
汶川震后滑坡体植被恢复不同阶段优势种种间关联研究
——以银厂沟谢家店滑坡体为例
2022-04-02 06:10刘鲁光朱兆棋刘守江
西北林学院学报 2022年2期
刘鲁光,朱兆棋,陈 曦,刘守江
(1.西华师范大学 生命科学学院,四川 南充 637009; 2.西华师范大学 国土资源学院,四川 南充 637009; 3.四川省干旱河谷土壤侵蚀监测与控制工程实验室,四川 南充 637009)
种间联结是指研究区域内不同物种在空间分布上的关联情况,是群落重要的数量和结构特征之一[1]。植物群落中的种间关系由于生物和环境因子的综合作用往往十分复杂,物种间的正联结代表物种为相互依赖的关系或有着相似的环境需求,物种间的负联结代表物种为竞争关系有着不同的环境需求,无联结则往往说明物种间相互独立[2]。研究物种间的联结状况对揭示群落演替过程中物种间的相互作用和群落动态具有重要意义[3]。
近年来,多位学者对国内不同区域植被的种间关联动态进行了研究。在鼎湖山落叶阔叶林群落的种间联结动态的研究中发现,群落整体呈正联结但正联结程度在逐年下降[4];在海南热带雨林群落联结动态的研究中发现,群落整体在向无联结的方向发展[5];在对退化生态系统植被恢复过程中种间关联动态的研究中,多位学者发现随着恢复时间的增加群落的正联结程度逐渐加强[6-8]。
对植物群落的研究在灾后水土保持和生态建设工作中发挥着极其重要的作用,目前对该区域植被恢复状况的研究主要集中在群落的物种组成及多样性变化[9-11]、植被状况与土壤理化性质的耦合关系[11,13-15]、植被生物量[10]、群落稳定性[16]等方面,对种间联结的研究还只有基于1次调查的短期研究[12],还未有人对该区域植被恢复过程中种间关系的动态变化进行研究。本研究根据调查的实际状况并结合前人对植被恢复阶段的划定[14-15],选取草本群落(2009年)、灌草群落段(2012年)、乔灌草群落(2015、2018年)3个恢复阶段,对谢家店滑坡体不同恢复阶段的群落整体关联性的变化和群落垂直层次内种间关联动态进行了研究,可为谢家店滑坡体后续的植被恢复和生物多样性保护提供依据。
1 材料与方法
1.1 研究区概况
谢家店滑坡体位于四川成都彭州市北部山区的龙门山镇,坐标介于30°54′-31°26′N,103°10′-103°40′E。滑坡体形成于2018年5.12汶川大地震后,位于龙门山断裂带上,构造运动活跃[17]。平均海拔1 265 m,土壤类型主要为山地灰棕壤。该区域属亚热带湿润气候区,全年气候温和,雨热同期。年均气温16 ℃,7月最热,最热月平均气温25 ℃,1月最冷,最冷月平均气温5 ℃;年均降水量867 mm,年内降水量的时空分配不均匀,夏秋季节降水多,冬春季降水少;该区域年均日照时数1 131 h,无霜期较长。谢家店滑坡体经过十余年的近自然恢复,群落已由次生裸地演替为垂直分层明显的乔灌草群落,2018年调查时滑坡体乔木层常见树种有桤木(Alnuscremastogyne)、柳杉(Cryptomeriafortunei)、盐肤木(Rhuschinensis),灌木层以蜡莲绣球(Hydrangeastrigosa)、灰白毛莓(Rubustephrodes)、川莓(Rubussetchuenensis)为主,草本层物种以木贼(Equisetumhyemale)、野棉花(Anemonevitifolia)、芒萁(Dicranopterispedata)、糯米团(Gonostegiahirta)为主。
1.2 数据来源
为了解谢家店滑坡体植被恢复过程中的发展规律,在2008年地震发生后,对谢家店滑坡体植被恢复状况进行了长期的动态观测。根据研究区域的实际状况,从滑坡体下方出发,以20 m的海拔梯度自下而上共设置9个固定样带。每个样带中设置1个10 m×10 m乔木样方,每个乔木样方中设置2个5 m×5 m的灌木样方,每个灌木样方中设置4个1 m×1 m的草本样方,记录样方中植物的种类、数量、盖度和频度,同时记录样方所在地点的海拔、坡度、坡向和经纬度等位置信息,样地基本情况如表1所示。本研究利用2009、2012、2015年和2018年的4次调查数据对群落恢复不同阶段的种间关联进行分析。
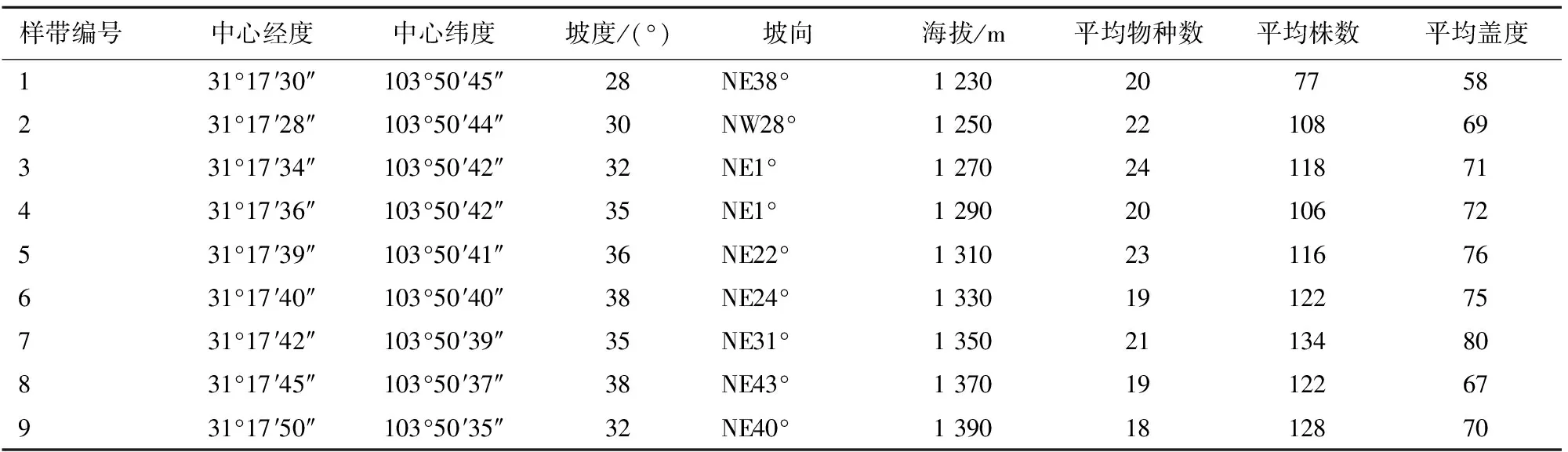
表1 样地基本情况
1.3 数据处理
分别计算各年份不同层次物种的重要值,并将各层次物种按重要值从大到小排序,选取各年份乔木层、灌木层、草本层各层次中重要值之和大于80%的物种作为当年的优势物种(表2)进行种间联结研究。数据计算处理使用Microsoft Excel 2016、Spss26.0软件完成,物种间关联矩阵利用R corrplot进行绘制[18]。

表2 谢家店滑坡体主要物种名及代码
1.3.1 重要值计算 重要值(IV)可表示物种在群落中的重要程度,本研究采用相对密度、相对频度、相对盖度3个指标进行物种重要值计算,计算公式[1]如下:
IV=(相对密度+为相对频度+相对盖度)/3
(1)
式中:相对密度为某个物种的个体数与所有物种个体数之比;相对频度为某物种的频度与所有物种频度和的比;相对盖度为某物种的总盖度与所有的物种盖度和之比。
1.3.2 多物种间的总体关联性检验 群落多物种间整体关联性采用Schluter提出的方差比率法(VR)来测定,并用统计量W来检验物种间联结程度,计算公式[19]如下:
(2)
Pi=ni/N
(3)
(4)
(5)
W=N×VR
(6)
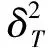
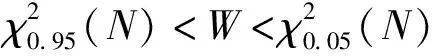
1.3.3 成对物种间联结性检验 成对物种的种间关联性利用物种的多度数据进行计算,采用spearman秩相关分析来进行分析,计算公式[19]如下:
(7)
式中:N为总的样带数;xij,xkj分别为物种i和k在样方j中的秩。
2 结果与分析
2.1 群落整体关联性的变化
群落优势物种间的种间联结动态可以反映出群落物种间关联性的总体变化趋势[20]。从物种间整体关联的结果(表3)可以得出,植被恢复过程中的草本阶段与乔灌草阶段方差比率均小于1,VR值未通过检验,群落整体呈不显著的正联结;在灌草阶段,方差比率小于1,VR值未通过检验,群落整体呈不显著的负联结。方差比率随恢复年限的增加整体呈上升趋势,但关联程度仍未达到显著水平。这说明恢复过程中群落物种间正联结程度不断加强,但种间关系仍较分散。
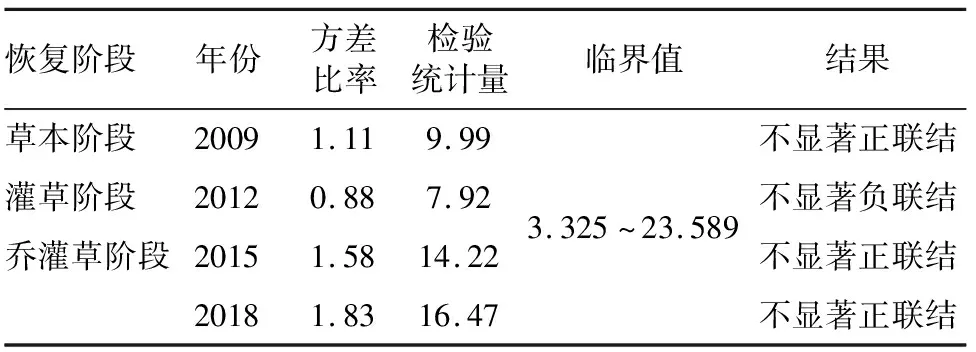
表3 物种间整体关联性检验
2.2 群落主要物种种间联结特征及其变化
2.2.1 草本层主要物种种间关系 草本层Spearman秩相关检验的统计结果(表4)表明,在恢复过程中,草本层正联结种对比例呈先下降后上升的趋势,由草本阶段的54.5%下降为灌草阶段的32.1%,在2018年乔灌草阶段上升到86.7%,负联结种对比例呈先上升后下降的趋势由草本阶段的45.5%上升到灌草阶段的64.1%,2018年乔灌草阶段下降到12.4%。草本层物种正负联结比例变化趋势与群落物种间整体关联状况变化一致。
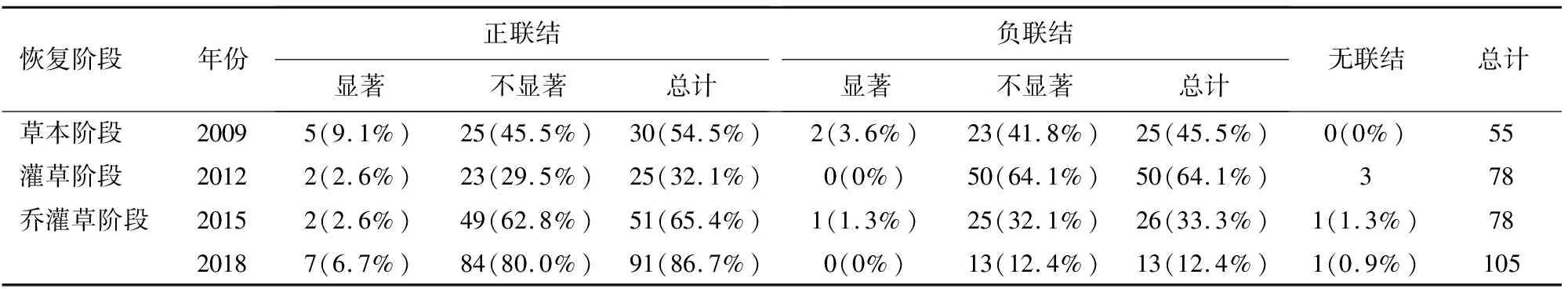
表4 草本层spearman秩相关统计结果
各恢复阶段中,草本层达到显著相关的种对所占比例较少。2009年草本阶段(图1)为艾-清明草、莎草-繁缕、莎草-画眉草、葎草-冷水花呈显著正相关,水蓼-冷水花、水蓼-葎草呈显著负相关;2012年灌草阶段(图2)为鸭儿芹-苔草、醉鱼草-隐子草呈显著正相关;2015年乔灌草阶段(图3)为鱼腥草-伏地卷柏、猪殃殃-知风草呈显著正相关,猪殃殃-杭子梢呈显著负相关;2018年乔灌草阶段(图4)为蜂斗菜-老鹤草、蜂斗菜-糯米团、蜂斗菜-野棉花、金粉蕨-木贼、老鹤草-糯米团、老鹤草-水蓼、糯米团-野棉花呈显著正相关。乔灌草阶段以前,呈显著正相关的物种均为生态习性相似的物种,草本阶段出现的水蓼的适生幅度特别宽,干旱和湿润的环境均能正常生长,与物种间竞争激烈,因此与多个物种呈显著的负相关。进入乔灌草阶段后呈显著相关的多为喜湿耐阴的物种,部分盖度较大的阳生草本也为喜阴湿环境的物种提供了适合的生存条件,如野棉花与糯米团,金粉蕨与木贼,物种间对资源的利用更加高效合理。
2.2.2 灌木层主要物种种间关系 灌木层物种间正联结种对比例随恢复时间增加呈现下降趋势,负联结种对比例随恢复时间增加呈上升趋势。在灌草阶段,灌木层物种间的正负联结种对之比大于1,2015年群落演替到乔灌草阶段时,灌木层主要物种间正负联结种对之比较草本阶段明显下降(1.50→0.91),但乔灌草阶段的2018年与2015年相比变化不明显(0.91→0.90)。
2012年到2018年的恢复过程中,灌木层大多数物种间均未达到显著的正负联结。2014年(图5)灌草阶段为灰白毛莓-柳杉幼苗呈显著负联结,毛葡萄-紫麻呈显著正联结,2015年(图6)乔灌草阶段主要物种间未形成显著联结种对,2018年(图7)乔灌草阶段为楝叶吴萸-川莓,楝叶吴萸-高山柳,高山柳-金丝梅,领春木-地石榴呈显著正相关,楝叶吴萸-地石榴,楝叶吴萸-领春木,楝叶吴萸-溲疏,领春木-高山柳,领春木-蜡莲绣球,蜡莲绣球-木莓呈显著负相关。
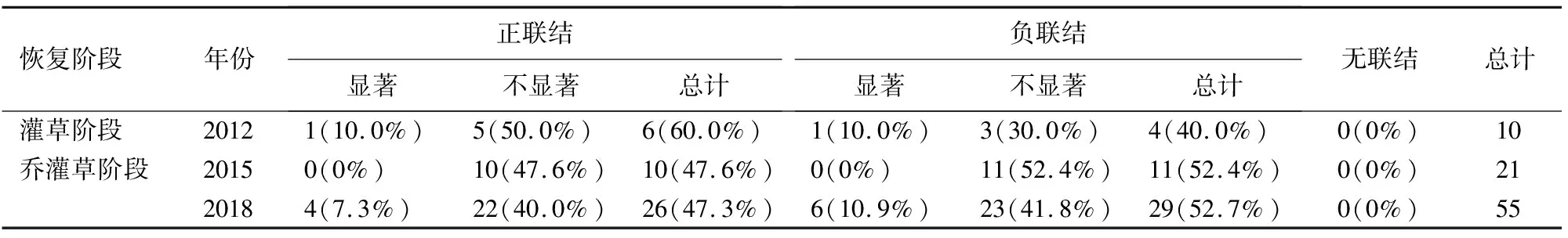
表5 灌木层spearman秩相关统计结果
2015年灌草阶段灌木层未有物种达到显著相关,较分散的种间关系在一定程度上降低了灌木层物种的竞争强度[21]。2018年乔灌草阶段,呈显著相关的物种多为喜光、抗逆性强的物种,如高山柳、金丝梅,同时较之前的恢复阶段出现对耐阴、多生长于林冠下层的领春木、地石榴等,灌木层的楝叶吴萸生长迅速,种子可依靠风力快速传播[22],在调查中各个样地均有分布,因此与多个物种间呈现显著相关的关系。
2.2.3 乔木层主要物种种间关系 乔木层spearman秩相关统计结果表明(表6),乔木层正负联结种对比由2015年的0.5上升到2018年的1.5。乔木层达到显著相关的种对比例较低,2015年(图8)为盐肤木-柳杉,盐肤木-胡桃呈显著正相关,2018年(图9)为盐肤木-木姜子呈显著正相关,柳杉-胡桃呈显著负相关。
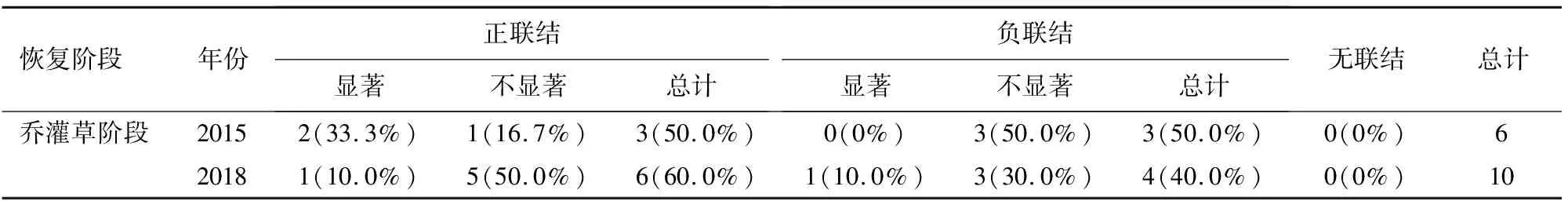
表6 乔木层spearman秩相关统计结果
3 结论与讨论
3.1 结论
物种间的整体关联性除2012年灌草阶段呈不显著的负联结外,其他各恢复阶均为不显著的正联结,且正联结程度随恢复时间的增加整体呈上升趋势,但还未达到显著程度,这说明群落还处于恢复阶段,在向稳定方向演替。
草本层在灌草阶段,负联结种对比例超过正联结,物种间竞争激烈,其他各恢复阶段均为正联结种对比例大于负联结,且正联结种对比例随恢复年限增加整体呈上升趋势,负联结种对比例整体呈下降趋势。
灌木层正负联结种对之比随恢复年限的增加呈下降趋势(1.50→0.91→0.90),但这一下降趋势趋于缓和。说明灌木层物种还处于演替的不稳定阶段。
乔木层物种随恢复年限的增加物种间正联结种对比例呈上升趋势,负联结比例呈下降趋势,与群落整体关联情况的变化趋势一致。
3.2 讨论
一般来说,随着演替的进行,物种间整体关联会逐步趋于正相关,以求得多物种共存,群落结构与物种组成也会逐渐趋于完善和稳定[3],谢家店植被在恢复过程中除2012年灌草阶段方差比率小于1,物种间整体呈不显著的负联结外,其他恢复阶段方差比率均大于1,物种间整体呈不显著的正联结,且正联结程度随恢复时间的增加呈增强趋势,谢家店群落种间关系的变化趋势符合这一规律,并与前人研究时的预测相符[12]。群落正联结程度的不断加强也说明植被恢复过程中群落的演替是正向的,物种逐渐与区域环境相适应,群落结构也在不断完善,物种间对资源和空间的利用更加合理。
从整个恢复过程来看,群落整体在向稳定方向发展。演替初期,环境恶劣,空间与资源的分布状况是影响这一阶段影响物种分布的主要原因[23-24],对环境适应性强,繁殖速度快的先锋草本物种快速出现,如艾、清明草、醉鱼草等,由于资源空间有限,这些对生境要求相似的物种聚集在一起,多数呈集群分布,由此导致这一阶段群落物种间的整体关联为不显著的正联结,这与王加国等[25]的研究结果相一致。演替的灌草阶段,环境还未明显改善,灌木层物种的出现使群落物种间对有限资源的竞争更加激烈,加之灌木出现后占据了大部分的上层空间,缩减了原有喜光植物可利用的光照资源和生存空间,加强了物种间的竞争关系,因此这一阶段群落整体呈现不显著的负联结。演替进入乔灌草阶段,群落在垂直层次上的分化更加复杂,草本层部分喜光耐旱的优势物种在灌木阶段已逐渐被喜湿耐阴的物种替代,如草本阶段的清明草、醉鱼草等阳生优势种已演替为木贼、伏地卷柏等喜阴湿的物种,因此乔木的出现对草本层多数物种更有利,在很大程度上缓解了草本层物种间的竞争关系,使得草本层物种的正负联结种对之比在乔木层出现后有明显的提升(1.96→7)。同时,乔木的出现加剧了灌木层物种间的竞争,灌木层物种的正负联结种对之比较灌草阶段下降明显,但下降趋势逐渐缓和(1.50→0.91→0.90),这是由于灌木层中山梅花、毛葡萄等喜光植物逐渐被川莓、蜡莲绣球、楝叶吴萸等喜湿耐阴物种所替代。总的来说,乔木的出现使物种在垂直层次上的分化更加合理,使物种对资源空间的利用更加充分,因此使物种正关联程度呈加强趋势。
猜你喜欢
草业科学(2022年6期)2022-06-29
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
中学生理科应试(2021年10期)2021-12-07
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
中国新技术新产品(2018年13期)2018-09-21
地球(2015年3期)2015-03-26
绿色科技(2014年4期)2015-01-06
小说月刊(2014年1期)2014-04-23
