
上海地区桃果实腐烂病原菌分离及鉴定
2022-03-08 00:09张丽勍李雄伟石大艳余永婷方献平周慧娟张学英叶正文
上海农业学报 2022年1期
张丽勍,李雄伟,石大艳,余永婷,方献平,周慧娟,张学英,叶正文*
(1 上海市农业科学院林木果树研究所,上海 201403;2 上海市设施园艺技术重点实验室,上海 201403;3贵州师范学院,贵阳 550018;4 安顺学院,安顺 561000)
桃为蔷薇科桃属植物,起源于我国,迄今已有4 000 多年的栽培历史[1]。2017—2018年我国鲜食桃(含油桃)产量1 430 万t,居全球首位[2]。我国桃果实成熟期通常在6—8月份,采运季节常遇高温湿热天气,导致桃采后生理代谢旺盛,易变软、腐烂和遭受病虫害。一般来说,桃果实品质从优良至腐烂变质、不可食用的时间仅为2—3 d[3]。其中,真菌病害引起的桃腐烂最为严重,轻者腐烂率为10%—30%,重者高达50%以上,给我国桃产业造成了严重的经济损失[3]。2018年上海市桃树种植面积0.45 万hm2,产量7.40 万t,产值7.37 亿元人民币[4]。
上海地处长江入海口,属亚热带季风性气候,雨量充沛。2019年上海市6月17日入梅,7月20日出梅,梅雨期33 d(常年23 d),出梅偏晚,降水量明显偏多,为常年平均值的2.1 倍,导致桃烂果增多。梅雨季适宜的温度和潮湿的天气最有利于病原菌的生长和传播。目前,我国已报道的引起桃腐烂的病原菌有10 种[3,5-6]。本研究对上海桃产区的桃腐烂样品进行分离和鉴定,并通过柯赫氏法则进行验证,以期明确上海地区引起桃腐烂的主要病原菌种类,为其有效防控提供可靠的依据。
1 材料与方法
1.1 病样采集及病原菌的分离
2019年6—7月从奉贤区、浦东新区和金山区等上海桃产区采集桃树上病果70 份。采用组织分离法对病样进行分离纯化,纯化后的菌株置于4 ℃和-80 ℃各保存1 份。
1.2 病原菌形态学观察
将在PDA 平板上培养的病原菌用无菌打孔器在菌落边缘处打孔,将菌块置于新的PDA 平板上28 ℃黑暗培养,5 d 后对菌落形态特征进行记录并在光学显微镜(Leica DM 2700 M)下观察菌丝形态、分生孢子形态及测量大小。美澳型核果褐腐病菌(M.fructicola)分生孢子形态学观察参照Hu 等[7]方法,将桃果实接种褐腐病菌,培养4 d 后,挑取病斑表面产生的分生孢子进行观察。葡萄座腔菌(B.dothidea)分生孢子形态学观察参照Yuan 等[8]诱导产孢方法:取剪成长5 cm 左右的健康松针,121 ℃灭菌,放置在水琼脂培养基上,接入菌株,28 ℃培养至菌丝长满培养皿;随后置于20 ℃有光照的培养箱中继续培养。
1.3 基因组DNA 提取、PCR 扩增及菌株ITS 序列分析
用无菌枪头刮取PDA 平板培养的菌丝,采用真菌基因组DNA 提取试剂盒提取DNA, -20 ℃保存。对84 个菌株的核糖体转录间隔区(ITS) 片段进行PCR 扩增。PCR 扩增引物序列:ITS1 (5’-CTTGGTCATTTTAGAGGAAGTAA-3’)∕ITS4(5’-TCCTCCGCTTATTGATATGC-3’)[9]。产物测序由生工生物工程(上海)股份技术有限公司完成。将测定的序列在GenBank 中进行Blast 比对,根据比对结果确定归属。
1.4 多基因建树分析
根据ITS 测序结果,选取15 株褐腐菌代表菌株进行甘油醛-3-磷酸脱氢酶基因(G3PDH)和β-微管蛋白基因(TUB2)的PCR 扩增及测序。PCR 扩增引物序列:Mon-G3pdhF(5’-ACGGTCAATTCAAGGGTGAT-3’)∕Mon-G3pdhR(5’-ATCGAAGATGGAGGAGTGGT-3’)和Mon-TubF1(5’-ATGCGTGAGATTGTACGTAT-3’)∕Mon-TubR1(5’-GTACCAATGCAAGAAAGCCT-3’)[7]。采用DNASTAR 软件和CLUSTAL X 1.81 软件对褐腐菌15 株代表菌株测序的G3PDH和TUB2 基因序列与下载自GenBank 的参考序列进行序列比对分析[10]。利用MEGA v7.0.26 软件对其进行手动校正[11];利用贝叶斯法进行系统发育树的构建,在MrBayes v3.2.2 软件中,利用马尔可夫链蒙特卡罗(MCMC)算法对贝叶斯后验概率树进行系统发育分析。利用MrModeltest v2.3 软件的Akaike 信息准则(AIC)选择最佳核苷酸替换模型[12]。利用MrBayes v3.2.2软件进行进化树分析,当分裂频率的平均标准差低于0.01 时,停止分析。生成的进化树利用FigTree 1.4.3 软件进行可视化。如果后验概率≥0.95,则视为显著支持。所得系统发育树选用FigTree 1.4.3 软件进行分析。
构建G3PDH-TUB2 系统进化树时,利用B.fuckeliana的G3PDH基因(GenBank 序列号AJ705006)和TUB2 基因(GenBank 序列号Z69263)序列作外群。
1.5 致病性测定
按照柯赫氏法则将分离到的病原菌进行回接。本试验利用黄桃品种‘锦绣’进行接种测定,分为刺伤和无刺伤接种。选取外观整齐、成熟度及大小相似、健康的桃果实,用体积分数为75%的乙醇对桃果实表面进行消毒后,无菌水冲洗3 次。刺伤接种:用无菌接种针在桃果实表面刺一个约5 mm 深的孔,同时用打孔器从新鲜菌落的边缘(PDA 平板上培养5 d)取直径5 mm 的菌饼,倒置(菌丝向下)放于桃果实表面的针孔上。无刺伤接种:用打孔器从新鲜菌落的边缘(PDA 平板上培养5 d)取直径5 mm 的菌饼倒置于桃果面上。将接种后的桃果实置于塑料盒中,盒子的底部放入润湿的吸水纸以保证培养湿度近100%,并盖上塑料盖。将塑料盒置于25 ℃的人工气候培养箱中,光照条件设为12 h 光照∕12 h 黑暗的交替,培养48 h 后去掉塑料盖。每隔24 h 对发病情况进行记录并测量病斑直径。刺伤接种和无刺伤接种的对照均取直径5 mm 的PDA 琼脂块倒置于桃果面,重复5 次。
2 结果与分析
2.1 病原菌的分离
对70 个病果进行病菌分离,共分离到84 个真菌菌株,根据菌落形态将菌株分为5 种类型,分别命名为SP1、SP2、SP3、SP4 和SP5,各类型菌落的分离率依次为75.00%、8.33%、8.33%、4.76%和3.57%。从图1 可以看出,SP1 菌落为灰褐色,边缘较为整齐,无玫瑰花瓣结构;SP2 菌落初期为白色,后从中央转为紫色,絮状菌丝;SP3 菌落颜色为白色、奶油色;SP4 菌落颜色初期为白色,后从中央开始转为灰色;SP5 菌落初期为白色,后从中央开始转为灰绿色,菌落蓬松。
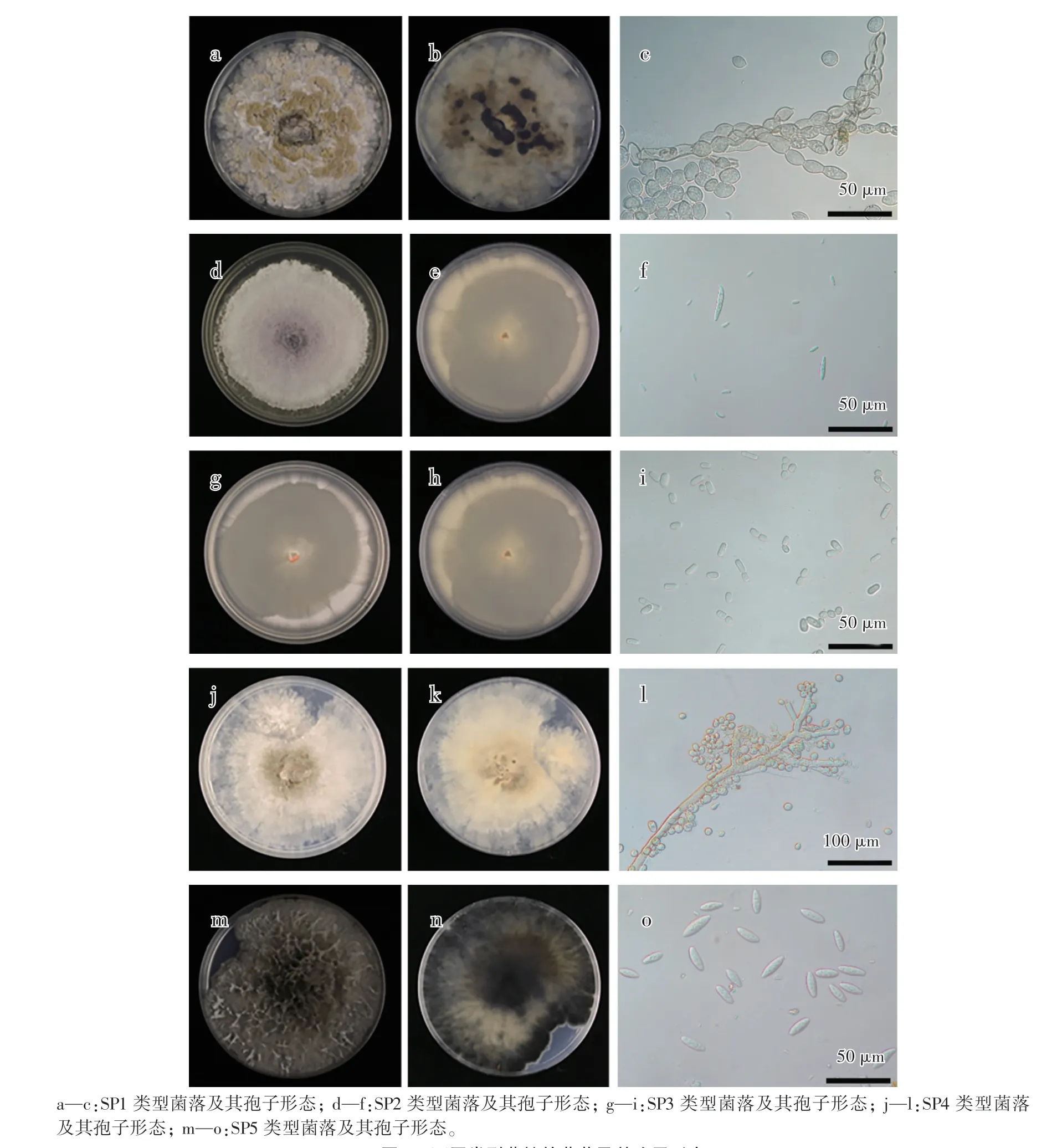
图1 不同类型菌株的菌落及其孢子形态Fig.1 Morphology of colony and spore of different pathogen species
2.2 病原菌鉴定
2.2.1 病原菌的形态学鉴定
SP1 的分生孢子透明,呈椭圆形、柠檬形或卵圆形,单细胞,链状,有分枝(图1),分生孢子大小为(10.2—27.8)μm× (7.1—18.4)μm(图1c)。SP2 的小孢子细长、椭圆形、无隔或具有一个隔膜,大小为(5.5—12.3)μm×(2.2—3.8)μm;大型分生孢子透明、弯曲、镰刀形,有1—5 个分隔,大小为(28.0—48.2)μm×(3.6—4.3)μm(图1f)。SP3 的孢子呈长方形两端略钝圆近方形或圆柱形,单细胞,大小为(5.0—12.5)μm×(2.5—4.0)μm(图1i)。SP4 的分生孢子呈卵圆形或椭圆形,单细胞,无色,分生孢子大小为(8.1—14.5)μm×(7.5—9.0)μm(图1l)。利用松针诱导的SP5 代表菌株呈纺锤形或梭形,薄壁、无隔、无色透明,分生孢子大小为(16.0—22.0)μm×(4.5—9.0)μm(图1o)。
2.2.2 病原菌的分子生物学鉴定
对5 种类型的代表菌株进行ITS 序列的扩增,扩增片段长度分别为537 bp(SP1)、564 bp(SP2)、376 bp(SP3)、537 bp(SP4)和585 bp(SP5);GenBank 登录号分别为MN689861、MN689858、MN689859、MN689856 和MN689841。BLAST 分析比对表明:5 种类型的代表菌株与美澳型核果褐腐病菌(M.fructicola) (MH860465.1)、 层 出 镰 孢 菌(F.proliferatum) (MK336501.1)、 白 地 霉(G.candidum)(KY495323.1)、灰霉菌(B.cinerea)(KT723007.1)和葡萄座腔菌(B.dothidea)(KC218814.1)的一致性分别为98.88%、99.82%、99.05%、99.25%和98.97%,覆盖率(Query coverage)分别为99%、96%、100%、99%和99%。根据比对结果,将上述5 种类型病原菌分别鉴定为M.fructicola、F. proliferatum、G.candidum、B.cinerea和B.dothidea。
对上海地区分离的15 株褐腐菌菌株分别扩增G3PDH和TUB2 基因序列(GenBank 登录号见表1),构建G3PDH-TUB2 多基因序列的系统发育树。结果表明,15 株菌株均以高支持率(贝叶斯后验概率≥0.95)与美澳型核果褐腐病菌(M.fructicola)菌株聚为1 个进化分支(图2)。

图2 基于G3PDH-TUB2 基因序列构建的15 株褐腐菌和14 株参考菌株的系统发育树Fig.2 Phylogenetic tree of 15 Monilinia isolates and14 reference isolates constructed based on G3PDH-TUB2 gene sequence

表1 Monilinia 菌株信息及GenBank 登录号Table 1 Monilinia isolates information and GenBank accession numbers
2.3 致病性
将5 种类型病原菌的代表菌株分别接种至‘锦绣’黄桃成熟果实,25 ℃培养,刺伤接种和无刺伤接种3 d 后均可见明显的病斑(图3、表2)。对接种的果实进行再分离并纯培养,均可获得与原来病原菌性状一致的病菌。

表2 5 种不同病原菌在桃果实上的致病性(25 ℃,接种后6 d)Table 2 Pathogenicity of 5 different pathogen species on peach fruits(25 ℃,6 days after inoculation)

图3 5 种不同病原菌接种桃果实(‘锦绣’)后的发病症状Fig.3 Symptoms of peach fruits(‘Jinxiu’) inoculated with 5 different pathogen species
3 讨论
本研究表明,引起上海桃腐烂的病原菌有5 种,分别是美澳型核果褐腐病菌(M.fructicola)、层出镰孢菌(F.proliferatum)、白地霉(G.candidum)、灰霉菌(B.cinerea)和葡萄座腔菌(B.dothidea)。褐腐病是引起桃腐烂的重要病害之一,其引起的桃果实腐烂病的发生率轻者为20%左右,重者高达100%[13-14]。桃褐腐病在采前、采后运输、贮藏期间均可发生,造成严重的经济损失[15]。近年来,我国东部、东南部、中部和西部桃主要产区均出现大量感染褐腐病的报道[16-18]。Moniliniaspp.病原真菌在生物学性状、发生流行规律及致病性方面各有不同[19],因此明确上海地区桃褐腐病病原种类具有重要意义。该菌群分类较为复杂,利用多基因系统发育进化树结合形态特征分析,结果更为准确。本研究选取分离自上海不同桃产区的15 株菌株,通过形态学鉴定和G3PDH-TUB2 系统进化树分析,明确了上海地区引起桃褐腐病的主要病原菌为美澳型核果褐腐病菌(M.fructicola)。美澳型核果褐腐病菌也是引起我国桃褐腐病的最主要病原菌[7]。因此,应将M.fructicola作为上海地区褐腐病防治管理策略中的主要目标重点防治,避免造成严重的经济损失。
我国有关层出镰孢菌(F.proliferatum)的报道主要集中在水稻、玉米等作物上[20-21]。2018年,福建省宁德市首次报道了F.proliferatum为引起桃果腐病的新病原菌,且当地的桃果实受害率达20%[22]。葡萄座腔菌(B. dothidea)寄主广泛,在苹果上可以引起苹果轮纹病,且在潮湿的环境中侵染较严重。B.dothidea释放分生孢子的高峰期在5—6月份,在温度适宜的条件下,病菌的分生孢子在降雨持续4 h 后就开始大量释放[23]。B.dothidea在桃树主干、主枝和侧枝上可以引起流胶,造成病原性桃流胶病[24]。我国目前有关B.dothidea引起桃果腐的报道较少。近年来上海地区桃流胶病发生严重,B.dothidea可以在为害枝干上越冬,且可存活4—5年[25],而上海6—7月份的梅雨天气尤其适合该病的侵染及传播,这也间接加重了桃腐烂病发生,推测此为果实受害的主要原因。B.dothidea具体传播侵染机制还有待进一步研究。开花至果实采收期间湿度过大是桃灰霉病(B.cinerea)发生和流行的主要条件。在桃真菌病害发病初期,葡萄座腔菌引起的桃腐和桃灰霉病等不能依靠肉眼有效区分,建议对两种病害进行综合防治。在我国,由白地霉(G.candidum)侵染引起的酸腐病是桃果实在采后运输和贮藏中的主要病害[26]。本研究首次证实了白地霉在采前生产过程中对桃果实的侵染。Schnabel 等[27]报道了由G.candidum侵染引起的桃酸腐病是美国南卡罗来纳州夏季重要病害之一。G.candidum孢子对桃果实的致病能力,可能会由于其他病原真菌的存在而增加。Morris 等[28]早在1982年就在柑橘果实中观察到了G. candidum与青霉菌(Penicillium digitatum)的孢子之间存在协同作用。因此,研究白地霉孢子与其他病原菌孢子之间可能存在的协同致病机理具有重要意义。
本试验首次明确了上海地区夏季桃腐烂病害的主要病原菌,为各果园后期进行有效的综合防治及预测预报奠定了基础,同时对桃采后病害的防控具有重要的指导作用。
猜你喜欢
现代食品(2022年17期)2022-10-20
无线互联科技(2022年2期)2022-04-20
医学概论(2021年18期)2021-01-21
少儿科技(2021年5期)2021-01-20
江苏农业科学(2019年11期)2019-07-22
热带作物学报(2019年4期)2019-06-11
安徽农学通报(2017年23期)2017-12-27
中国当代医药(2016年30期)2017-01-07
中国质量万里行(2015年1期)2015-01-27
食品与生活(2014年8期)2014-10-20
