
肝特异性LCMT1基因敲除小鼠模型的制备及鉴定
2022-03-24 05:51莫娇敖青青郑志坚吴懿洁梁宁静廖思米王新航陆彩玲唐深李习艺
中国实验动物学报 2022年1期
莫娇敖青青郑志坚吴懿洁梁宁静廖思米王新航陆彩玲唐深李习艺*
(1.广西医科大学公共卫生学院营养与食品卫生学系,南宁 530021;2.广西医科大学基础医学院免疫学教研室,南宁 530021)
亮氨酸羧基甲基转移酶1(Leucine carboxyl methyltransferase 1,LCMT1)是一种大小为38×103包含334个氨基酸的蛋白质,其序列高度保守,广泛存在于动物、植物和酵母中[1-3]。LCMT1通过催化蛋白磷酸酶2A催化亚基C(PP2Ac)的羧基端亮氨酸309位点发生甲基化,从而发挥其重要作用[4-5]。蛋白磷酸酶2A(PP2A)是丝氨酸/苏氨酸家族的特异性蛋白磷酸酯酶之一,由结构亚基A,调节亚基B和催化亚基C组成的异源三聚体复合物,通过对下游信号通路去磷酸化发挥重要的作用[6-8]。研究发现LCMT1和蛋白磷酸酶甲基酯酶1(PPME1)共同维持PP2Ac甲基化水平的动态平衡[9-10],干扰甲基化动态平衡导致有丝分裂纺锤体大小异常、细胞生长缓慢、细胞损伤甚至死亡[11-12]。全身敲除LCMT1基因,导致全胚裂解物PP2Ac甲基化水平显著降低,胎儿肝造血功能严重缺陷,胚胎死亡率高达90%[13]。高表达LCMT1促进PP2Ac去甲基化,保护细胞免受H2O2诱导的氧化损伤[14-16]。上述证据表明LCMT1在细胞增殖和胚胎发育过程中是必不可少的。目前关于LCMT1在肝的研究较少,国内外尚无在肝中特异性敲除LCMT1基因的动物模型。本课题通过Cre/Loxp系统特异性敲除肝LCMT1基因,在动物水平上探讨LCMT1基因在肝中的生物学功能。
1 材料与方法
1.1 材料
1.1.1 实验动物
采用CRISPR/Cas9技术构建8周龄SPF级C57BL/6N背景的LCMT1flox/+小鼠与Alb-Cre+/+小鼠雌雄各2只,体重18~22 g。该工作与赛业生物科技有限公司【SCXK(粤)2018-0032】合作完成。动物饲养在广西医科大学SPF级实验动物中心【SYXK(桂)2020-0003】,8周龄的WT鼠和KO鼠各5只,体重18~21 g。饲养环境:温度20~26℃,湿度40%~70%,氨浓度不超过14 mg/m3,每12 h进行明暗交替,垫料、鼠盒、水瓶高压灭菌。所有操作均符合实验动物伦理学要求(伦理审查备案号:202107001)。
1.1.2 主要试剂与仪器
鼠尾DNA提取试剂盒、逆转录试剂盒、DL2000 DNA Marker(TaKaRa公司);TRIzol(天根生化科技有限公司);SYBR Green Master(Roche公司);I型核酸染料(北京索莱宝科技有限公司);琼脂糖粉、PVDF膜、化学发光液(Invitrogen公司);蛋白预染Marker(Thermo Fisher Scientific公司);BCA蛋白浓度测定试剂盒、Anti-Tubulin(上海碧云天生物技术有限公司,货号:AF0001);Anti-LCMT1(上海圣克鲁斯生物技术有限公司,货号:SC-81609);山羊抗鼠IgG/HRP、山羊抗兔IgG/HRP(北京中杉金桥生物技术有限公司);4%多聚甲醛通用型组织固定液(大连美仑生物技术有限公司)。
PCR仪、iBright蛋白免疫印迹成像系统、-80°C超低温冰箱(Thermo Fisher Scientific公司);水平凝胶电泳装置、电泳槽、电泳仪(Bio-Rad公司);凝胶扫描成像系统(北京天根生化科技有限公司);超纯水仪(Millipore公司);超净工作台(苏州净化设备有限公司);水平摇床(上海智城分析仪器制造有限公司)。
1.2 方法
1.2.1 肝LCMT1敲除小鼠的构建策略
LCMT1基因位于小鼠7号染色体,有11个外显子,在2号和3号外显子两端插入Loxp位点(图1A)。根据CRISPR/Cas9技术原理,用显微注射法将sgRNA、Cas9蛋白和打靶载体注射到小鼠受精卵。经胚胎移植,获得阳性F0代小鼠。由阳性F0代小鼠与野生型小鼠交配得到F1代(LCMT1flox/+)杂合子小鼠。
1.2.2 肝LCMT1敲除小鼠的繁殖流程
将引进的LCMT1flox/+小鼠与Alb-Cre+/+小鼠进行交配,筛选出子代基因型为LCMT1flox/+/Alb-Cre+/-的小鼠进行自交,最终获得子代基因型为LCMT1flox/flox/Alb-Cre小鼠(KO小鼠)和Alb-Cre-/-小鼠(WT小鼠)(图1B)。

图1 LCMT1敲除小鼠的构建策略与流程图Figure 1 strategy and flow chart of LCMT1 knockout mouse
1.2.3 肝LCMT1敲除小鼠的基因型鉴定
对4周龄小鼠编号,剪下约1 cm鼠尾,置于EP管内,在冰上剪碎小鼠尾巴,加入180μL的Buffer GL、20μL的Proteinase K和10μL的RNase A,混匀后于56℃水浴锅消化过夜提取DNA。取1μL DNA进行PCR扩增,扩增产物在1%琼脂糖凝胶电泳。电泳完成后在凝胶扫描成像系统显影,根据对应片段长度进行基因型鉴定。Loxp基因的上游引物:5′-GTGCAGGCATCAGAGAGACAAGC-3′,下游引物:5′-GACAAGGTCTCACTTCATATCCTGG-3′;Cre基因上游引物:5′-GAACGCACTGATTTCGACCA-3′,下游引物:5′-GCTAACCAGCGTTTTCGTTC-3′。
1.2.4 观察小鼠的基本情况和繁育能力
观察两组小鼠的饮食、饮水、活动、生长发育状态和繁殖能力等。
1.2.5 小鼠肝LCMT1基因mRNA水平检测
(1)总RNA提取:取8周龄小鼠新鲜肝,置于EP管中,加入150μL TRIzol,用匀浆器研磨使组织充分溶解,再加入850μL TRIzol溶液,经酚/氯仿法提取总RNA,使用Bio Spec-nano分光光度计测定RNA浓度。
(2)RNA逆转录:利用TaKaRa公司的逆转录试剂盒逆转录得到cDNA。
(3)实时定量PCR体系:反应体系总共20μL:ddH2O 3μL,SYBR Premix(2×)10μL,前后引物各1 μL,cDNA 5μL。PCR仪扩增条件:95℃预变性1 min,95℃变性20 s,60℃退火延伸1 min,40个循环。内参β-actin上游引物:5′-GTGACGTTGACATC CGTAAAGA-3′,下游引物:5′-GCCGGACTCATCGTA CTCC-3′。目 的 基 因 LCMT1 上 游 引 物:5′-AGAAGGAACGGCTGCTGTTGAATG-3′,下 游 引 物:5′-CTGCCCGAGGTAGCCCACTG-3′。
1.2.6 Western Blot
(1)组织总蛋白提取:随机选取LCMT1-KO小鼠和WT小鼠各3只,使用RIPA裂解液提取小鼠肝、大脑、心和肾组织总蛋白,利用BCA蛋白定量试剂盒测定蛋白浓度。
(2)SDS-PAGE电泳:采用10%的SDS-PAGE凝胶电泳2 h,转膜1.5 h,37℃摇床封闭1 h,4℃过夜孵育LCMT1一抗(1∶1000),次日用TBST洗膜3次,每次10 min,加入山羊抗兔IgG(1∶4000),37℃孵育2 h。TBST洗3次,每次10 min。
(3)化学发光显影:化学发光剂A液和B液按1∶1配置,用枪头向PVDF膜加适量发光液,并小心涂抹均匀,用iBright蛋白免疫印迹成像系统显影。
1.2.7 肝苏木精-伊红染色
小鼠肝置于4%多聚甲醛组织固定液中,石蜡包埋,切片,苏木精-伊红染色,观察肝形态学。
1.2.8 LCMT1 sgRNA脱靶分析
登录Cas-OFFinder网站(http://wwww.rgenome.net/cas-offinder/),将sgRNA序列录入,以小于等于3个碱基错配作为判断标准,脱靶信息如表1所示,其中红色代表靶序列与sgRNA系列不同的碱基。根据脱靶位点的DNA序列设计PCR引物,对LCMT1-KO小鼠鼠尾DNA进行扩增,琼脂糖凝胶电泳,切胶回收纯化目的片段,测序检测脱靶情况。

表1 脱靶位点信息Table 1 Off-target information
1.3 统计学分析
采用SPSS 24.0软件进行统计学分析,采用平均值±标准差(±s)Graphpad Prism 8软件绘图。两组数据比较采用独立样本t检验,以双侧P<0.05表示差异具有统计学意义。
2 结果
2.1 肝LCMT1敲除小鼠基因型鉴定
小鼠交配产生子代的基因型鉴定结果见图2。使用Loxp引物和Cre引物对小鼠的基因型进行鉴定,若DNA中存在LCMT1flox/flox,则能扩增出234 bp的片段,否则只有172 bp的片段;若有Cre基因,则能扩增出204 bp的片段,反之则无法扩增出条带。如图2A所示1、2、3、5、6、8均为带有234 bp的LCMT1flox/flox基因目的条带;如图2B所示2、3、5、6均为带有204 bp的Cre基因目的条带。其中2、3、5、6为LCMT1-KO小鼠,1、4、7、8为WT小鼠,9、10、11为空白对照。

图2 小鼠基因型鉴定Figure 2 Mouse genotype identification
2.2 小鼠的一般情况
8周龄的WT小鼠和KO小鼠每日饮水、饮食和体重无明显差异。两组小鼠生长发育良好,毛发柔顺有光泽(见图3)。

图3 小鼠的一般情况Figure 3 General situation of mouse
2.3 小鼠的繁育情况
LCMT1-KO母鼠每胎产5~9只幼鼠,成活率>90%,其中LCMT1-KO小鼠与WT小鼠的雌雄比例如图4所示,LCMT1-KO小鼠与WT小鼠繁殖情况无明显差异。
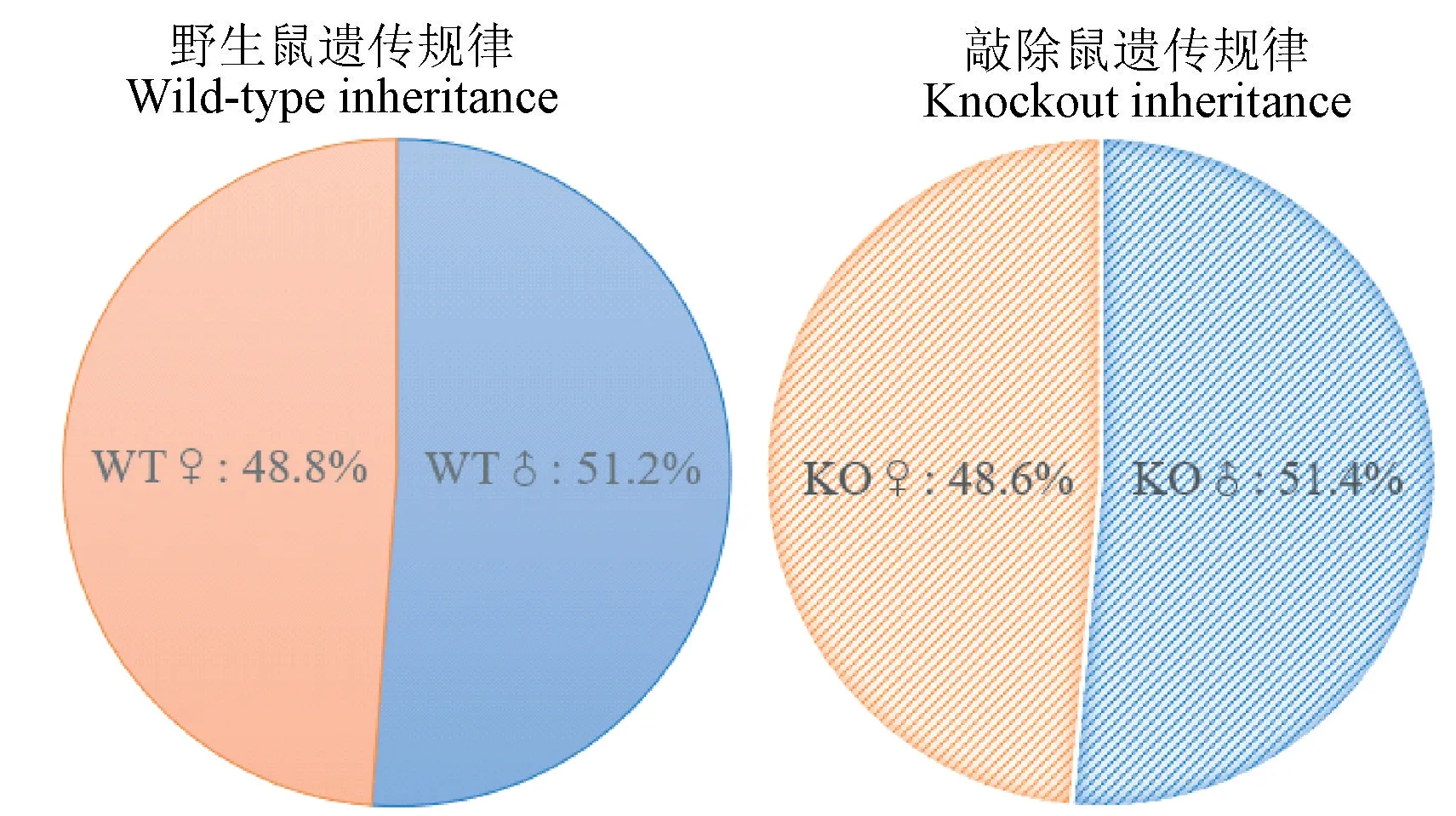
图4 小鼠的繁殖规律Figure 4 Reproduction law of mouse
2.4 小鼠肝LCMT1的mRNA和蛋白水平
实时定量PCR结果显示LCMT1-KO小鼠与WT小鼠相比肝中LCMT1 mRNA水平显著降低,具有统计学差异(P<0.001)(见图5A)。Western Blot结果显示,与WT小鼠相比,LCMT1-KO小鼠肝LCMT1蛋白表达显著降低(P<0.01)(见图5B)。

图5 肝LCMT1基因表达和蛋白表达Figure 5 Liver LCMT1 gene expression and liver LCMT1 protein expression
2.5 小鼠肝组织形态学改变
两组小鼠肝质地柔软、边缘锐利、表面光滑无结节,呈棕红色(见图6A)。HE染色结果显示两组小鼠肝小叶以中央静脉(CLV)为中心,呈放射性排列整齐,肝细胞核呈紫蓝色,细胞质着色均匀呈红色。未发现脂肪滴异常积累和炎细胞浸润(见图6B)。

图6 小鼠肝组织及HE染色Figure 6 Liver of mouse and HE staining
2.6 小鼠肝外组织LCMT1的蛋白水平
Western Blot检测小鼠LCMT1在肝外组织的表达,两组小鼠LCMT1的表达在心脏、大脑和肾组织中没有显著差异(见图7)。

图7 肝外组织LCMT1蛋白表达Figure 7 LCMT1 protein expression in extrahepatic tissue
2.7 LCMT1敲除小鼠脱靶位点分析
LCMT1-KO小鼠PCR产物的测序峰图如图8所示,在可能脱靶的位点处未发现碱基突变、移码、套峰、少峰和乱峰等现象,因此LCMT1-KO小鼠sgRNA未发生脱靶。
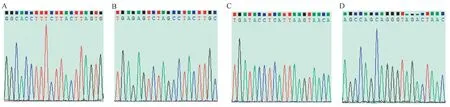
图8 PCR产物的测序峰图Figure 8 Sequencing peak map of PCR products
3 讨论
LCMT1催化PP2Ac甲基化,调节PP2A的活性和生物学功能[17]。PP2A在真核生物中广泛表达,通过调节关键蛋白的去磷酸化,参与细胞代谢、细胞分化和细胞周期等生理活动,已有大量报道显示PP2A在肿瘤、阿尔兹海默病和糖尿病等多种疾病的病理进程中发挥关键性作用[18]。LCMT1作为细胞内唯一催化PP2Ac甲基化的酶,其对机体的作用尚未知。为探讨LCMT1在体内的功能,既往研究尝试构建全身LCMT1敲除小鼠,但结果显示LCMT1敲除纯合鼠子代存活率下降,在胚胎期12.5 d时LCMT1蛋白几乎全部失活,胎儿肝发育和造血功能受损,基因敲除鼠通常在胚胎期的14.5~16.5 d死亡[13],无法满足后续功能研究需求,同时也提示LCMT1对小鼠胚胎早期发育具有不可或缺的作用,对LCMT1功能的体内研究需采取条件敲除的策略。
既往研究结果显示,PP2A在肝糖代谢发挥重要作用。肝特异性敲除PP2Ac小鼠对外周葡萄糖的清除能力增强,促进糖原沉积,抑制糖异生,增加小鼠糖代谢稳态[19]。课题组前期研究发现肝细胞MIHA中siRNA LCMT1,PP2Ac去甲基化水平和糖原合成增加,糖异生基因的表达水平降低[20],提示LCMT1可能通过调控PP2A在肝糖代谢中发挥重要作用。为进一步探讨肝LCMT1在体内的功能,本研究采用CRISPR/Cas9方法构建肝特异LCMT1敲除小鼠。由于LCMT1的敲除作用是通过Alb-cre对两个同向LoxP位点的特异性识别剪切而实现,而肝特异Alb-cre小鼠的Alb-cre活性仅局限于肝,在胚胎发育早期并未对全身的LCMT1位点发生剪切,因而小鼠胚胎得以发育存活,从而实现肝特异性敲除LCMT1小鼠的成功构建。
本研究构建成功的基因敲除鼠,经基因测序未发现sgRNA脱靶,采用实时定量PCR和Western Blot检测小鼠各脏器LCMT1的表达,显示肝LCMT1水平显著降低,而在肝外组织中的表达没有显著变化。结果表明肝LCMT1特异性基因敲除小鼠表型符合预期。在目前完成的8周饲养时长中,该基因敲除小鼠生长发育良好,毛发柔顺有光泽,繁殖子代符合孟德尔遗传定律,子代存活率高。解剖见肝发育良好表面无结节,HE染色显示小鼠肝细胞排列整齐,未见细胞变性和坏死等病变,未见与野生型鼠存在显著性差异。后续研究将探讨在高糖高脂饮食、衰老等代谢压力下,肝LCMT1基因特异敲除对小鼠糖脂代谢的影响,以进一步明确LCMT1在肝的功能。
猜你喜欢
传染病信息(2022年4期)2022-11-23
分子催化(2022年1期)2022-11-02
西部医学(2022年9期)2022-09-26
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
中学生物学(2020年10期)2020-12-25
电脑知识与技术(2018年19期)2018-11-01
生物学教学(2018年8期)2018-09-03
医学研究杂志(2015年12期)2015-06-10
