
时间尺度上草地物种多样性和地上生产力的关系
2022-03-07 08:04晏昕辉李美慧拓行行温仲明
草地学报 2022年2期
杨 雪, 晏昕辉, 李美慧, 拓行行, 张 博, 温仲明,2, 李 伟
(1.西北农林科技大学草业与草原学院, 陕西 杨凌 712100; 2. 中国科学院水利部水土保持研究所, 陕西 杨凌 712100;3. 中国科学院大学, 北京 111049; 4. 西北农林科技大学水土保持研究所, 陕西 杨凌 712100)
在过去的几十年里,关于物种多样性与生产力的关系一直是生态学研究的重要议题[1-6],但仍存在诸多争议。首先,是物种多样性影响生产力?还是生产力影响物种多样性?亦或是二者本来就没有相关关系[7]?其次,物种多样性-生产力之间的具体模式是什么?它们在时间和空间尺度上是否存在一致性?可能的机制有哪些[3,8-11]?研究清楚这些基础问题有助于我们更加深刻了解生态系统结构和功能的时间动态变化,以便更好的进行生态系统的保护和适应性可持续管理,尤其是在当今全球变化的大背景下。
研究表明,物种多样性-生产力的关系一般有以下几种模式:“单峰”型、“U”型、线性正相关、线性负相关以及不相关[2,12-14]。其中,“单峰”型是至今观察到的最普遍的模式,即物种多样性在低生产力水平和高生产力水平时候时较低,而在中等生产力水平时达到最大[15-16]。但我们发现这些研究仍存在一些不足:第一,大多数试验都是在人工草地上进行,这些人工草地面积较小,生物多样性较低,生态系统组成结构比天然草地要简单的多[12,17],并且这些多样性-生产力的试验常使用随机的物种组合,这些都与自然生态系统植物组成有很大的差异[18-19],因此得出的结果可能并不具有普遍性;第二,这些试验通常都是在相对较短的时间(通常为2~10年)内完成,其研究结果也许只能够反映生态系统较短暂或早期的演替动态[20-22],并不能确定多样性和生产力接近稳定或平衡时的状态[23];第三,对物种多样性-生产力关系的研究常集中在空间尺度,所得出的结果也都与空间尺度直接相关[2-3,5,24-25],却很少关注时间尺度上的变化[26-27],是否在时间尺度上的模式也与空间尺度上的模式一致?这些问题都需要进一步的研究和探讨。
生态系统演替模型可用来检验物种多样性-生产力关系在时间尺度上变化,可避免仅由早期演替引起的偏差[29],同时也能够考虑实际的物种丰富度,而不仅仅是起始的物种丰富度[30-31]。但至今只有少数几个试验研究检验了不同的演替阶段对物种多样性-生产力关系的影响[27,30,32-33]。一些数学模型已经表明,物种丰富度-生产力的关系会因演替阶段的不同而不同[34-35]。恢复演替试验表明,物种丰富度随时间的变化表现为“单峰”状增加,而生物量随时间呈线性或非线性增加[6,32,36]。Guo[26]的研究表明:在时间尺度上,演替早期,物种多样性和生物量均有增加,二者之间呈正相关关系;演替后期,物种生物量持续增加,但多样性开始减少,二者之间表现为负相关关系。Matthiessen[37]发现在演替早期物种丰富度和生物量间为正相关关系,而在演替后期均匀度和生物量之间表现为强的负相关关系。也有研究表明,随着演替的进行,草地经历连续的扰动后物种多样性-生产力关系可能表现为没有相关关系[38-39]。此外,生态系统始终是动态变化的,并且受到“生物互作”的强烈影响[40],演替前期的物种多度分布往往影响着演替后期的植物群落动态[41],深入了解物种多度的变化能使我们更清晰地理解生态系统的动态变化机制[42]。因此,研究长时间演替梯度上物种多样性、生产力的变化模式及二者的关系不仅能丰富植被演替相关理论,也可为生物多样性-生态系统功能的研究提供试验证据。本研究以宁夏云雾山典型草原生态系统为研究对象,基于40年的长期定位监测,研究物种组成、植物多度(总的植物个体密度)和地上生物量的变化趋势,旨在揭示长时间尺度上物种多样性与地上生物量的关系及其潜在机制,为草地生物多样性保护和区域畜牧业可持续发展提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区位于西北农林科技大学宁夏云雾山草原生态系统国家长期科研基地(106°24′~106°28′ E,36°13′~36°19′ N),该基地位于宁夏回族自治区固原市北部45 km处的宁夏云雾山国家级自然保护区内,海拔高度为1 800~2 150 m。该地属中温带半干旱大陆性季风气候,年均温7.01℃,7月最高气温22~25℃,1月最低气温—18~—15℃,全年0℃以上积温2 847~3 592℃,年平均日照时间为2 300~2 500 h,年均降水量为440 mm,其中有60%~75%的降水集中在6—9月,是地区水资源补给的主要来源。研究区土壤类型为山地灰褐土和黑垆土,pH值7.9~8.2;植被类型为暖温带典型草原,优势种和建群种主要有本氏针茅(Stipabungeana)、大针茅(Stipagrandis)、甘青针茅(Stipaprzewalskyi)、白莲蒿(Artemisiasacrorum)、百里香(Thymusmongolicus)和甘菊(Chrysanthemumlavandulifolium)等[43]。该草地在1982年封禁管理前属于自由放牧草地,放牧强度为重度放牧,载畜率为5~8只羊单位·hm-2。草地群落密度仅为3~8株·m-2,主要以蒿属植物(冷蒿(Artemisiafrigida)、茵陈蒿(Artemisiacapillaris)等)为主,群落高度为10~13 cm,覆盖度为20%~30%,地上生物量为750 kg· hm-2左右。草地封育初期,土壤有机质含量为(8.81±0.05) g·kg-1,土壤全氮含量为(0.42±0.04) g·kg-1。
1.2 试验设计
自1982年云雾山保护区成立以来,实行了严格的封山禁牧措施。同年4月,在围栏内选择相对平坦的天然草地(坡度约为5°)作为固定样地,随机设置30个20 m×30 m的小区,对每个小区进行编号并用于长期定位监测,小区之间至少相距50 m,小区四角用铁桩固定。每个小区分成两部分:一部分为20 m×12 m,用于植被调查和土壤采样;另一部分为20 m×18 m,用于植物个体采样。
1.3 研究方法
每年8月中旬生物量达到高峰时,在每个定位小区的20 m×12 m部分,随机选取1个1 m×1 m的样方进行群落学调查,为避免边缘效应,样方距边缘的距离大于0.5 m。调查内容包括每个物种植株高度、多度以及盖度。将样方中的所有植物按种分类,对地上部分进行齐地面刈割,按物种装入纸袋封袋编号,带回实验室在65℃条件下将物种地上部分烘干至恒重,称其干重。所有物种干重之和即为群落地上生物量。本研究利用物种丰富度(样方内所有物种的数目)和植物多度(样方内所有物种的个体总数)作为物种多样性的衡量指标[44]。物种丰富度是群落结构的数量化表征,而植物多度可在此基础上更进一步的对群落性质进行阐释,两者相辅相成。地上生物量可以近似代表地上净初级生产力[5]。
1.4 数据处理及分析
我们首先构建时间序列(Time series),通过局部加权回归(Loess)来确定物种丰富度、植物多度和地上生物量随时间变化的趋势并据此划分演替阶段。使用一般线性模型(GLM)检验物种丰富度-地上生物量、物种丰富度-植物多度和植物多度-地上生物量的关系[27],以上三组变量之间的关系中,前者作为预测变量,后者作为解释变量。使用赤池信息准则(Akaike’s information criterion,AIC)选择最优模型,即AIC值越小,拟合效果越好。此外,通过MOS (Mitchell-Olds and Shaw)检验来确定曲线峰值是否在实际观察范围之内,若峰值在实际观察范围之内,则物种丰富度-植物多度-地上生物量的关系呈典型的“单峰”状[5,45]。所有统计分析采用R-3.2.3完成,统计检验的概率显著性水平为P=0.05。
2 结果与分析
2.1 时间尺度上物种丰富度、地上生物量和植物多度的变化模式
在40年的演替过程中,共发现115种植物,分属29科,83属,其中物种数排在前5位的科是:禾本科(19)、菊科(19)、豆科(11)、蔷薇科(10)和唇形科(9),这5科植物约占植物总数的60%。物种丰富度的范围为3~28种·m-2,平均为18种·m-2。地上生物量的范围为30~690 g·m-2,平均为351 g·m-2。植物多度的范围为10~860株·m-2,平均为129株·m-2。
在时间尺度上,物种丰富度的变化模式为“急剧上升→相对稳定→急剧下降”地上生物量的变化模式为“急剧上升→平缓下降”。物种丰富度(图1A)和地上生物量(图1B) 随时间的变化整体表现为“单峰”状,植物多度(图1C)则随时间的变化整体呈上升趋势。
综合物种丰富度、地上生物量和植物多度的变化模式,将整个演替过程分为三个阶段,分别为:演替早期(1982—1993)、演替中期(1994—2011)和演替后期(2012—2021)(图1D)。
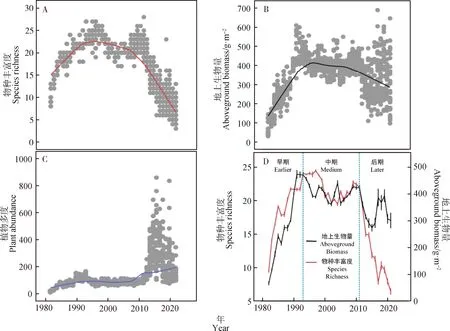
图1 物种丰富度、地上生物量以及植物多度在时间序列(1982—2021)上的变化趋势Fig.1 Changes in species richness, aboveground biomass and plant abundance from 1982 to 2021注:图1A,1B,1C分别为物种丰富度、地上生物量以及植物多度在时间序列上的变化趋势,1D为物种丰富度和地上生物量在三个演替阶段上的变化趋势Note:Fig.1A,1B and 1C show the changing trends of species richness,aboveground biomass and plant abundance in time series,respectively. Fig.1D shows the changing trends of species richness and biomass in three succession stages
2.2 时间尺度上物种丰富度和地上生物量的关系
我们发现物种丰富度和地上生物量的关系从演替早期(图2A,表1)到演替中期(图2B,表1)和后期(2C,表1)都表现为“单峰”关系,但随着演替的进行,决定系数(R2)逐渐降低,表明地上生物量对物种丰富度的影响在演替早期相对较大,在演替中后期相对较小。在整个时间尺度上,物种丰富度和地上生物量的关系表现为“单峰”关系(图2D,表1)。
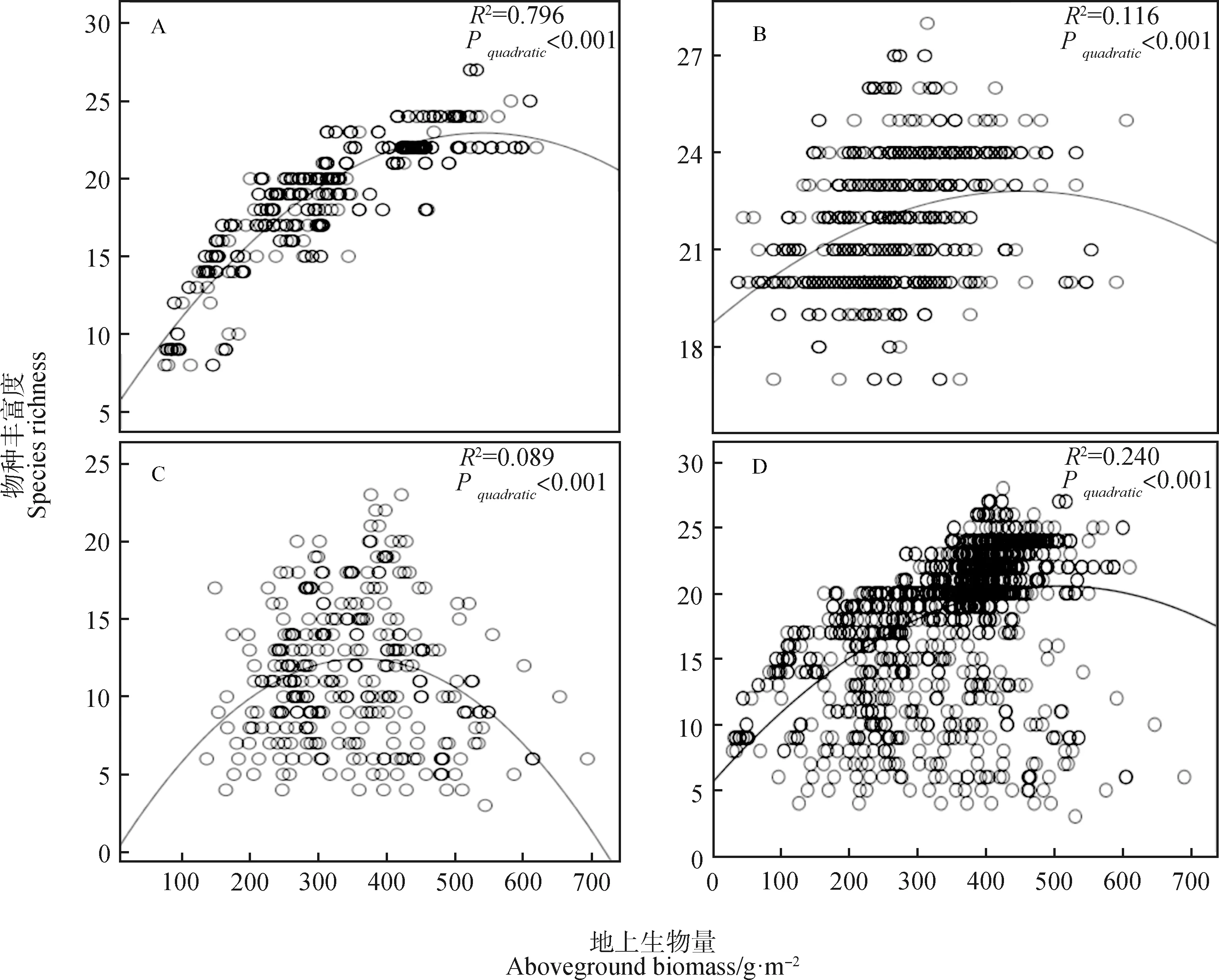
图2 不同演替阶段物种丰富度-地上生物量的关系Fig.2 Relationship between species richness and aboveground biomass during the different successional stages注:图2A,2B,2C分别为物种丰富度和地上生物量关系在演替早期、中期、后期的变化趋势,图2D为物种丰富度和生物量关系在整个演替上的变化趋势Note:Fig.2A,2B and 2C are the changing trend of the relationship between species richness and aboveground biomass in the early,middle and late stages of succession,respectively,Fig.2D is the changing trend of the relationship between species richness and aboveground biomass in the whole succession
2.3 时间尺度上植物多度和地上生物量的关系
植物多度和地上生物量在演替早期表现为正的线性相关关系(表1,图3A),在演替中期并没有相关关系(表1,图3B),而在演替后期表现为负的线性相关关系(表1,图3C)。以上结果表明在演替早期地上生物量有利于植物多度的增加,在演替后期地上生物量抑制植物多度的增加,而在演替中期,这种促进作用和抑制作用可能相互抵消,表现为没有相关关系。在整个时间尺度上,植物多度和地上生物量的关系表现为“单峰”关系。
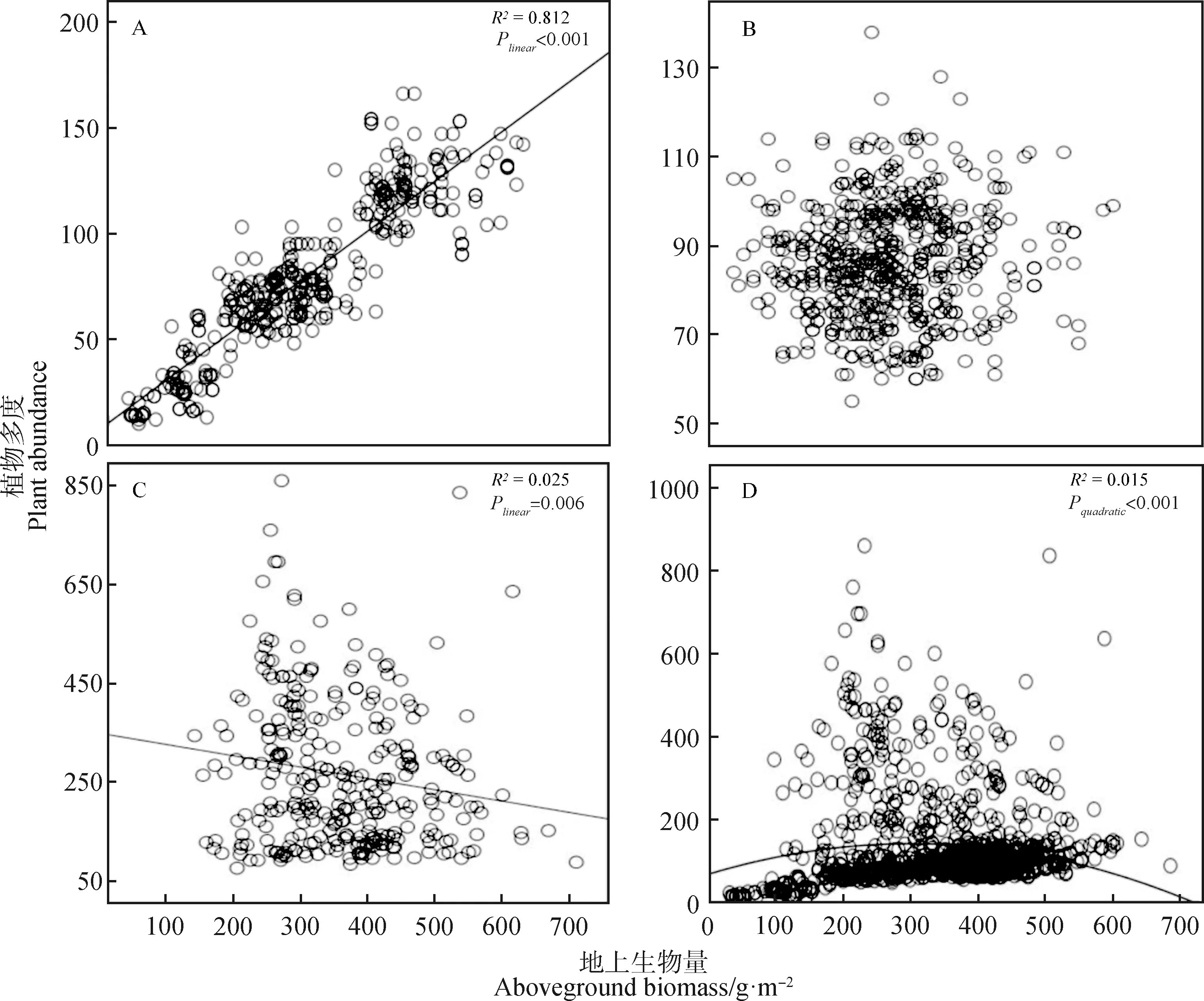
图3 不同演替阶段植物多度-地上生物量的关系Fig.3 Relationship between plant abundance and aboveground biomass during the different successional stages注:图3A,3B,3C分别为植物多度和地上生物量的关系在演替早期、中期、后期的变化趋势,3D为植物多度和地上生物量的关系在整个演替上的变化趋势Note:Fig.3A,3B and 3C show the changing trend of the relationship between plant abundance and aboveground biomass in early,middle and late successional stages,respectively,Fig.3D shows the changing trend of the relationship between plant abundance and aboveground biomass in the whole successional stage
2.4 时间尺度上物种丰富度和植物多度的关系
不同于物种丰富度和地上生物量的关系,物种丰富度和植物多度关系在演替早期和演替后期表现为“单峰”关系,但在演替早期植物多度对物种丰富度的影响要大于演替后期植物多度对物种丰富度的影响(表1,图4A和图4C)。在演替中期,物种丰富度和植物多度之间并没有相关关系(表1,图4B)。在整个时间尺度上,物种丰富度和植物多度的关系表现为“倒单峰”关系(表1,图4D)。与地上生物量对物种丰富度的影响相比,植物多度对物种丰富度的影响相对较小(表1)。
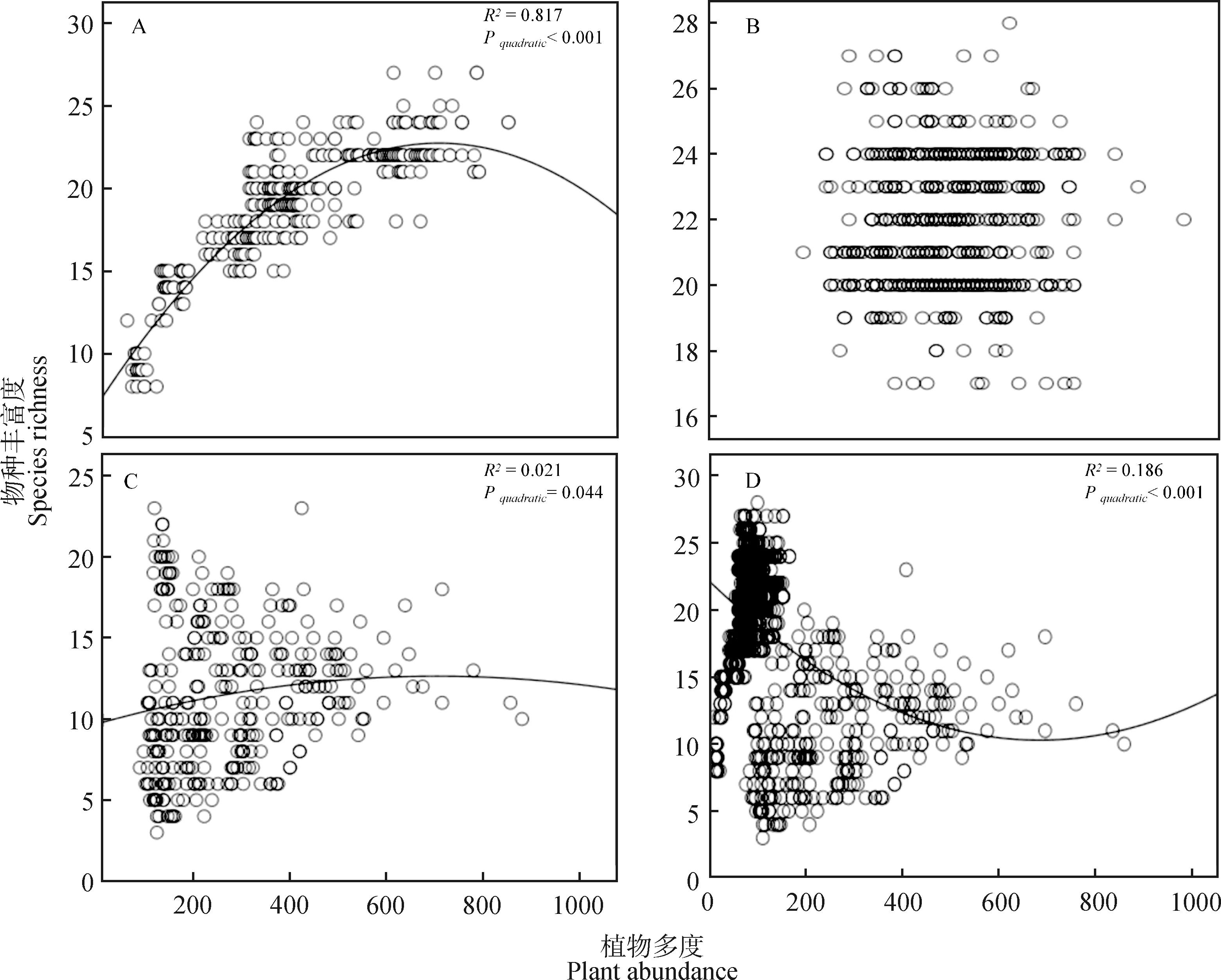
图4 不同演替阶段物种丰富度-植物多度的关系Fig.4 Relationship between species richness and plant abundance during the different successional stages注:图4A,4B,4C分别为物种丰富度和植物多度的关系在演替早期、中期、后期的变化趋势,4D为物种丰富度和植物多度的关系在整个演替上的变化趋势Note:Fig.4A,4B and 4C show the changing trend of the relationship between species richness and plant abundance in the early,middle and late succession stages,respectively,and Fig.4D shows the changing trend of the relationship between species richness and plant abundance in the whole succession

表1 物种丰富度-植物多度-地上生物量之间的回归分析Table 1 Summary of regression analyses of relationships among species richness,plant abundance and aboveground biomass during the different successional stages
3 讨论
3.1 演替过程中物种丰富度、植物多度和地上生物量的变化模式
与其它时间序列的研究类似[6,15,36],本研究结果表明:自1982年退化草地实施封禁管理后,物种丰富度和植物地上生物量迅速增加。1993年,物种丰富度和生物量都达到峰值,并维持相对稳定状态,到了演替后期(2011年后),随着生物量累积速度的降低和物种间竞争的加剧,物种丰富度显著降低,地上生物量则表现为平缓下降的趋势,但植物多度会随着演替的进行表现为不断增加的趋势。因此,40年的演替结果表明物种丰富度和地上生物量在时间尺度上表现为“单峰”状(图1),即在演替前期迅速增加,在演替中期达到峰值并维持相对稳定,在演替后期降低迅速。Bonet等人[36]根据一个60年时间序列的研究发现,物种丰富度在前10年迅速增加并于第18年达到峰值,随后逐渐下降。这种现象可能原因是在演替早期,群落主要以生长迅速、扩散能力强的一年生植物和短寿命的多年生杂草为主(r对策植物)[46],随着演替的进行,一年生植物和短寿命多年生杂草无法与长寿命、高抵抗力的多年生禾草植物(K对策植物)竞争,在演替后期被取代[47]。即在演替早期,物种的扩散速度通常主导群落组成结构的变化[48],导致物种丰富度和植物多度的迅速增加[49]。此后,随着群落演替的继续,物种的获得和丧失可能相互抵消,群落将趋于相对平衡状态[12]。到演替后期,随着资源(光照、水分和养分等)竞争的加剧,物种的丧失速度也随之加快,进而导致物种丰富度迅速降低[33,50]。此外,在演替过程中,物种多样性的增加会增加资源的利用效率,进而增强生态位互补效应,导致地上生物量的增加[51-52]。然而,在演替的中后期,随着群落中凋落物的累积,地上生物量也会逐渐降低[53]。因此,对于物种丰富度、植物多度和地上生物量随演替时间变化的模式的理解可能有助于提高我们对植被恢复动态的理解,并为保护和恢复提供管理指导[54]。
3.2 时间尺度上物种丰富度和地上生物量的关系
我们发现,物种丰富度-地上生物量在不同的演替阶段和整个时间尺度上均表现为“单峰”关系,但地上生物量对物种丰富度的影响程度在演替早期大于演替中后期。越来越多的研究也表明,物种多样性对地上生产力的影响在时间上是动态的[10,12,33-34,55]。Guo[26]提出的演替模型表明,在演替的早期阶段,物种多样性和地上生产力是正相关关系,而负相关关系和“单峰”关系则可能出现在不同的演替阶段。此外,无论短期或长期试验,自然群落或人工合成系统,大多数草地试验表明物种多样性和生产力之间的关系是对数-线性或曲线-线性关系[6,9-10,56-62]。但与这些试验研究结果不同,我们的结果表明物种丰富度和地上生物量间存在显著的“单峰”关系,特别是在演替早期,地上生产力对物种丰富度有较强的影响,这可能是因为在演替早期,有较丰富的“空”的生态位(时间和空间),物种受环境的限制相对较小,物种间的竞争作用较弱,而扩散等随机过程决定了演替前期的群落动态[27],但随着演替的进行,地上生产力对物种多样性的影响可能会因资源的限制而逐渐减弱。而其它一些研究则表明物种多样性和生产力之间的关系会随着时间的推移而不断增强[10,12]。此外,物种多样性-生产力的关系也会因不同的生态系统类型而发生变化,例如,在热带雨林中发现[33],在演替早期,物种多样性和生产力之间是正的线性关系,而在演替后期则表现为没有相关关系。总之,我们的长时间序列数据表明,生产力对物种多样性影响的大小或方向会因时间尺度的不同而有所不同。
3.3 时间尺度上物种丰富度-植物多度及植物多度-地上生物量的关系
群落中植物多度的变化可以很好地解释物种丰富度的变化[54]。理论预测,在演替前期,由于此时植被较稀疏,随着生产力的增加,植物多度也会逐渐增加,当生产力进一步增加时,植物多度可能会下降;在演替中后期,伴随着生产力的增加,植物个体大小也会相应增加,而个体大小的增加会导致植物多度的降低,引起自疏过程的发生[54],自疏的关键不是具有某些特性的物种排除了其他物种,而是所有的物种都具有同等的消失几率。因此,随着演替的进行,地上生物量逐渐增大,起始多度较少的物种更容易从群落中消失,从而引起物种多样性的降低。一些研究表明,植物多度变化最多可使多样性降低42.9%[63]。本研究结果表明,在不同的演替阶段物种丰富度-植物多度的关系表现不同,但总体上表现为“倒单峰”关系,这与几个人工控制试验的结果一致[12-17]。此外,有研究表明,演替过程中植物多度的增大与群落中优势种群的扩张直接相关[64]。在演替早期,植物的扩散能力较强,植物多度逐渐增大,生物量也随之增大,但在演替后期,因竞争等确定性过程占优势,物种多样性降低,物种的多度也随之降低,因此,与物种丰富度-地上生物量的关系一致,植物多度和地上生物量之间也表现为“单峰”关系,这与宋晓谕等[65]、吴建波等人[66]的研究结果相似,并且植物多度和地上植物生物量的关系会随着时间的推移而增强。
物种间正相互作用(促进)和竞争的相对重要性可能是引起时间尺度上物种丰富度-植物多度-地上生物量关系发生变化的主要原因[42]。在演替早期,生境中可利用资源较多,竞争较少,扩散、正相互作用和环境忍耐等因素限制了物种丰富度的变化[46,50];同时随着植物多度的不断增加,特别是繁殖能力强的杂草迅速扩张、生长和大量繁殖,地上生产力也逐渐提高。在演替后期,受环境承载力的限制,地上生产力开始下降[67],竞争在此阶段成为主导因子[34,50],加之地面凋落物的不断累积,植物正常生长发育和繁殖更新受到阻碍[68],从而引起物种丰富度降低,群落组成结构趋于简单[69]。至于演替中期植物多度与二者的关系均不显著,可能的原因是随着群落结构复杂化,植物生长受到生境中多种因素的综合影响[70],单以植物多度变化无法进行解释;还可能是两者促进作用和抑制作用相互抵消,从而表现为中性作用[52,71]。
根据植物多度与丰富度相似的变化趋势及其与地上生产力的关系,我们认为可以用Stevens和Carson[54]提出的集合水平自疏假说(Assemblage-level thinning hypothesis)以及Newman[72]的光竞争假说(Light competition hypothesis)共同解释二者关系的内在机制[73],即物种多样性的变化与植物多度直接相关,且随着种群密度的增大,竞争从地下资源竞争转变为地上的光竞争,在高生产力的生境中,大的或高的物种能够提前占据光资源(有更大的竞争优势),进而排除光竞争能力弱的物种。未来的研究还需结合其它生物因素和非生物因素进一步明确二者的关系及其可能的机制。
4 结论
总的来说,草地生态系统的演替过程是动态变化的,物种丰富度和地上生物量在演替前期迅速增加,在演替中期达到峰值并维持相对稳定,在演替后期迅速降低。物种多样性(丰富度和多度)与地上生物量的关系会因演替阶段的不同而发生变化,在整个时间尺度上表现为“单峰”关系。因此,详细了解物种多样性与生产力之间的动态关系是生物多样性保育和生态系统健康管理的关键。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
农业工程学报(2022年10期)2022-08-22
中国音乐学(2022年2期)2022-08-10
现代园艺(2022年6期)2022-04-08
科学导报(2021年64期)2021-09-29
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
吉林农业(2018年8期)2018-05-01
中学生理科应试(2017年6期)2017-09-27
