
富贵草叶斑病病原的鉴定
2022-03-08 06:43钟杰尹秀娟钟双玉陈锦朱俊子李晓刚
湖南农业大学学报(自然科学版) 2022年1期
钟杰,尹秀娟,钟双玉,陈锦,朱俊子,李晓刚*
(1.湖南农业大学植物保护学院,湖南 长沙 410128;2.湖南省农业科学院农业环境生态研究所,湖南 长沙 410125)
富贵草(Pachysandra terminalisSieb. et Zucc)为黄杨科板凳果属多年生常绿草本植物或亚灌木,又称为顶花板凳果、顶蕊三角咪、转筋草、吉祥草等。富贵草具有重要的药用价值,从富贵草中提取的孕烷生物碱具有抗菌和抗癌功效[1]。富贵草分布较广,因其观赏效果好,且耐寒、耐阴、易繁殖,常作为盆栽和绿化植物[2]。
2019 年5 月,在湖南长沙一中药材苗圃的富贵草上发生了一种不明原因的叶部病害,约20%的植株叶片受到侵染。笔者对这种富贵草叶斑病进行了病原菌分离,结合形态学和分子生物学鉴定及科赫氏法则验证,确定了该病害病原,以期为病害的科学防控提供依据。
1 材料与方法
1.1 材料
2020 年5 月,在湖南长沙一中药材苗圃摘取不明原因的富贵草叶斑病典型症状叶片。
1.2 方法
1.2.1 富贵草叶斑病病原菌的分离和鉴定
剪取患病叶片病健交界部大小5 mm×5 mm 的组织,用75%乙醇消毒30 s、0.1%升汞再消毒1 min后无菌水漂洗3 次,用无菌滤纸吸干水分,置于含50 μg/mL 链霉素的马铃薯葡萄糖琼脂培养基(PDA)上,26 ℃黑暗培养。将长出的菌落转接至新的PDA培养基,并通过单孢分离进行纯化后,于4 ℃ PDA斜面保存。
按照文献[3]的方法,刮取菌丝,提取病菌DNA。采用通用引物ITS4/ITS5[4]、ACT-512F/ ACT-783R[5]、CHS-79F/CHS-354R[5]和GDF/GDR[6]扩增核糖体转录间区 rDNA (ITS)、肌动蛋白基因(ACT)、几丁质合成酶1 基因(CHS1)和甘油醛-3-磷酸脱氢酶基因(GAPDH)序列(表1)。PCR扩增: 2×EsTaqMaster Mix 25 μL、总DNA 模板2 μL、10 μmol/L上、下游引物各2 μL、ddH2O 19 μL。PCR 反应程序:95 ℃ 预变性5 min;95 ℃变性30 s,56 ℃退火30 s,72 ℃ 延伸45 s,共30 个循环;最后72 ℃延伸10 min,4 ℃保存。

表1 用于PCR 扩增和测序的引物信息Table 1 Information of pr imers use d for PCR a mplification and sequencing
PCR 产物经1%琼脂糖凝胶电泳检测后送上海生物工程有限公司测序。所获序列提交GeneBank数据库,获得登录号(ITS、ACT、CHS1、GAPDH基因序列分别为MT645674、MT648689、MT663546和 MT663547),并进行同源性比对分析。利用MEGA 6 软件中邻接法(NJ)构建串联系统发育树,采用自举法(bootstrap)进行1000 次重复检验[7]。
1.2.2 富贵草叶斑病病原菌致病性的测定
采用菌丝块接种方法进行柯赫氏法则验证。将菌株在PDA 培养基上26 ℃培养7 d,用消毒后的5 mm 打孔器打取菌落边缘的菌丝块,接种于健康的富贵草叶片。接种前叶片经75%乙醇表面消毒,并将叶片针刺伤口。以无菌PDA 琼脂块为对照处理。将所有接种体置于含有湿润滤纸的保鲜盒26 ℃保湿培养,逐日观察发病情况,并对发病叶片再次进行病原菌分离鉴定。
2 结果与分析
2.1 富贵草叶斑病病原菌的分离和鉴定结果
富贵草叶斑病侵染症状如图1 所示。受害植株叶片从叶尖和叶缘开始,呈现不规则的黄褐色病斑,叶片健康部分变黄;随着病斑扩展,颜色变成深褐色,叶片枯萎凋落。

图1 富贵草叶斑病侵染症状Fig.1 Symptoms of leaf spot disease observed on Pachysandra terminalis leaves
共获得5 个培养性状一致的菌株,代表性菌株命名为FGC3-1。PDA 培养基26 ℃恒温培养,菌丝由白色逐渐转变成灰色,菌落呈圆形,边缘较平滑。培养7 d 左右,产生大量分生孢子,分生孢子单生、透明、呈镰刀形,大小为(20.5~27.5) µm×(3.8~4.2) µm (n=50)。从发病植株叶片病斑处镜检发现黑褐色刚毛,较僵直,末端尖锐,长约85~115 μm。分生孢子萌发形成附着胞,附着胞呈深褐色、形状近圆形或棍棒形,大小为(8.8~12.5) μm × (6.5~8.5)μm (图2-C、图2-D、图2-E)。病原菌形态特征与已报道的[8]冬麦刺盘孢(Colletotrichum liriopes)相似。

图2 富贵草叶斑病病原菌的形态Fig.2 Morphological characteristics of the isolated pathogens
对分离纯化后菌株的ITS、ACT、CHS1和GAPDH进行PCR扩增、克隆、测序,分别获得了长度为581、245、260、257 bp的片段序列。利用BLASTn进行同源性比对分析,结果表明,除ITS序列外,ACT、CHS1和GAPDH序列与冬麦刺盘孢(C. liriopes)菌株对应序列相似性为97.21%~100%(对应菌株登录号为MK644098.1 、 KY995503.1 、 KY995452.1 、MK644100.1等)。利用多基因构建系统进化树(序列登录号信息如表2所示),结果,FGC3-1与冬麦刺盘孢聚为一支(图3)。结合形态学和分子鉴定结果,可确定该病原菌为冬麦刺盘孢。

图3 基于ITS、ACT、CHS1 和GAPDH 序列的联合进化树Fig.3 Phylogenetic tree based on combined sequences of ITS, ACT, CHS1 and GAPDH
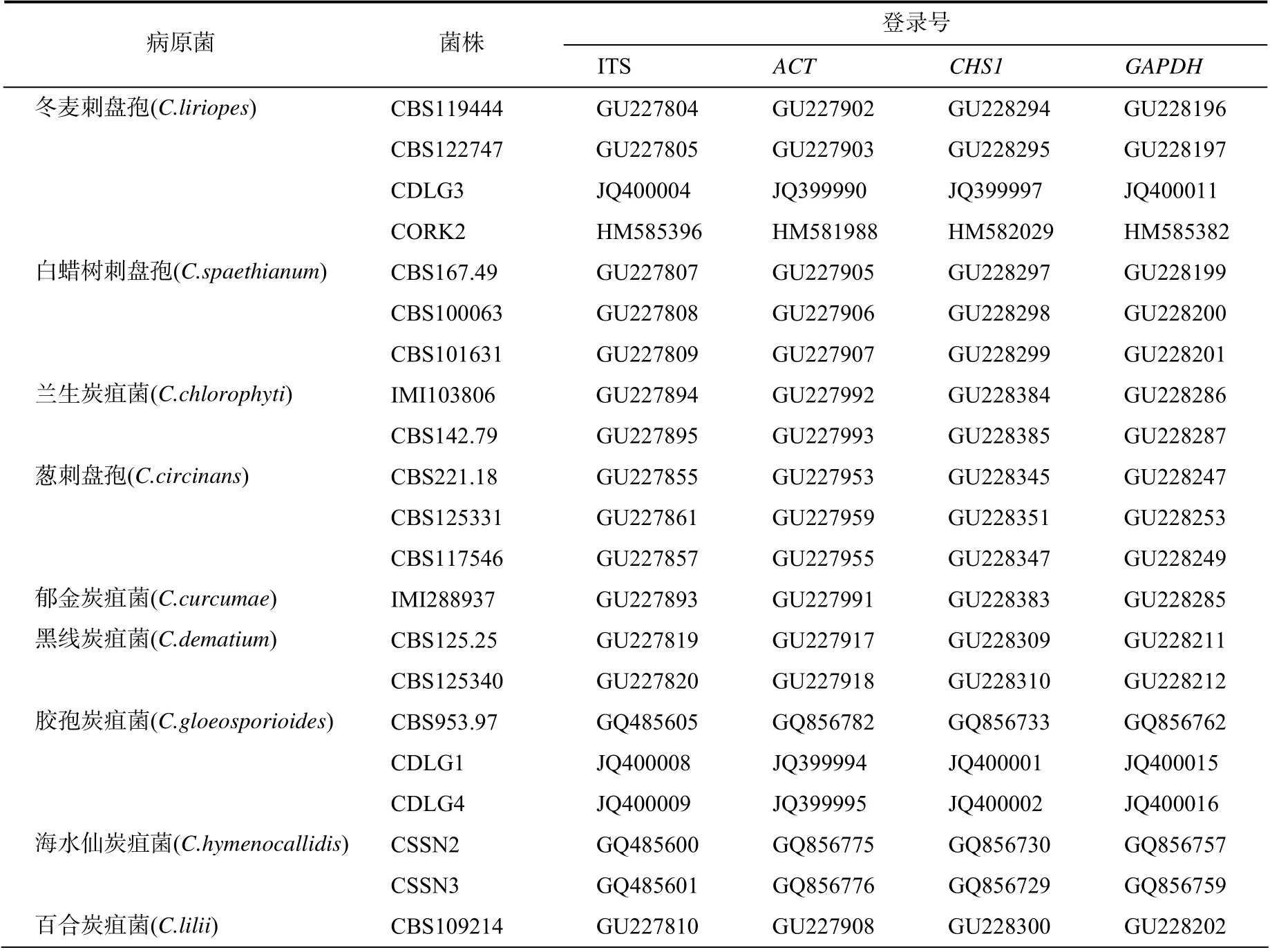
表2 用于系统发育分析的真菌菌株信息Table 2 Information of the fungal strains used for phylogenetic analysis

表2(续)
2.2 富贵草叶斑病病原菌的致病性
利用菌丝块接种富贵草健康叶片,7 d 后引起叶片发病。病斑呈褐色、近圆形或不规则形,病斑周围变黄,接种症状与田间自然发病症状相似。无菌PDA 琼脂块接种的对照叶片未发病。从接种病斑处可镜检到病原菌分生孢子,且可再次从病斑处分离并鉴定出相同的病原菌,由此证明接种的分离菌株为富贵草叶斑病的致病菌。
3 结论
对富贵草上不明原因叶斑病进行了症状观察、病原菌分离及鉴定,完成了科赫氏法则验证,证明该富贵草叶斑病病原为冬麦刺盘孢(C. liriopes)。冬麦刺盘孢也可导致其他植物病害:TRIGIANO 等[9]在美国报道了冬麦刺盘孢可引起万年青(Rohdea japonica)叶部炭疽病;CHEN 等[10]对青岛发生的山麦冬(Liriope spicata)叶斑病进行鉴定,确定其病原为冬麦刺盘孢;OO 等[11]鉴定了由冬麦刺盘孢引起的阔叶山麦冬炭疽病。此外,还有关于冬麦刺盘孢和胶孢炭疽菌(C. gloeosporioides)共同侵染引起兰花三七(Liriope cymbidiomorpha)叶斑病的报道[12]。YANG 等[13]在贵州、广西和辽宁等省份的萱草属植物茎秆病害中分离到了冬麦刺盘孢和其他炭疽菌。关于冬麦刺盘孢的寄主范围及是否存在致病力分化等还有待进一步研究。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
疯狂英语·新策略(2021年9期)2021-11-02
甘肃教育(2021年10期)2021-11-02
热带作物学报(2019年4期)2019-06-11
安徽农学通报(2017年23期)2017-12-27
现代农业科技(2016年23期)2017-04-06
现代农业科技(2009年17期)2009-03-08
