、速效N呈负相关;速效N与土壤含水量、有机质、全N和全P呈正相关;而pH对微生物C限制具有消极的总体影响(图4)。此外,土壤含水量与全N、全P和速效N呈正相关,而与土壤容重呈负相关;土壤容重与有机质和速效P呈负相关;全氮与有机质、速效N和速效P呈正相关。土壤容重和全P对微生物P限制具有积极的总体影响,而速效P和有机质对微生物P限制具有消极的总体影响(图4)。
3 结论与讨论
3.1 讨论
3.1.1 不同植被类型对土壤养分的影响 不同植被类型会改变土壤中养分含量,并极大地影响养分的有效性,从而导致微生物代谢受到C和P的相对限制[15]。锐齿栎混交林输入了更多的凋落物,增加了腐殖质层厚度;与锐齿栎纯林相比,土壤中有机C与全N含量分别提高了8.3%与7.3%,这与王强等[16]、L.Huo等[17]的研究结果一致。此外本研究发现,土壤有机质、全N与有效P含量与坡度呈负相关关系,与黄安香等[18]研究结果相似。李昌兰等[19]的研究表明,土壤养分N、P、K流失分为地表流失和地下流失,主要的影响因子为坡度,表现为随着坡度的增加流失量增加。有研究结果表明,受风向、地表径流的影响,坡度越大,土壤N、P、K含量相对越低[20]。本研究表明锐齿栎纯林39°坡土壤肥力低于锐齿栎混交林33.5°坡(表1与表2)。微生物C限制与SOC、TN和Olsen-P无显著线性关系(P>0.05;图2)。微生物P限制与有机C、全N、速效N和速效P呈负相关(P<0.05;图3)。因此,有机C、N、P对微生物养分限制的影响不同。本研究还发现,与锐齿栎纯林相比,锐齿栎混交林平均各土层速效N含量之间无显著差异,有效P含量提高了116.6%(表2),而全P下降了17.9%,这是由于有机C分解后增加了土壤中的有效P。研究结果与Ma等[13]的研究结果相似。C∶P和N∶P比率与微生物P限制相关(P<0.05;图3),从而表明养分化学计量很大程度地影响了微生物对养分的吸收能力。微生物的养分需求是由微生物生物量的元素化学计量学相对于营养物质的环境利用决定的[21]。土壤水分是能够影响土壤结构和活性的重要因素[22],良好的水分条件有利于土壤中有机质和全N的分解,在本研究中,锐齿栎纯林较少的C和N的输入,同时锐齿栎纯林提供了良好的水分条件,造成土壤中有机C含量下降(表2、表4),限制了微生物的生长[9,14]。

表1 样地信息
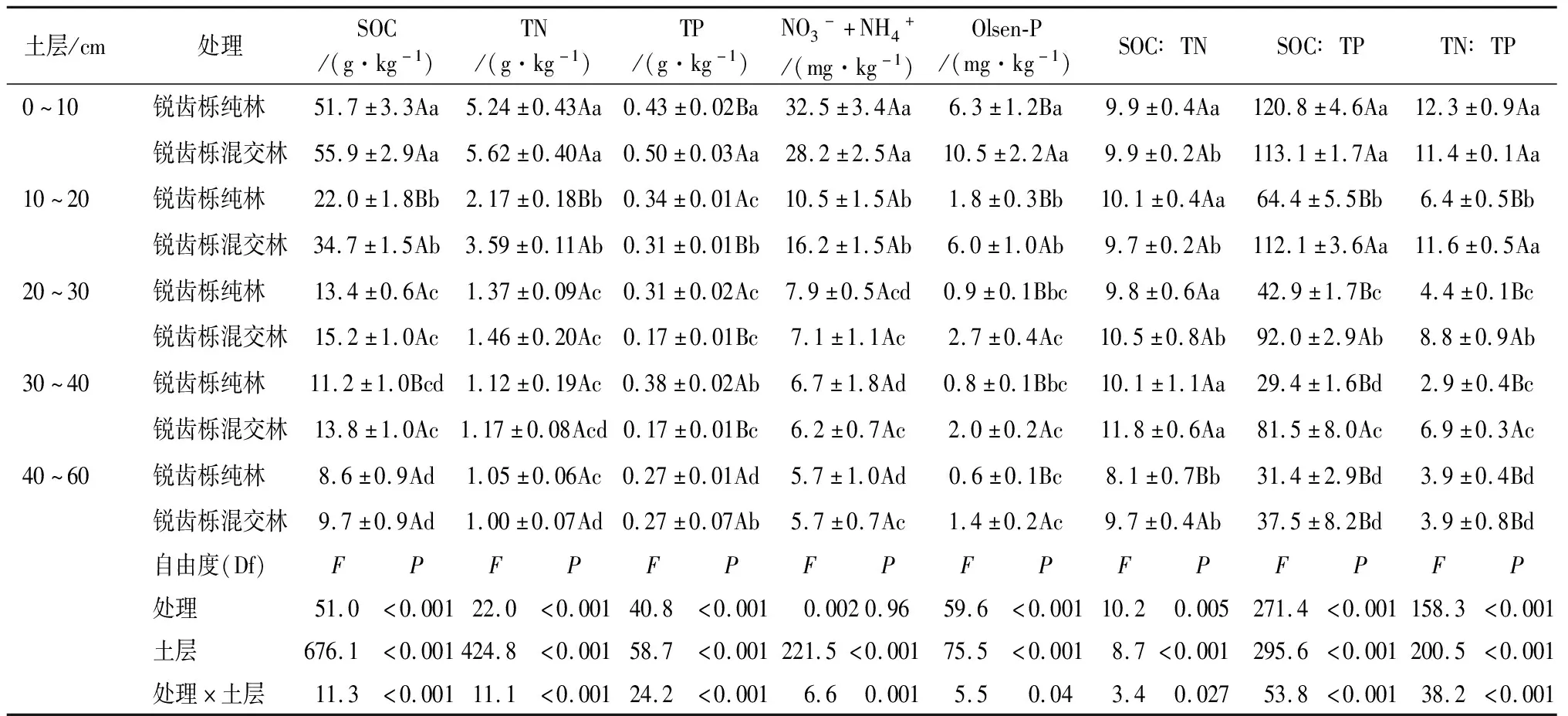
表2 不同植被类型下土壤养分和养分比的双向方差分析
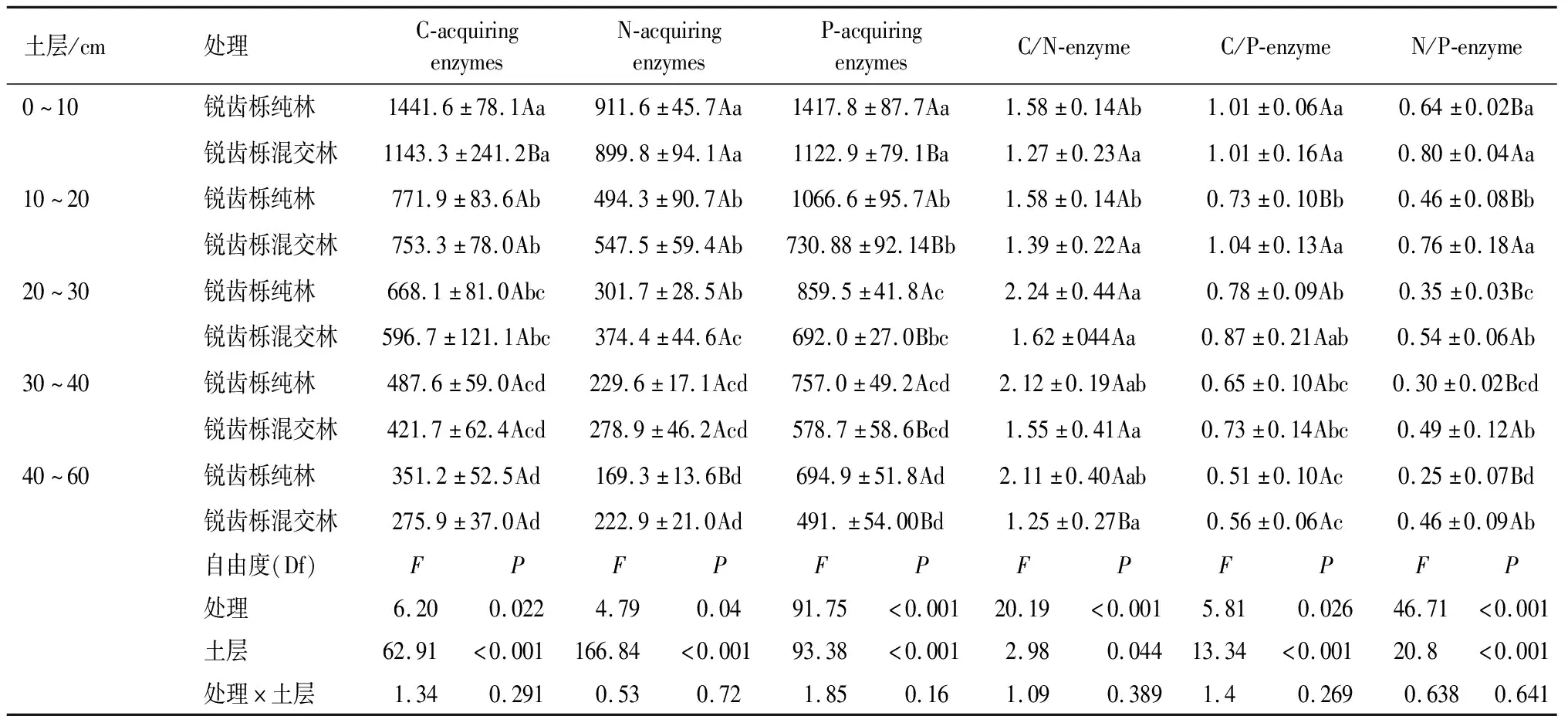
表3 不同植被类型下细胞外酶活性的双向方差分析
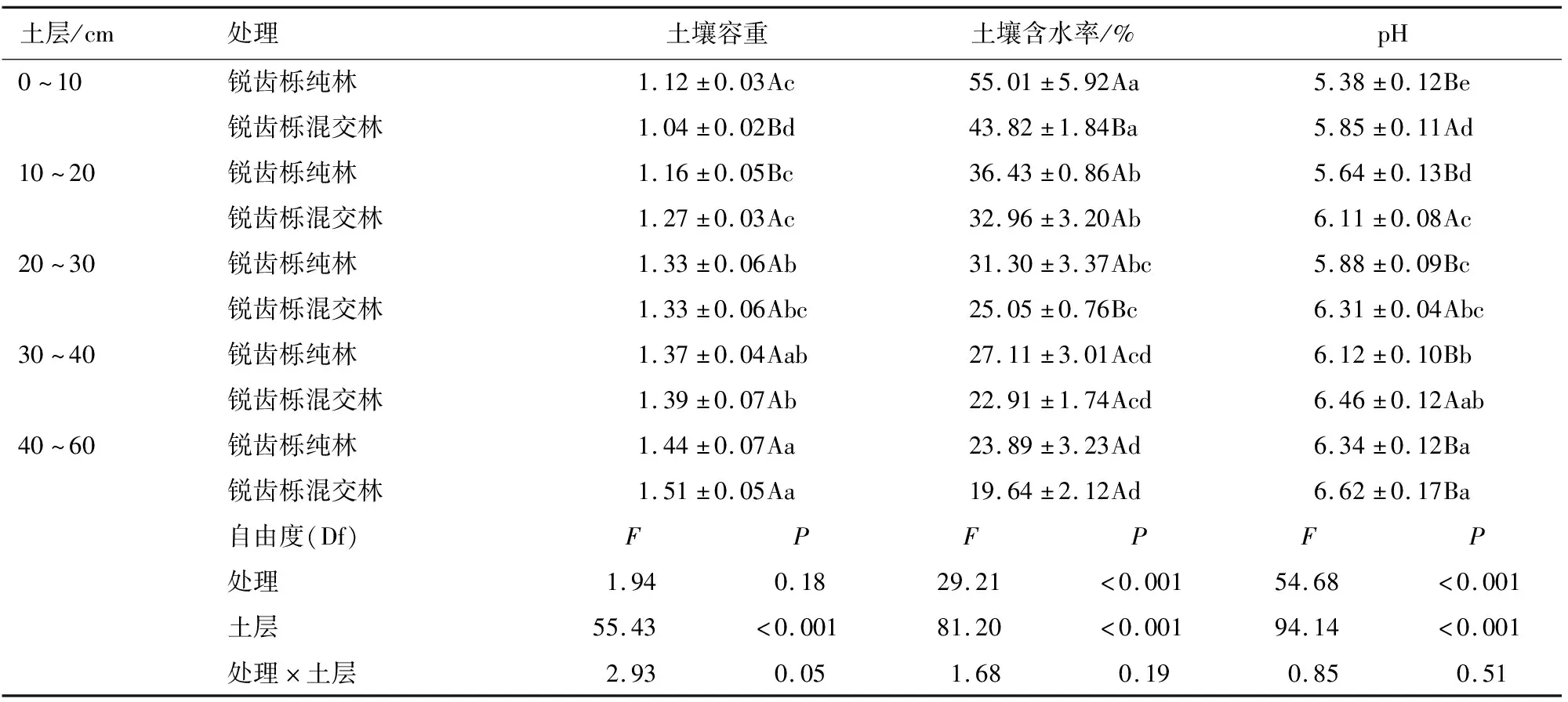
表4 不同植被类型下土壤理化性质的双向ANOVA分析
3.1.2 不同植被类型对土壤酶活性的影响 CBH能够促进纤维素的降解,BG能够将纤维二糖水解成葡萄糖[23]。NAG和LAP是与N循环相关的关键酶。AP能将有机P水解为可利用的无机P,其活性与土壤P供应潜力和P有效性直接相关[9]。不同植被类型和土层深度都会影响与C、N、P循环相关酶的活性(表3)。研究表明,不同森林植被类型土壤酶活性存在显著差异,且不同土层的土壤酶活性也有一定差异性[24]。植被在不同土层有着不同的养分需求,不同土层的土壤含水率都会影响土壤酶活性。本研究中,锐齿栎纯林与锐齿栎混交林处理在0~10 cm土壤含水量是0~50 cm土层含水量的2.3倍和2.2倍,良好的水分加速了土壤有机P的矿化,并促进了植被的生长,因此锐齿栎纯林土壤微生物释放出更多的磷酸酶分解养分来满足微生物的代谢(表3、表4)。本研究表明,锐齿栎纯林与锐齿栎混交林的酶活性随土层深度增加不断下降,锐齿栎纯林与锐齿栎混交林0~10 cm土层的BG+CBH与NAG+LAP酶活性是40~60 cm土层的3倍,AP酶活性是40~60 cm土层的2倍,研究结果与宋霄君等[25]的研究相似。
3.1.3 不同植被类型对微生物体内C、N和P有效性的影响 本研究中,土壤微生物的代谢受到土壤有效P的限制(所有数据点的角度均>45°,图1A)。同样,在其他研究中确定的微生物P限制归因于酸性土壤中Fe和Al离子的强结合。水溶性P在酸性土壤中慢慢转化为稳定的闭蓄态P,降低了P的利用率[26-27]。
本研究发现,锐齿栎纯林的土壤微生物P限制显著高于锐齿栎混交林(图1C)。锐齿栎纯林的有效P含量显著低于锐齿栎混交林(表2)。此外,锐齿栎纯林的土壤含水量与容重高于锐齿栎混交林(表4),他们加快了土壤有机C的分解,导致了锐齿栎纯林处理更强烈的微生物P限制。为了应对土壤中P素的不足,与N循环相关酶活性相比,微生物产生了更多的与P循环相关的酸性磷酸酶[28]。除土壤含水量和土壤容重外,土壤有机质与全N也是影响P限制的关键因素(图3A、图3C),但是土壤全P与矢量角之间没有显著的相关性(图3C),基于C∶P和N∶P值与矢量角之间的负相关关系(图3e、3f),认为有机P的矿化速率受到C∶P与N∶P的影响,当C∶P与N∶P大时,出现了微生物与植被竞争P的现象,发生P的生物固定,加重了微生物P限制。这与Ma等[13]的研究结果一致。酸性土壤中,锐齿栎混交林适当提高pH值有助于促进铁、铝形成氧化物沉淀减少它们对P的固定[28],从一定程度上降低了微生物P限制。与锐齿栎纯林相比,锐齿栎混交林下较高的有机C导致较高的有效P含量和较低酸性磷酸酶活性(表2、表3)。
前人研究表明,高C∶(N、P、S)的外加C源年输入量不会导致土壤有机C含量的增加[29]。本研究中,凋落物的年输入量显著增加了土壤有机C含量,以及有机残渣中释放了更多的有效P。由于竞争争夺吸附位点和在Fe、Al、氢氧化物等胶体表面形成保护膜,有机C的增加也降低了土壤对P的吸附潜力[30]。然而,锐齿栎纯林显著增加了土壤水分和土壤容重,但降低了土壤pH(P<0.05,表4),从而增强了微生物的活性,加速了有机C的分解以及增加了P的生物固定[31-32],因此释放更多的有效P用来减缓土壤微生物的P限制(直接作用为-0.73,图4)。经过长年锐齿栎纯林种植后,土壤有机C含量比锐齿栎混交林下的土壤有机C降低0.2倍,从而影响了有效P的释放,并且锐齿栎纯林加速了土壤C的矿化,这不利于森林土壤C的固存。
3.2 结论
本研究结果表明,山地次生林土壤微生物的生长主要受到土壤有效P的限制。锐齿栎纯林与锐齿栎混交林相比,增加了微生物P的限制,这主要是由于锐齿栎纯林下土壤水和容重的升高加速了土壤有机C的分解,最终增加了土壤微生物受到有效P的限制。在锐齿栎混交林模式下,由于较多的有机C输入,降低了土壤中Fe、Al离子对P的吸附,同时凋落物会释放更多的有效P,因此锐齿栎混交林减缓了微生物P限制。
此外,微生物的P限制也受到了土壤容重、pH与化学养分计量比的影响,但有机C是土壤微生物P限制的关键因素。本研究通过将次生林植被土壤的理化性质与微生物的代谢限制联系在一起,从而加深了对土壤养分循环机理的研究。
猜你喜欢
养生阅刊(2021年5期)2021-06-03
共产党员·下(2018年6期)2018-12-05
方圆(2018年23期)2018-01-07
中老年健康(2016年12期)2017-01-18
家庭医药·快乐养生(2015年8期)2015-09-10
