
137份茶树人工杂交F1代单株表型性状及遗传多样性分析
2022-03-12 05:31邓少春田易萍陈林波陈春林庞丹丹徐丕忠包云秀朱兴正
山东农业科学 2022年2期
邓少春,田易萍,陈林波,陈春林,庞丹丹,徐丕忠,包云秀,朱兴正
(云南省农业科学院茶叶研究所/云南省茶树种质资源创新与配套栽培技术工程研究中心/云南省茶学重点实验室,云南 勐海 666201)
金萱(Camellia sinensiscv.Jinxuan)为灌木型、中叶类、中早生茶树品种,选育自台湾省,在广东、广西、福建等地引种较多[1,2],适制乌龙茶,兼制绿茶。云茶1号(Camellia sinensisvar.assamicacv.Yuncha 1)为乔木型、大叶类、早生无性系茶树品种,从云南省元江细叶糯茶群体品种中采用单株育种法育成,在云南西双版纳、普洱、保山等州(市)有种植,适制红茶、绿茶、普洱茶。金萱与云茶1号芽叶表型差异较大[3]。
杂交育种是创制新种质及发掘新变异的最有效方法[4-6]。杂交育种是否成功取决于杂种优势是否在杂交后代中充分显现[7,8],同时,培育一个新品种要建立在对杂交后代表型性状及遗传规律研究的基础上[9]。王治会等[10]对17份黄金菊自然杂交后代单株的23项表型性状的变异与多样性进行研究,结果表明单株的性状变异较丰富;部分单株的简单儿茶素和游离氨基酸含量表现为显著提高,而总儿茶素、酯型儿茶素及茶多酚含量表现为显著降低。杨军等[11]为实现茶树种质资源的系统分类,利用17对SSR引物对94个茶树种质资源的亲缘关系、遗传多样性与群体结构进行分析,结果将这94个茶树种质分为3个类群,分别为闽南高香资源类群属性、闽北乌龙茶种质资源类群属性、乌龙茶与绿茶种质资源混合类群属性。王小萍等[12]对古蔺县139份野生大茶树资源(7个天然居群)叶片18个表型性状的遗传多样性分析结果显示野生大茶树资源叶片性状变异丰富,各性状在各居群间的变异系数最大值均超过10%,UPGMA聚类分析结果显示这7个居群的亲缘关系较近。
芽叶能为植物的生长发育及开花结实提供充足的养分保障,是植物最重要的农艺性状之一。研究表明,经济型树种的品质和产量与芽叶性状之间存在较大的关联[13-15],要对杂交后代进行早期预筛和鉴定,芽叶性状表型特征的鉴定是必不可少的环节[16-18]。本试验分别以中叶种金萱和大叶种云茶1号为母本和父本进行杂交,得到137份杂交后代材料,对这些材料及其父母本的芽叶特征进行观测和分析,并探究其遗传规律,以期了解大中叶种亲代和子代的表型特征及遗传变异规律,为茶树杂交育种提供理论指导。
1 材料与方法
1.1 试验材料
以云南省农业科学院茶叶研究所试验基地栽种的金萱(♀)、云茶1号(♂)及其人工杂交育成的F1代单株为材料(编号1~137)。
1.2 试验方法
以茶树种质资源数据质量控制规范为参考[19],于春、夏、秋三季调查供试材料的芽叶性状,并于冬季采茶结束时调查成熟叶性状。对芽叶表型数量性状共12个指标进行测量和统计。
相对遗传力:假设基因的加性效应严格控制子代的遗传特性,对应的相对遗传力为a1、a2,则大值亲本表现型平均值的相对遗传力a1=(F1-P2)/(P1-P2),小值亲本表现型平均值的相对遗传力a2=(P1-F1)/(P1-P2),其中P1为大值亲本,P2为小值亲本[20]。
杂种优势:超亲优势表示为Hb、中亲优势表示为Hm、超亲优势率表示为RHb、中亲优势率表示为RHm。子代某一性状的平均值表示为Fm、双亲平均值表示为MPV,双亲中较大亲本值表示为BPV。计算公式如下:Hb=Fm-BPV,Hm=Fm-MPV,RHb=(Hb/BPV)×100[2],RHm=(Hm/MPV)×100。
1.3 数据处理与分析方法
利用IBM SPSSStatistics 26和Microsoft Excel 2016进行正态性检验、频率分析、方差分析、描述统计和聚类分析,叶片表型遗传变异使用CV(变异系数)和H′(Shannon-Weaver遗传多样性指数)进行评价,叶片表型组间相关程度和方向用Pearson’s相关系数来度量,芽叶表型性状的遗传模型使用杂种优势理论和相对遗传力进行分析。
2 结果与分析
2.1 离散特征
由表1可知,人工杂交F1代12个芽叶表型性状的遗传多样性丰富,变异系数为6.37%~32.84%,极大值与极小值的比值为1.48~9.19,变异系数超过10%的性状有11个。叶面积、芽重、一芽二叶重和一芽三叶重的变异系数均超过30%,变异较大;其余性状除叶长叶宽比、芽长、一芽一叶长外变异系数均在15%~30%,为中等变异水平。叶面积的标准差最大,群体中极易出现极值个体,芽重的标准差最小。从图1可以看出,各表型性状均符合多基因控制数量性状遗传特征,具有较好的连续性正态分布趋势。

图1 F1代12个芽叶表型性状的频率分布
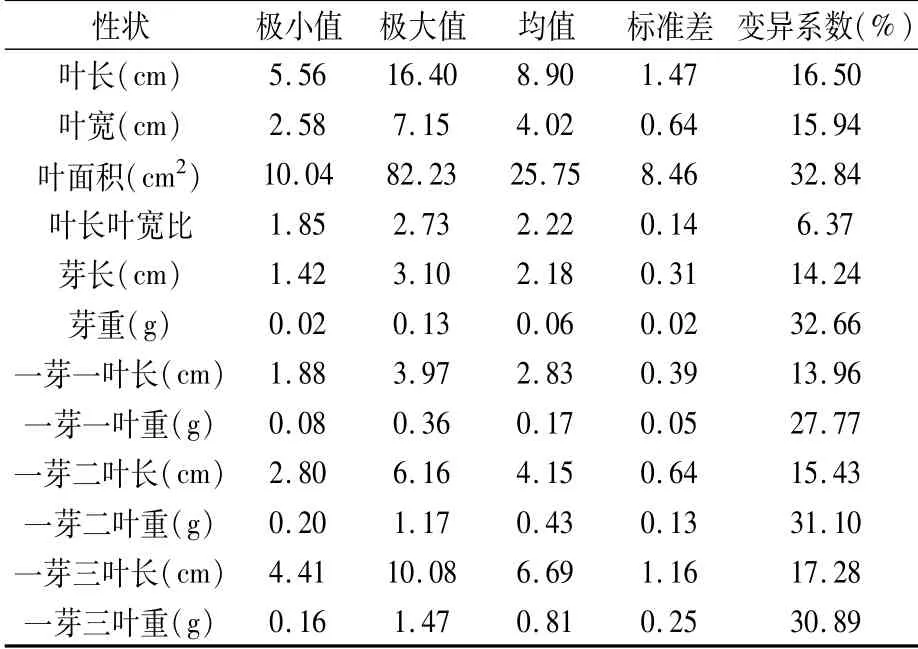
表1 芽叶表型性状在F1群体分离的特征值
2.2 相关性分析
由表2可知,12个芽叶性状间的66对相关性分析中有56对达极显著水平(P<0.01),2对达到显著水平(P<0.05),相关系数在0.85以上的有14对,达极显著水平,相关程度极高。其中,叶长叶宽比与一芽二叶长、一芽三叶长呈正向弱相关关系;叶长叶宽比与叶宽、一芽一叶重、一芽二叶重、一芽三叶重呈负相关关系;而叶长叶宽比与芽重则不存在直线相关关系。
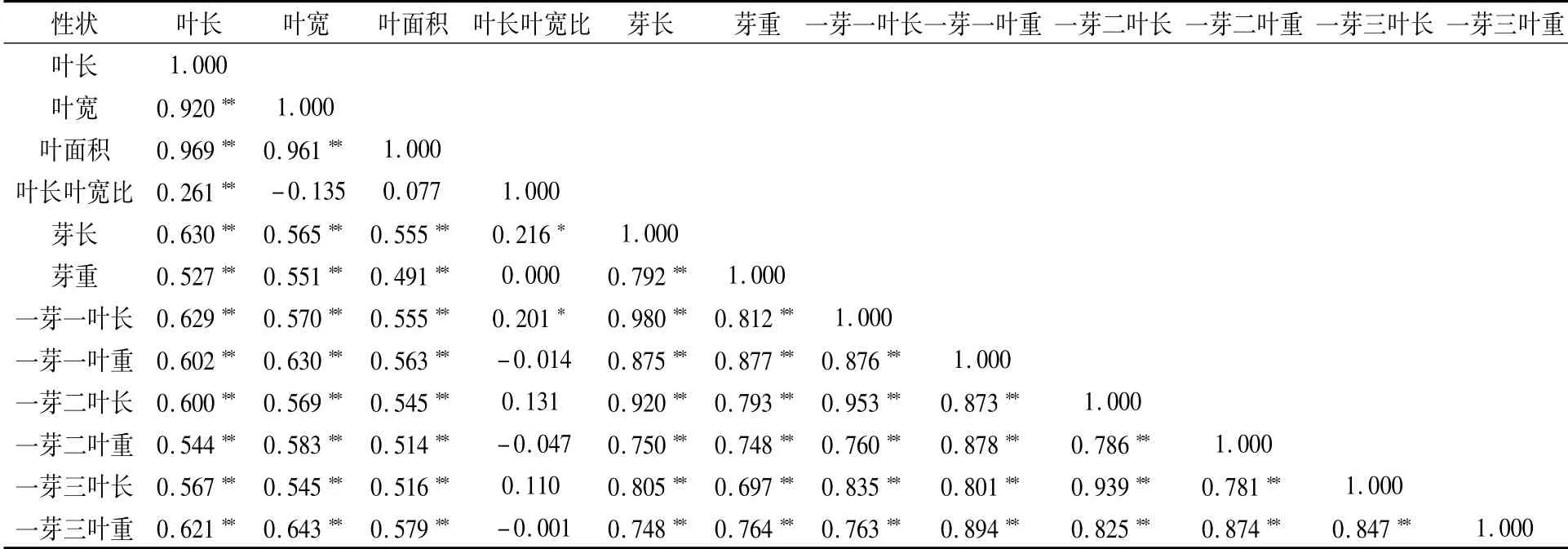
表2 F1代芽叶表型性状的相关性分析结果
2.3 杂种优势表现
如表3所示,金萱和云茶1号杂交得到的F1代单株叶长、芽重、一芽一叶重、一芽二叶重、一芽三叶长、一芽三叶重为负向显性;仅叶长叶宽比的遗传方向为正向显性;叶宽、叶面积、芽长、一芽一叶长、一芽二叶长的相对遗传力表现为负向超显性。从父母本相对F1代的遗传力强弱来看,父本云茶1号在叶长叶宽比、芽长、芽重、一芽一叶长、一芽一叶重、一芽二叶长、一芽二叶重、一芽三叶长、一芽三叶重九个表型性状中的相对遗传力强于母本,母本金萱在叶长、叶宽、叶面积三个性状中的相对遗传力强于父本。另外,12个性状在F1代的中亲优势率为-0.53%~0.01%,超亲优势率为-0.69%~-0.03%,其中叶长叶宽比的中亲优势为正值。

表3 F1代芽叶表型性状的杂种优势表现
2.4 聚类分析
由系统聚类分析结果(图2)可知,父母本及F1代群体在遗传距离为25处可划分为2个类群,第1个类群仅包括2个单株,占群体的1.44%,第2个类群分为2个亚类,分别包括1个单株和136个单株,占群体的98.56%。母本金萱被分在第1类群,而父本云茶1号则被划分在第2类群。结合137份供试材料的芽叶数量性状可以看出,第1类群的芽长、芽重、一芽一叶长、一芽一叶重、一芽二叶长、一芽二叶重、一芽三叶长、一芽三叶重等8个指标均高于第2类群;而叶长、叶宽、叶面积、叶长叶宽比4个指标均低于第2类群,尤其是叶面积,比第2类群低59.02%。

图2 杂交F1代单株系统聚类分析结果
3 讨论与结论
杂交育种是目前较为有效的获得植物新品系和突破性品种的方法之一,杂交后代不同群体之间具有性状分离的特征,且性状出现大幅度变异的可能性较大[21-23]。在本研究中,12个芽叶性状均具有大幅变异的特征,说明F1代具有较高的离散程度,表型多样性和变异水平也较为丰富,这为下一步选择优良单株提供了方向和材料。
杨亚军[24]研究发现,基因型重组决定了茶树无性后代的叶片小于有性杂交后代,且杂交后代具有与母本更相近的表型特征,且叶宽与叶面积表型具有很大的相关性,叶宽具有比叶长更高的遗传力。本研究结果表明,母本在叶长、叶宽、叶面积三个性状中的相对遗传力强于父本,这与杨亚军[24]的结论一致;但通过计算父母本相对遗传力发现叶宽的遗传特性较复杂,F1代叶长、叶宽与叶面积均呈极显著正相关,且相关系数大小相当,这与杨亚军[24]的结论有差异。
在本研究中,云茶1号和金萱分别作为父母本,杂交得到的F1代单株在12个性状上的表现为:9个性状表现为亲父特征,3个性状表现为亲母特征;6个性状遗传方向为负向显性,1个性状遗传方向为正向显性,5个性状的相对遗传力表现为负向超显性。说明云茶1号和金萱杂交后代各性状的遗传特性较复杂,出现变异的几率较大,可进一步扩大表型测定范围及F1代单株数量,以获得更多变异类型及具有优良性状的F1后代。
本研究的系统聚类分析结果将F1代群体划分为2个类群,第1个类群仅包括2个单株,其明显特征为叶面积偏小,叶片大小为中小叶,这为进一步挖掘茶树抗寒品种、选育不同产茶区适应性品种提供了思路,在今后的遗传测定和种质资源开发利用中应当予以特别关注。另外,在系统聚类时,父母本被分在了两个不同类群,说明父母本的性状差异较明显,可在杂交育种及特异资源培育方面加以利用。
猜你喜欢
农学学报(2022年8期)2022-11-16
农业与技术(2022年16期)2022-09-01
中国现代医生(2022年21期)2022-08-22
瞭望东方周刊(2021年6期)2021-03-30
瞭望东方周刊(2021年6期)2021-03-30
三农资讯半月报(2020年2期)2020-03-09
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
江苏农业科学(2017年17期)2017-11-15
江苏农业科学(2016年9期)2016-11-28
