
基于Maxent模型预测水曲柳的潜在适生区
2022-04-02 05:53施晨阳赖文峰文国卫蒋天雨朱晓如吕增伟张国防
西北林学院学报 2022年2期
施晨阳,赖文峰,文国卫,蒋天雨,朱晓如,吕增伟,张国防
(福建农林大学 林学院,福建 福州 350002)
气候是物种分布格局的决定性影响因素,第四纪冰期以来,物种的地理分布格局与气候的反复剧烈波动有着不可分割的联系[1]。人类活动导致全球的植被、气候条件、水热变化、冰架和冰盖体积以及海平面发生了剧烈变化[2],而植物的生长主要受水热、土壤、海拔、坡向以及CO2浓度等因素的影响,这使得研究气候变化对植物适生区分布的影响成为近年来的研究热点之一[3]。自第一次工业革命以来,全球变暖日益加剧,据研究表明,在未来全球持续变暖的大环境下,到2100年全球气温中值或将上升4 ℃[4],生物及生态系统多样性受到极大威胁,尤其加剧了濒危植物的生境和分布范围的变化[5],其灭绝的风险也进一步加大。因此,在全球变暖的大环境下,探索濒危植物对气候环境变化的反应,提出合理的保护措施具有重大的意义。
随着对生物分布格局受气候变化影响的深入研究,物种分布模型(species distribution models,SDMs)[6]成为研究适生区变化的便利工具,同时也是生物多样性研究和保护的重要工具[7],其利用现存的物种分布数据和环境变量,并依据特定的算法评估出自然界中物种的生态位并进行投影[8],可以应用于多种学科。尽管目前国内外学者已经开发出了多种预测工具,但大都预测精度较低[9],最大熵模型(Maxent)[10]是基于现存的物种分布记录和环境数据构建的物种分布模型,在样本量极少的情况下就能很好地预测,具有精确度高[11]、样本量需求小、稳定性好[12]等优点,是最受欢迎的模型之一[13]。自2004年问世以来,已被广泛用于预测珍稀植物的适生区变化[14]。利用Maxent分析濒危植物适生区的变化,可对其今后的保护提供科学依据。
水曲柳(Fraxinusmandschurica)属木犀科(Oleaceae)梣属(Fraxinus),已被列为我国二级濒危保护植物[15]。其主要生长在海拔200~2 100 m的山坡疏林中或河谷平缓山地的下坡位[16],是红松(Pinuskoraiensis)等树木的优良伴生种。造成水曲柳濒危的原因主要有两方面:首先其对生境要求严苛,扩散能力差[17];其次因为其材质致密坚韧,是不可多得的工艺良材。近年来随着对水曲柳开发利用强度的加大,乱砍滥伐现象严重,以至于天然林面积骤减。因此水曲柳作为重要的经济树种和用材树种,制定科学的培育措施对其人工林的引种开发、种质资源的利用与保护、促进林业可持续发展具有重要意义。遗憾的是目前水曲柳适生区预测的相关报道较少,多集中于木材组分[18]、木材性质[19]、基因表达[20]、森林抚育[21]等方面,有关水曲柳潜在适生区在过去和未来气候变化背景下如何变化,制约水曲柳地理分布的主要因子有哪些,水曲柳未来的高等适生区在哪,这些问题尚未解决,因此制约着水曲柳的保护和开发工作。
本研究利用Maxent模型,模拟推测出水曲柳现代的适生区分布格局,进一步推测出其在末次间冰期(last interglacial,LIG,距今约12万~14万a)、末次盛冰期(last glacial maximum,LGM,距今约2.2万a)、全新世中期(mid-holocene,MH,距今约6 000 a)以及2050s、2070s的潜在分布区,同时利用ArcGIS10.4.1软件进行适生分布区的划分,根据百分比贡献率、置换重要值、jackknife检验和环境限制因子探讨影响水曲柳潜在分布区形成的重要环境因子,为水曲柳种质资源保护和管理开发提供基础数据,同时为人工林的引种栽培规划提供参考。
1 材料与方法
1.1 研究区域
水曲柳野生群居跨越中国东北部,中国西北部分地区,俄罗斯东部,日本北部及朝鲜和韩国,呈间断散生分布,生境破碎化程度较高,水曲柳在我国的天然分布区经纬度跨度为28°-52°N,102°-134°E,最北达黑龙江大兴安岭山区的黑龙江盘中国家级自然保护区(52°46′N,123°52′E),南至四川省屏山县北部山区(28°33′N,104°12′E),最西至四川省康定市东北部低山丘陵(30°7′N,102°6′E),东界为黑龙江省佳木斯市抚远市和同江市境内的三江国家级自然保护区(48°16′N,134°20′E)。
1.2 水曲柳样本搜集与筛选
2019-2021年课题组对宁夏、陕西和甘肃3省的水曲柳天然种群进行了实地调查,搜集分布记录84条;并通过检索中国国家标本资源平台(NSII,http://www.nsii.org.cn)、教学标本资源共享平台(http://mnh.scu.edu.cn/main.aspx)、全球物种多样性信息库(http://www.gbif.org/)等资源共享平台,并检索已出版的文献资料,分别获得水曲柳分布记录137、113、108、58条,同时查阅地方植物志等网站获得15条,共计434条水曲柳分布记录,参照李垚等[22]的方法对水曲柳的全部分布记录进行筛选,删除人工引种栽培记录、采样点模糊无具体位置记录、重复多余等记录,从而降低样本误差,每个网格(10 km×10 km)只保留1个分布点的原则,进行缓冲区分析,最终获得有效的水曲柳天然分布点88个(图1)。
1.3 环境因子数据的筛选
参与本研究气候因子和海拔因子来自于WorldClim数据库(http://worldclim.org),土壤因子来源于国家青藏高原科学数据中心(http://westdc.westgis.ac.cn)的“基于世界土壤数据库(HWSD)(http://www.fao.org/faostat/en/#data.)的中国土壤数据集(V1.1)(2009)”,将土壤因子和气候因子的空间分辨率统一为30″(30 arc-second,约1 km2),按照中国1∶100万中国地图矢量图剪裁为一致大小,使用ArcGIS10.4.1软件对所有的因子进行掩膜、重采样等过程,生成与当前气候像元大小一致的栅格数据图层[23],并保存为asc格式。将所有的因子进行点插值提取并将数据整理在表格中,用SPSS软件对整理的数据进行相关性分析,根据相关性分析数据并结合水曲柳的生长习性和生境特点,筛选出对水曲柳的潜在适生区影响最大的环境因子。
在本研究使用EMNeval数据包优化的Maxent模型[24],利用其中的RM(regularization multiplier)和FC(feature combination multiplier)2个参数,将RM的数值设置为0.5~5,每次增加0.5,总共10种调控倍频。Maxent模型提供了5种特征,分别是线性(linear,L)、二次型(quadratic,Q)、片段化(hinge,H)、乘积型(product,P)和阈值性(threshold,T)。采用FC的8种特征组合:即L、LQ、H、LQH、LQHP、LQHPT、HPT、QPT,ENMeval数据包将上述 80种参数组合进行测试,最终采用Akaike信息量准则的delta AICc模型,并采用OR10和AUCDIFF检验模型的拟合度与复杂度。优化完成后,采用优化参数进行模拟和预测水曲柳不同时期适生区,将正则化乘数设置为3.5,背景点最大数量设置为10 000,最大迭代次数设置为10。利用受试者曲线下的面积(AUC)评估Maxent预测的准确性,AUC值的取值范围为0~1,值越大表示适生区和不适生区区分可信度越高,通常AUC值小于0.7被认为是低精度,0.7~0.9为是中等精度,而大于0.9则是高精度[25]。
1.5 适生区划分
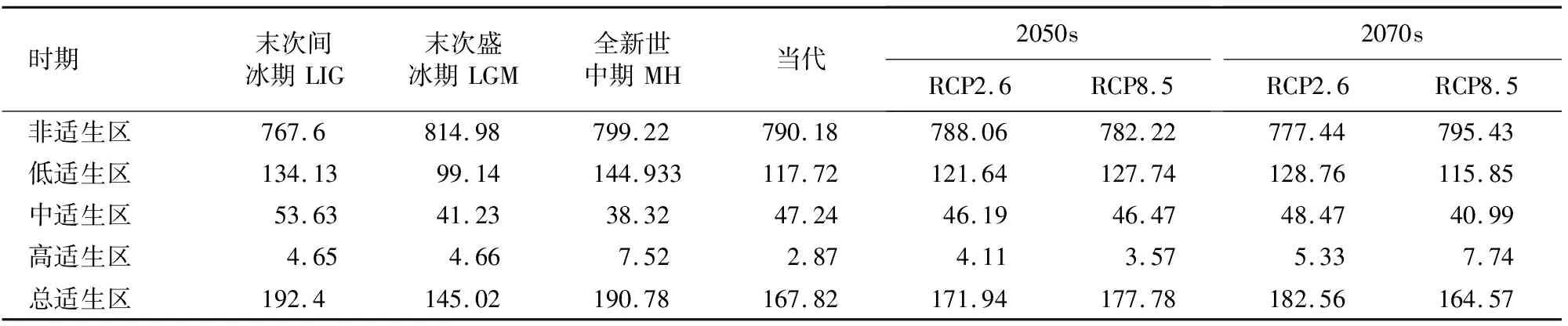
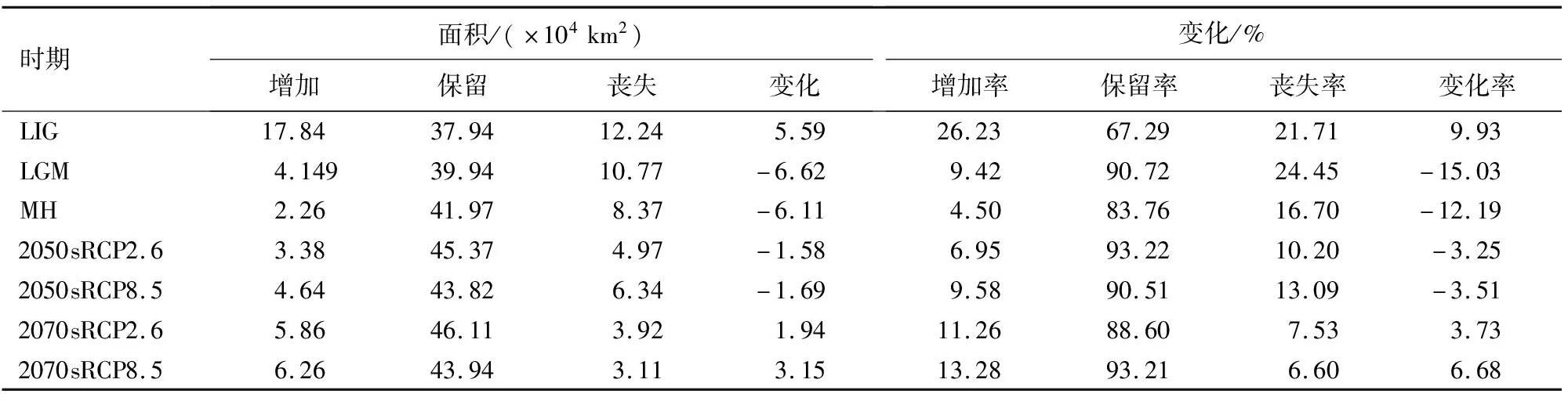
用ArcGIS10.4.1软件对Maxent的模拟数据进行处理,将模拟10次的平均值导入其中,并转化为Raster栅格数据,栅格的数值表示水曲柳的生存概率,对概率P进行重分类,将水曲柳的预测分布区划分为4类,分别为非适生区(P<0.2)、一般适生区(0.2 参照张殷波等[26]的方法,将50%设置为阈值,水曲柳的分布概率Q<50%设置为非适生区,其赋值为0,Q≥50%作为适生区,赋值为1,以此建立水曲柳在过去、当前和未来气候变化情景下潜在地理分布的(0,1)矩阵,将矩阵值0→0定义为非适生区,0→1为新增适生区,1→0为丧失适生区,1→1为保留适生区。最后在ArcGIS10.4.1中加载矩阵变化值,基于水曲柳当前适生区面积计算过去和未来不同情景下的面积,再基于过去、当前气候与未来气候变化情景下的矩阵变化值。 基于88个天然记录和15个环境因子,利用Maxent模型对水曲柳的潜在适生区进行预测,当最大迭代次数为10时,得出当代AUC最大值为0.952 4,最小值为0.806 3,平均值为0.874 0(±0.004 5,标准偏差),表示此次预测结果极准确。 表2表明,贡献率(percent contribution,PC)排在前6位的环境因子为年降水量(bio12,35.4%)、最暖季度平均气温(bio10,15.39%)、等温性(bio3,11.58%)、降水量季节性变化(bio15,7.48%)、海拔(elev,7.46%)和最干月降水量(bio14,7.15%),累计值为84.46%;置换重要值(permutation importance,PI)位居前6环境因子的为年降水量(bio12,29.28%)、海拔(elev,22.05%)、年平均气温(bio1,10.1%)、降水量季节性变化(bio15,8.56%)、最干月降水量(bio14,8.11%)、最暖季度降水量(bio18,6.6%),累计值为84.7%;刀切法分析结果显示(图2),仅使用单独变量模拟时对正规化训练增益影响最大的环境因子为年降水量。综合上述结果,本研究认为降水因子(年降水量、降水量季节性变化、最干月降水量、最暖季度降水量)是制约水曲柳现代适生区分布的主要环境因子。除此以外,温度因子(最暖季度平均气温、等温性、年平均气温)和海拔因子也是制约水曲柳分布的重要环境因子。 表1 水曲柳主要环境因子参数 从表2可知,当前时期水曲柳的潜在适生区面积为167.82×104km2,其中一般适生区面积为117.72×104km2、中等适生区的面积为47.24×104km2、高度适生区面积为2.87×104km2。水曲柳的潜在适生区主要分布在长白山、千山、大小兴安岭、六盘山和秦岭等山脉。由图3可知,水曲柳在LIG、LGM、MH的西北的潜在适生区逐渐减少,其在东北的潜在适生区逐渐增加。而20世纪50年代(RCP2.6、RCP8.5)的情境下,水曲柳在西北地区分布逐渐减少,东北地区的潜在适生区与现代相当。20世纪70年代(RCP2.6、RCP8.5)的情境下,东北的适生区面积增加较明显且出现南移趋势。 表2 不同时期水曲柳适生区面积变化 由不同气候情景下水曲柳适生区空间变化(表3)和不同时期水曲柳适生区空间变换格局(图4)可以得出,过去3个时期,LIG时期的增加率最高,达26.23%,贵州西部、甘肃和宁夏南部、内蒙古东北部和黑龙江中北部增加区有较大面积扩张;LGM时期的丧失率最高,达24.45%,丧失区面积为10.77×104km2。在未来4种气候情景下,水曲柳的保留率较高,分别为93.22%、90.51%、88.60%和93.21%,山东、河北、辽宁等地出现零星的增加区,其中水曲柳在21纪70年代的2种气候情景下增加率均大于丧失率,说明水曲柳在21世纪70年代具有生长优势。 表3 不同气候情景下水曲柳适生区空间变化 在LIG、LGM、MH、2050sRCP2.6、2050sRCP8.5、2070sRCP2.6、2070sRCP8.5气候情景下,水曲柳88个现代分布点的平均多元相似度分别为-1.48、7.19、12.5、19.17、15.96、16.08、13.31,多元相似度为负值的点比例分别为27.85%、11.84%、0、0、1.25%、0、2.53%,由此表明,LIG时期气候异常程度最高,LGM时期次之,其他时期都比较低。在末次间冰期,现代适宜区内的主要气候异常区域分布在西部和南部,最不相似变量为bio15和bio10。在LGM时期,现代适宜区内的主要气候异常区出现在北部,最不相似变量为bio1和bio6。在全新世中期,现代适宜区内的主要气候异常区仅零星分布,最不相似变量为bio3(图5)。未来种气候情景下,现代适宜区内的主要气候异常区均主要出现在东南部,最不相似变量为bio3、bio8、bio10、bio18(图6)。 通过搜集水曲柳的天然分布点,利用Maxent模型分析影响水曲柳分布的主要环境因子,模拟各时期水曲柳的潜在适生区。研究结果表明,年降水量、最暖季度平均气温、等温性、降水量季节性变化、海拔和最干月降水量是影响水曲柳分布的主要环境因子。在当前气候情景下,水曲柳的高等、中等和低等适生区面积分别为2.87、47.24、117.72×104km2,高等适生区主要集中在我国东北和西北,但在2地的变化趋势不同,西北适生区破碎化严重且呈收缩趋势,东北则呈现纬度上的上下波动;水曲柳的高等适生区面积在过去和未来几个时期较当代均有增加,推测过去此地区可能存在水曲柳稳定的冰期避难所,在未来4种气候情境下,随着时间的推移,高等适生区的面积呈扩张趋势,说明水曲柳可能更适合未来的高温高湿环境。本文通过预测水曲柳的潜在适生区和对其产生重要影响的环境因子,对水曲柳种质资源的保护和开发具有重要的参考价值,并为水曲柳潜在种植区的规划和设计提供一定的依据。 本研究中,按照表2中8种主要的评估途径来看,降水因子的重要性列入第1位5次,第2位3次,气温因子的重要性列入第1位1次,第2位2次,表层土砾石含量的重要性列入第1位2次,表层土壤有机碳含量列入第2位2次,海拔仅占第2位1次,影响水曲柳分布的环境因子按其重要程度可以排列为降水因子、气温因子、土壤因子和海拔因子,其中降水和气温因子是制约水曲柳分布最重要的2个因子。 贡献率和重要置换值均显示年降水量居首位,且仅使用单独变量模拟时AUC值影响最大的环境因子均为年降水量(图2)。对比他人的研究结果,Wan等[27]研究同样认为制约水曲柳适生区的最重要环境因子是年降水量。此外水曲柳和蒙古栎(Quercusmongolica)同为红松阔叶林的主要建群树种,其生长环境相似,贾翔等[28]研究表明制约红松的最主要环境因子是年降水量,殷晓洁等[29]研究发现年均降水量是影响蒙古栎适生区分布的最重要变量,与本研究结论相似。 气候变化对物种的空间分布产生重要影响,尤其是第四纪以来环境条件的剧烈变化,生物应对气候变化适生区发生迁移,形成了现代物种的地理分布和遗传结构,但气候因子并非相互独立地影响生物的生长和分布,而是与多种气候因子共同作用的结果。多元环境相似性面和最不相似变量分析表明,LGM时期的气候异常程度中等,多种因素的改变导致现代分布区的适宜度降低,其中北部主要与年均温和最冷月最低气温有关,中部与最暖季度平均气温有关;未来4种气候情景下,水曲柳分布区的东南部气候异常程度较剧烈,此地区最不相似变量主要为最湿季度平均气温和最暖季度平均气温,与此同时,水曲柳在这些地区均出现新增区域,降水因子是制约水曲柳扩张的重要变量,综上所述水曲柳地理分布变迁主要受主导因子的影响并受到其他因子的制约,是多种因子综合作用的结果。 水曲柳的潜在适生区在过去3个时期面积波动明显,这可能是第四纪气候多次循回震荡造成,而东北和西北一直是其高度适生区,本文推测这2地可能为水曲柳的冰期避难所。这与众多学者的研究结果一致,叶俊伟等[30]研究结果显示,长白山和朝鲜半岛是针阔混交林植物最重要的2个避难所,且秦岭也是众多动植物重要的冰期避难所[27]。 在未来4种气候情景下,水曲柳的高等适生区面积以及分布略有差异,但总体相比现代高等适生区均呈增加趋势,说明水曲柳较为适应未来的高温环境,文冠果(Xanthocerassorbifolia)[31]也有类似的趋势。未来时期水曲柳的潜在适生区在西北局部和东北地区保留较好,丧失区主要出现在甘肃、陕西、山西,增加区在河北、山东、辽宁等地零星出现,但高等适生区面积较现代明显增多,故本研究推测未来气候环境可能更加适应水曲柳的生长发育。 水曲柳应对未来气候变化的响应表现为高度适生区不同程度的扩张和空间格局的显著变化。水曲柳虽被列为国家重点保护树种,但仍然受人类采伐的威胁较大[17],在其未来的保护过程中,应不仅关注保留区的变化,对丧失区和保留区的变化也要给与足够的重视,制定对应的保护措施。 保留区在温室气体不同浓度梯度条件下,可能成为水曲柳应对气候变化的安全避难所,重要地位由此可见一斑,因而更应该注重对此区域的保护与管理。目前水曲柳集中分布在长白山、千山、大小兴安岭、六盘山、秦岭等山脉,但多地重视程度不足,建议未来在此地区建立或扩大自然保护区范围。 对于丧失区,应分析水曲柳趋向丧失的原因,建立立地保护模式。在未来气候条件下,甘肃、陕西、山西等地区出现了不同程度的丧失区,推测该地区的气候条件可能不再适宜水曲柳的生长和发育,建议加大经费投入,积极开展水曲柳繁殖生物学、群体生态学和人工驯化等方面研究,避免遗传适应性变异及遗传资源丧失等风险。同时应减杜绝乱砍滥伐,因地制宜地选择营造水曲柳人工群落,通过迁地移栽大苗、种子育苗和无性繁殖等方式对其进行培育。 新增区可作为水曲柳的引种区,应结合其生理习性,选择合理的位置并制定合理的土地规划利用方案,逐步建立开发引种示范区和试点,减少人类活动对水曲柳的影响,制定有效的监管措施并严格实施,因水曲柳的自生扩散能力较差,需要人工辅助迁移来帮助其扩散并定殖。1.6 物种适生区空间格局变化
2 结果与分析
2.1 模型优化结果
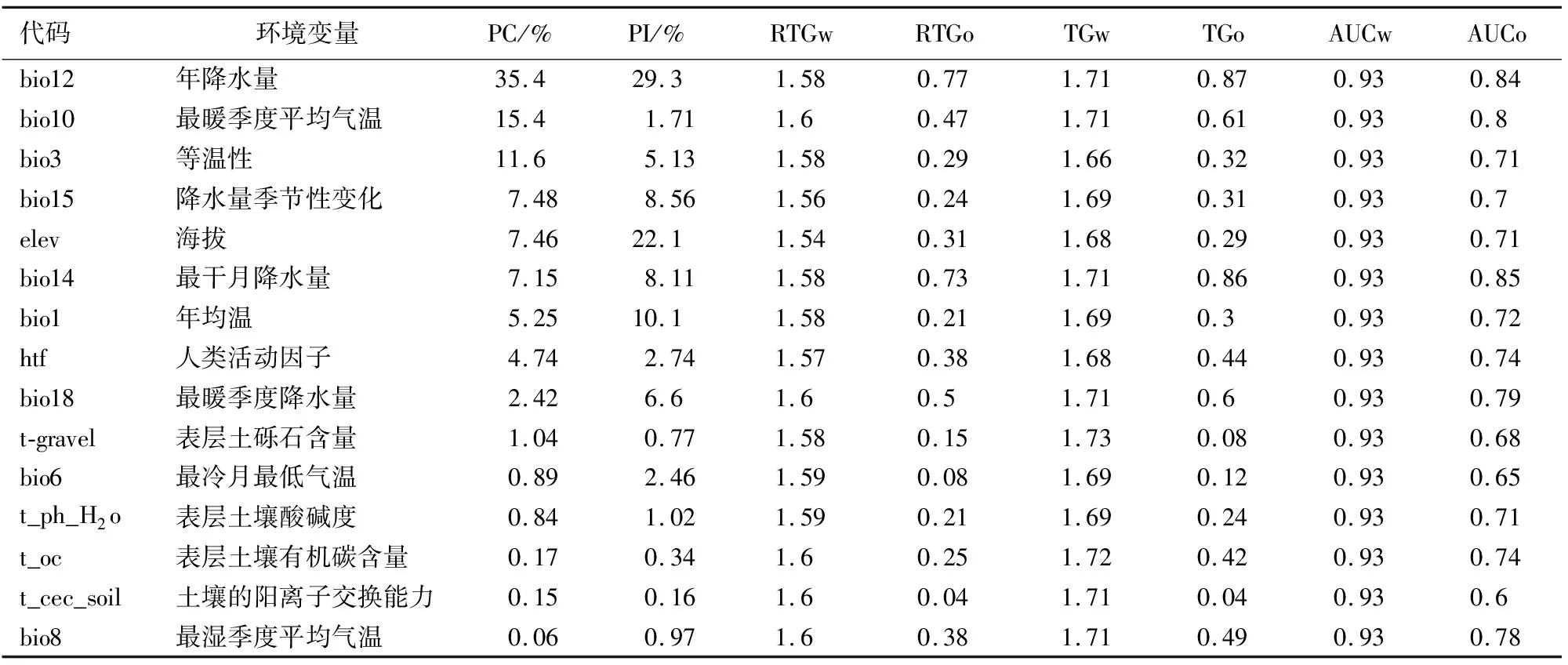
2.2 影响水曲柳分布的环境变量
2.3 水曲柳潜在适生区面积变化和空间格局变化
2.4 多元环境相似度面和最不相似变量分析
3 结论与讨论
3.1 气候因子对水曲柳地理分布的影响
3.2 水曲柳潜在潜在适生区的地理变迁
3.3 水曲柳资源的保护与开发
猜你喜欢
西北林学院学报(2022年5期)2022-10-04
西北林学院学报(2022年4期)2022-08-02
成都信息工程大学学报(2022年3期)2022-07-21
中国中医药信息杂志(2022年7期)2022-07-17
云南农业大学学报(自然科学)(2022年2期)2022-04-07
福建农林大学学报(自然科学版)(2022年1期)2022-01-18
现代农业科技(2019年22期)2019-12-25
南方农业·下旬(2019年4期)2019-08-26
华人时刊(2018年17期)2018-11-19
现代农业科技(2017年16期)2017-09-22
