
Metabolomics provide insights into the endogenous mechanism of strobilation in the scyphozoan jellyf ish Rhopilema esculentum*
2022-03-05 09:36JianlongGEXiaohuiCHENChanglinLIUJieTANLiBIANLifeiCHENSiqingCHEN
Jianlong GE , Xiaohui CHEN , Changlin LIU , Jie TAN , Li BIAN , Lifei CHEN ,Siqing CHEN ,**
1 Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071, China
2 Yancheng Huayi Aquatic Products Co. Ltd., Yancheng 224300, China
Abstract Strobilation is a well-organized metamorphosis process in scyphozoan jellyf ish, through which the benthic polyp develops into the pelagic medusa. So far, knowledge on the molecular mechanisms behind the drastic morphological and physiological changes remains limited. In the present study, the metabolic prof iles in polyp and strobila stages of jellyf ish Rhopilema esculentum were examined using ultra-performance liquid chromatography coupled to a mass spectrometer. A total of 3 071 metabolites with putative annotation were detected, of which 167 were identif ied as diff erential metabolites between the polyp and strobila. Among the metabolites, 31 signif icantly decreased and 136 signif icantly increased in abundance in the strobila. Thyroxin, one of the previously proposed strobilation inducer of Aurelia, was not detected in this study. The indole-containing compounds are known for triggering strobilation in Discomedusae. Two indole derivatives and an indole-isomer containing metabolite were detected among the most signif icantly increased metabolites, which may be potential endogenous molecules for strobilation initiation. The membrane-associated phospholipids and the inf lammation and oxidative stress-related eicosanoids were found to have signif icant changes. Although the functions of these metabolites in strobilation are not yet completely known, they provide some clues to the induction of the process of strobilation and metabolic responses that take place during strobilation.
Keyword: Rhopilema esculentum; scyphozoan jellyf ish; strobilation; metabolomics
1 INTRODUCTION
Scyphozoan jellyf ish are conspicuous members of the ocean ecosystem and are notorious more recently for their interference with human and industrial activities (Purcell et al., 2007; Dong et al., 2010).Their life cycles are among the most complex of any non-parasitic animal, which consists of two morphologically disparate generations with three well-def ined life stages—planula, polyp, and medusa(Helm, 2018). In this life cycle, the transition from polyp to medusa is achieved through an asexual reproduction process called “strobilation”, which has gained considerable interest in the past century(Berrill, 1949; Spangenberg, 1967, 1974; Fuchs et al.,2014; Gold et al., 2015; Helm and Dunn, 2017).
Strobilation starts just below the polyp’s ring of tentacles and proceeds down the body like a wave.The process becomes visible in a successive formation of constrictions. Finally, the discs of the tissue between each constriction develop into ephyrae.During this transformation, two processes take place and they are well known to play a key role in the formation of various higher metazoa: segmentation and metamorphosis (Kroiher et al., 2000). Helm(2018) recently provided a comprehensive and detailed review of strobilation processes. Temperature shift, endogenous rhythms of the animal and chemical cues from the environment could trigger strobilation(Prieto et al., 2010; Kuniyoshi et al., 2012; Fuchs et al., 2014). Several compounds have been reported for the chemical induction of strobilation in laboratory,such as indomethacin (Kuniyoshi et al., 2012;Cabrales-Arellano et al., 2017; Helm and Dunn,2017), H2O2(Berking et al., 2005), iodine and thyroxine (Spangenberg, 1967, 1974; Olmon and Webb, 1974), retinol, 9-cis-retinoic acid and the indole compounds: 5-methoxy-2-methyl-3-indoleacetic acid, 5-methoxyindole-2-carboxylic acid, 2-methylindole, and 5-methoxy-2-methylindole(Fuchs et al., 2014). Nevertheless, the endogenous molecular cascades in strobilation initiation remain an enigma in scyphozoans.
Several studies have attempted to characterize the endogenous compounds associated with strobilation.In the moon jellyf ishAureliaaurita, a compound that is similar to thyroxine was detected in the surrounding medium of strobilating polyps (Spangenberg, 1971).Similarly, two compounds, tentatively identif ied as iodotyrosines and thyroxine, were identif ied in a Pelagiidae speciesChrysaorachesapeakei(Black and Webb, 1973). Loeb (1974a) also isolated a factor named ‘neck-inducing factor’ (NIF) from the surrounding water ofC.chesapeakeipolyps after exposing them to strobilation induction clues. The NIF is a peptide of 1 650 Da, while its molecular structure and amino acid composition are still unknown (Loeb,1974b). Recently, research on strobilation has focused primarily on transcriptomics, providing new insights on the strobilation mechanism at a molecular level(Fuchs et al., 2014; Brekhman et al., 2015). Fuchs et al. (2014) particularly demonstrated that retinoic acid signaling seems conserved in controlling the transition of polyps to free-swimming jellyf ish. In addition, they deduced that one of the upregulated proteins upon temperature changes may function as temperaturesensitive ‘‘timer’’ and encodes the precursor of the strobilation hormone ofAurelia. Meanwhile,Brekhman et al. (2015) presented transcriptome prof iles covering the entire developmental stages ofAureliaand highlighted the potential key roles of some stage-specif ic transcription factors in strobilation.Metabolomics, however, is essential in revealing the mechanisms involved in phenotypic development and in physiological responses that initiate complex developmental processes. To our knowledge, no metabolomics study has yet been conducted on scyphozoan strobilation.
Rhopilemaesculentum, belonging to the Class Scyphozoa, Order Rhizostomeae, and Family Rhopilema, is widely distributed in the northwest Pacif ic Ocean. It is one of the most commercially important edible jellyf ish with high nutritional and medicinal value (You et al., 2007). For these reasons,it has been an important aquaculture species and pond cultured in coastal areas of China for several decades.As in most scyphozoan species,R.esculentumhas a complex life cycle, alternating between a sessile polyp and free-swimming medusa (Ding and Chen,1981). The polyps under low winter temperatures(2-10 °C) could be induced to strobilate when temperatures were raised to 18-27 °C (Chen and Ding, 1983). Our previous transcriptomics study pointed out the role of transcription factors in strobilation and produced a list of novel candidate genes (Ge et al., 2018). Nonetheless, our results were inconclusive particularly on the endogenous mechanisms of strobilation initiation. In the present study, we conducted a comparative metabolomics analysis between the polyp and strobila stages ofR.esculentum. A set of signif icantly diff erential metabolites were identif ied, which included potential metabolites for strobilation initiation or progression.These f indings will be useful for a more comprehensive understanding ofR.esculentumstrobilation.
2 MATERIAL AND METHOD
2.1 Sample collection
The sessile polyps ofR.esculentumwere obtained from Yancheng Huayi Aquatic Products Company(120°23′E, 33°59′N) in Sheyang, Jiangsu Province of China. The producing and culturing processes of polyps were as previously described (Ge et al., 2018).Culturing temperature was raised from 10 °C to 18 °C to induce strobilation. The early strobila, polyp with a clear constriction under the tentacle or a few transverse segments, appeared in one week. Fifty polyps (reared under 10 °C) and 50 early strobila polyps were collected to serve as samples for the two stages. The samples were rinsed thoroughly with f iltered sea water, frozen immediately in liquid nitrogen, and then stored at -80 °C for the following analysis.
2.2 Sample preparation for metabolomics
Thirty milligrams of each sample was added to 400 μL of 80% methanol and 20 μL of internal standard(2-chloro-L-phenylalanine in methanol, 0.03 mg/mL;Lyso PC17:0 in methanol, 0.01 mg/mL). The samples were precooled at -20 °C for 2 min and then ground at 60 Hz for 2 min. The metabolites were ultrasonic extracted for 30 min and centrifuged at 13 000 r/min for 10 min at 4 °C. The supernatants (200 μL) from each tube were collected using crystal syringes, f iltered through 0.22-μm microf ilters and transferred to liquid chromatography (LC) vials. The vials were stored at-80 °C for the following analysis. Quality control(QC) samples were prepared by mixing aliquots of all the samples, and were pooled and analyzed in the same manner as the analytical samples.
2.3 LC-MS analysis
A Dionex Ultimate 3000 RS ultrahigh-performance liquid chromatography (UHPLC) system, f itted with Q-Exactive quadrupole-Orbitrap mass spectrometer and equipped with heated electrospray ionization(ESI) source (Thermo Fisher Scientif ic, Waltham,MA, USA), was used to analyze the metabolic prof ile in both ESI positive and negative ion modes. The ACQUITY UPLC BEH C18 columns (1.7 μm,2.1 mm×100 mm) were employed in both positive and negative modes. The procedures and detection parameters for LC-MS were as described in Yang et al. (2018) with a few modif ications. Brief ly, the binary separation elution was 0.1% formic acid in water (A)and 0.1% formic acid in acetonitrile (B). The separation gradient was set as follow: 5%-20% B for 0-2 min, 20%-60% B for 2-4 min, 60%-100% B for 4-11 min. The composition was held at 100% B for 2 min, then 13-13.5 min, 100% to 5% B, and 13.5-14.5 min holding at 5% B. The main parameters were listed as follows: the f low rate, 0.4 mL/min; the column temperature, 45 °C; the injection volume,5 μL. The mass range was from 66.7 to 1 000.5m/z.The resolution was set at 70 000 and 35 000 for the full MS scans and HCD MS/MS scans, respectively.The collision energy was set at 10, 20, and 40 eV. The QCs were injected at regular intervals (every 10 samples) throughout the analytical run to assess the repeatability of the data.
2.4 Data processing and statistical analysis
The acquired raw data were processed by the Progqenesis QI software (Waters Corporation,Milford, USA) for baseline f iltering, peak identif ication, integration, retention time (RT)correction, peak alignment, and normalization. The parameters for precursor tolerance, fragment tolerance, fragment ion threshold were set as 5×10-6,10×10-6, and 5%, respectively. Internal standard detection parameters were excluded for peak RT alignment and isotopic peaks were excluded from the analysis. The level of noise elimination was 10.00,and minimum intensity was set at 15% of the base peak intensity. A f ile containing datasets such asm/z,peak RT, and peak intensities was obtained. The RTm/zpairs were used as identif iers for each ion. The resulting matrix was further reduced by removing any peak with missing value (ion intensity=0) in more than 60% of the samples. The METLIN (https://metlin.scripps.edu/), Lipidmaps (http://www.lipidmaps.org/), and Human Metabolome Database(HMDB) (http://www.hmdb.ca/) were used for qualitative metabolite analysis and alignment. The qualitative results were f iltered according to its score and the assigned metabolites with score less than 30(maximum score of 60) were deleted. The data quality(reproducibility) was assessed by an internal standard.
A dataset integrating the positive and negative data were imported into the SIMCA software package(Version 14.0) for multivariate statistical analysis.Based on the relative abundances of the compounds,principal component analysis (PCA) was conducted to observe the cluster mode of all of the samples and the repeatability of the QC samples. Pairwise orthogonal partial least-squares discriminant analysis (OPLSDA) was performed to improve the discrimination between groups and to screen diff erential metabolites.Variable importance in the projection (VIP) ranks the overall contribution of each variable to the OPLS-DA model and variables with VIP>1 were considered relevant for group discrimination (Yin et al., 2020). To avoid OPLS-DA analysis overf itting, the default 7-round cross-validation and permutation testing (200 times) was applied. The diff erential metabolites were selected based on the combination of VIP values obtained from the OPLS-DA model. The p values from a two-tailed Student’st-test on the normalized peak areas, where metabolites with VIP>1.0 andP<0.05, were considered as signif icantly diff erent metabolites. Finally, those diff erential metabolites were mapped to metabolic pathways using the Kyoto Encyclopedia of Genes and Genomes (KEGG)database. The model cnidarian sea anemone,Nematostellavectensis, was selected as reference species for KEGG mapping. Those with false discovery rate (FDR) correctedP<0.05 were def ined as signif icantly enriched pathways.
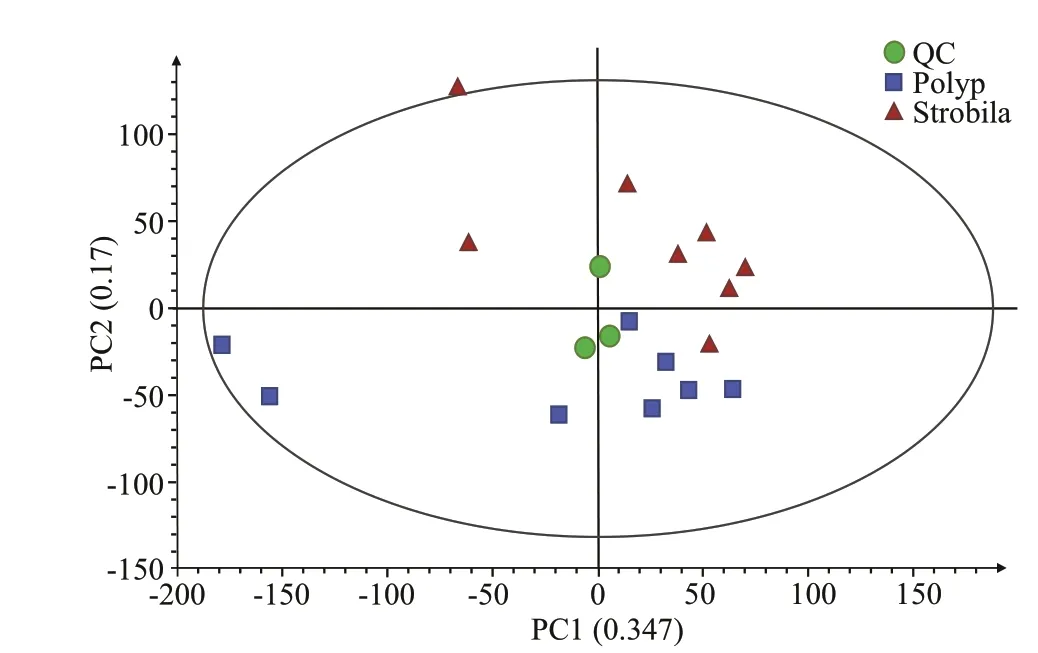
Fig.1 Principal component analysis (PCA) score plot of the metabolic prof iles of the polyp and early strobila o f Rhopilema esculentum
3 RESULT AND DISCUSSION
3.1 General characteristics of LC-MS analysis
The representative base peak intensity (BPI)chromatograms under positive and negative ion modes are shown in Supplementary Figs.S1 & S2,respectively. A total of 7 195 and 6 273 metabolomic features (m/z) were detected in positive ion and negative ion modes, respectively, and from which the internal standards, pseudo-positive peaks, and redundancies were excluded. Of these metabolite features, 3 071 were assigned putative IDs in the METLIN, HMDB, and Lipidmaps databases. The full list of those putatively identif ied metabolite mass features and concentration information are provided in Supplementary Table S1. The unsupervised principle component analysis (PCA) shows that the three QC samples cluster closely indicating the stability and repeatability of LC-MS. The score plots also show that the samples representing the two stages were apparently separated from each other and that there were no signif icant outliers (Fig.1).
3.2 Diff erential metabolites between polyp and early strobila
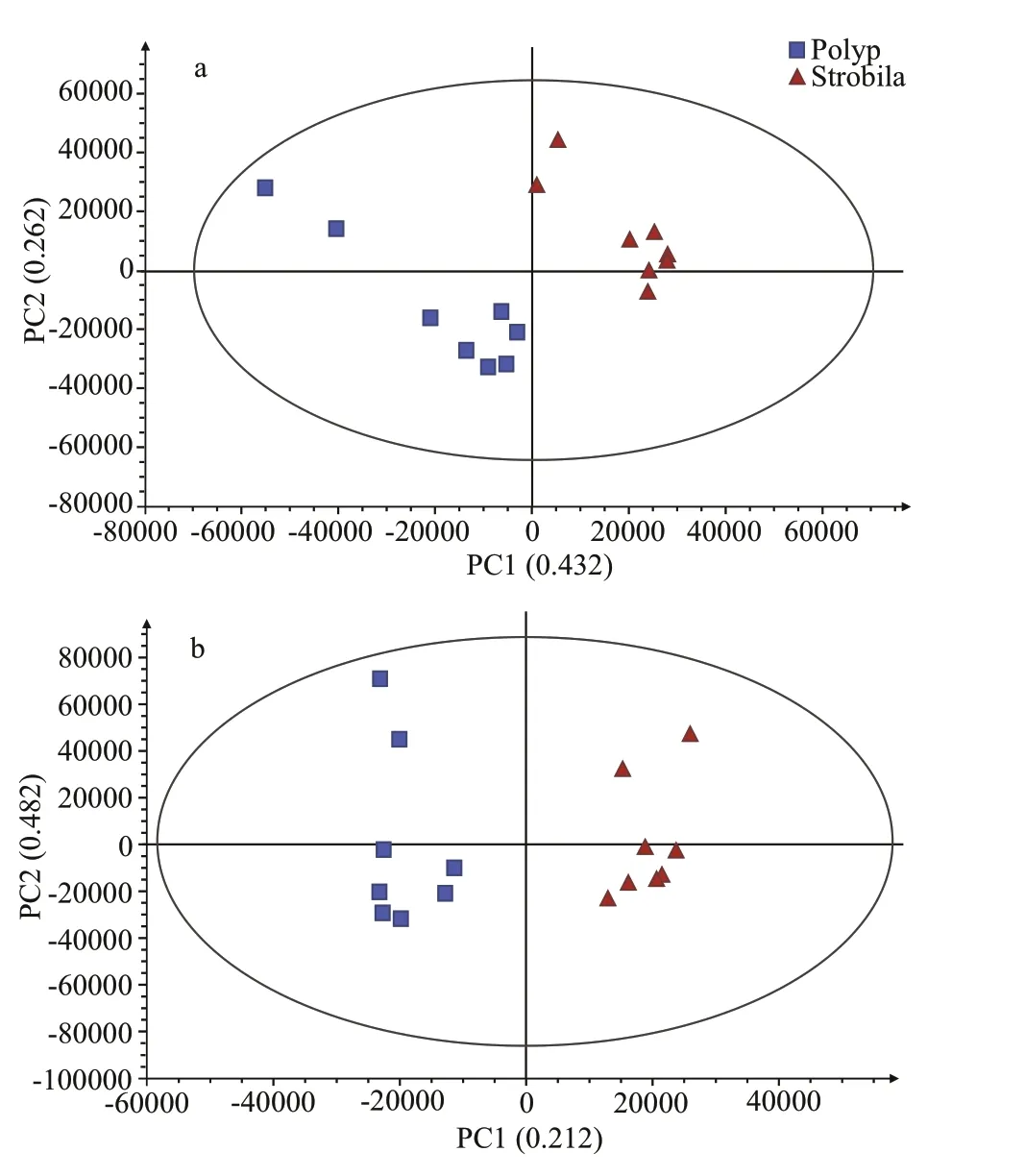
Fig.2 (Orthogonal) partial least squares-discriminant analysis ((O)PLS-DA) score plot of the metabolic prof iles of the polyp and early strobila of Rhopilema esculentum
(O)PLS-DA was then conducted to investigate the detailed metabolic diff erences between the polyp and early strobila. The score plots showed that the metabolic prof iles were signif icantly diff erent between the two groups (Fig.2). Cross-validation was conducted to avoid over-f itting by the OPLS-DA (Supplementary Fig.S3) and the resultingQ2value was less than 0,which indicated that the OPLS-DA model for this study was robust. In addition, the results of univariatetStudent’s analysis and fold change analysis were shown in a volcano plot (Supplementary Fig.S4).These were combined to determine the signif icantly diff erent metabolites. Finally, 149 metabolites, which displayed signif icant changes in abundances between the polyp and early strobila, were identif ied as diff erential metabolites under the criteria of VIP>1 andP<0.05 (Supplementary Table S2). Among them,125 metabolites signif icantly increased and 24 metabolites signif icantly decreased in the early strobila. Besides the above 149 diff erential metabolites,another 18 metabolites were also identif ied as candidate diff erential metabolites under the criteria of FC>4 or<0.25, VIP<1 andP<0.05 (Supplementary Table S3), of which the abundances of 11 metabolites signif icantly increased while 7 metabolites decreased in the early strobila. Among these 18 metabolites, 3 metabolites (samin, N′-hydroxyneosaxitoxin and barbituric acid) were prominently present in the strobila, while they were in extremely small amounts in polyps (Supplementary Table S3).
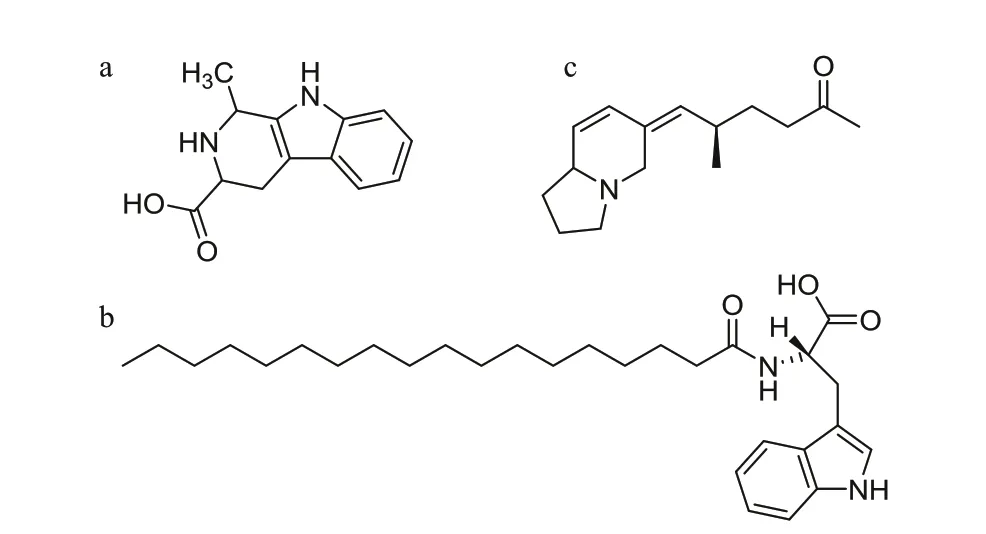
Fig.3 The structural formula of potential endogenous molecules for strobilation initiation of R. esculentum
Strobilation is a metamorphosis process involving tight regulation and coordination of each tissue as they undergo cellular reprogramming, proliferation,and apoptosis. For example, it was reported that polyp tentacles were resorbed and new medusa tentacles developed separately during strobilation(Spangenberg, 1991; Gold et al., 2015). Similar events occur for ephyra musculature, which forms de novo during strobilation (Helm et al., 2015).According to Berrill (1949), the grooves undergo cellular proliferation and heightening in both the epidermis and gastrodermis, and the discs of the tissue between each furrow are subject to cell proliferation in the ecto- and endoderm (Helm, 2018). Meanwhile,the ephyra rhopalial nervous system may be partially remodelled from the polyp nervous system (Schwab,1977). The putative identif ication of mass features provides some insights into endogenous molecular cascades under strobilation.
Thyroxin was considered necessary in strobilation in the two species,A.auritaandC.chesapeakei(Spangenberg, 1967, 1971; Black and Webb, 1973).In the present study, only thyroxin sulfate (metabolite of thyroxin) was detected in all samples. There was no signif icant diff erence between polyp and early strobila concentrations. The result indicates that thyroxin is not a common or key regulator of strobilation in scyphozoans. Consistent with our result, Olmon and Webb (1974) found that inorganic iodide rather than organically bound iodine was the eff ective factor in the initiation of strobilation inA.aurita. In contrast, thyroxine only caused strobilation occasionally inCassiopeaxamachana(Cabrales-Arellano et al., 2017).
The indole-containing compounds appear conserved for triggering strobilation in all Discomedusae (Kuniyoshi et al., 2012; Cabrales-Arellano et al., 2017; Helm and Dunn, 2017). They were proposed to activate an endogenous pathway leading to metamorphosis (Kuniyoshi et al., 2012) or to resemble the pharmacophore of the natural occurring inducer (Fuchs et al., 2014). However, it is still unknown whether there are endogenous indole or indole derivative compounds produced by polyps that could trigger metamorphosis. In the present study,two indole derivatives and an indole-isomer containing metabolite were detected among the most signif icantly increased metabolites. They were annotated as 1,2,3,4-Tetrahydro-1-methyl-betacarboline-3-carboxylic acid (HMDB0037942),N-stearoyl tryptophan (LMFA08020097), and (5R)-5-methyl-6-((Z)-1,2,3,8a-tetrahydroindolizin-6(5H)-ylidene)-2-hexanone (METLIN no. 263735),respectively. The structural formulae of those metabolites are shown in Fig.3. These compounds could function as potential endogenous molecules for strobilation initiation ofR.esculentum. Further research on the inf luence of those compounds on the metamorphosis ofR.esculentumand other scyphozoans is of interest.
In the present study, we found that the membraneassociated phospholipids (phosphatidylcholines, PCs;phosphatidylethanolamines, PEs; phosphatidylserines,PSs; phosphatidylinositols, PIs; and phosphatidylglycerols, PGs) were among the most signif icantly changed metabolites in abundances. A total of 26 phospholipids were detected which accounted for the largest proportion (17.4%) of the total identif ied diff erential metabolites (Table 1). The predominance of phospholipids among metabolites was also observed in the tadpole to froglet metamorphosis (Ichu et al., 2014; Luehr et al., 2018).Those results indicate that phospholipids may carry out some important functions during metamorphosis,while, their roles in those complex processes are still not understood (Luehr et al., 2018). It was proposed that the up- or down-regulation of these phospholipids are likely correlated with the switching of cell types in the tissues that are being reprogrammed, as diff erent cell types have diff erent requirements for membrane f luidity and structure (Luehr et al., 2018).
Eicosanoids were another important group, and nine metabolites were detected in the present study(Table 2). Three metabolites including PGD2 decreased and seven metabolites, including 5-HETE,Leukotriene D4, and Leukotriene F4, increased during the early strobilation. This particular result seems to be consistent with the suggestions that oxidative stress and reactive oxygen species (ROS)may be involved in strobilation induction (Herrmann et al., 2004; Berking et al., 2005). Eicosanoids,including prostaglandins (PG), leukotrienes (LT) and lipoxins (LXs), act as signaling molecules that play a crucial role in modulating inf lammatory conditions(Harizi et al., 2008). Similar to eicosanoids, ROS are also important inf lammatory mediators that are extensively involved in the innate inf lammatory responses (Bryan et al., 2012; Esser et al., 2012;Mittal et al., 2014; Rahmati et al., 2016). The inf lammation conditions could most likely lead to oxidative stress and vice-versa, which is consistent with the notion that the interactional oxidative stress and inf lammation play some roles in triggering strobilation or its progression.
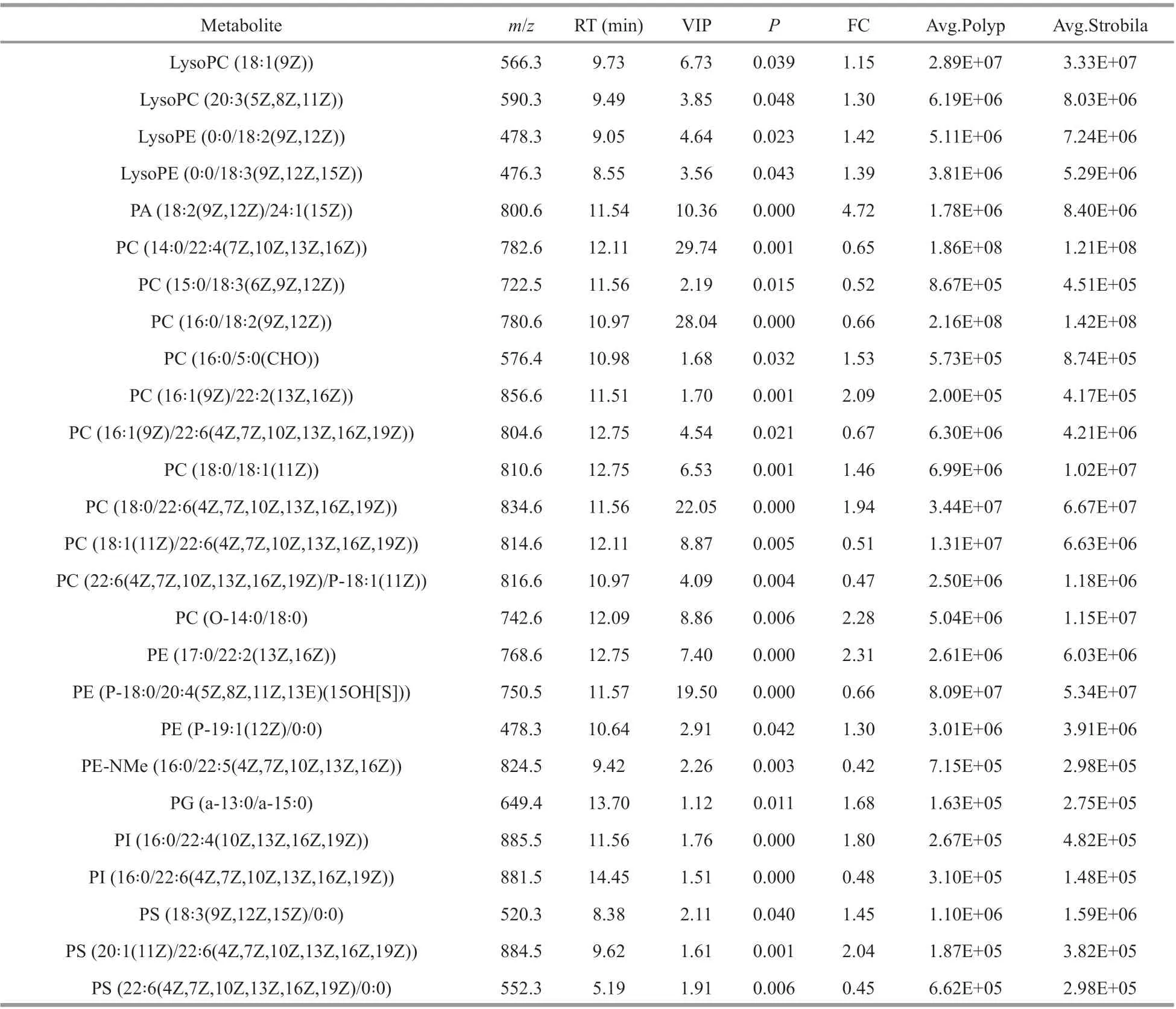
Table 1 Phospholipids that signif icantly changed ( P <0.05) in average abundances in the polyp and early strobila stages of R. esculentum
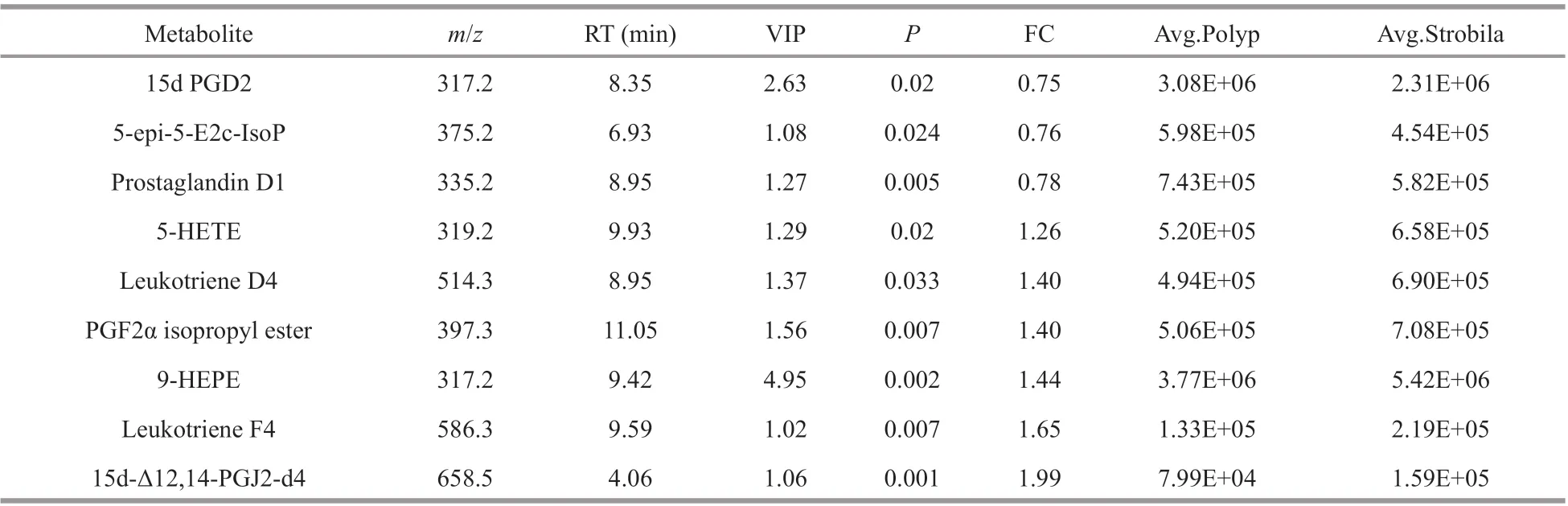
Table 2 Eicosanoids that signif icantly changed ( P <0.05) in average abundances in the polyp and early strobila stages of R. esculentum
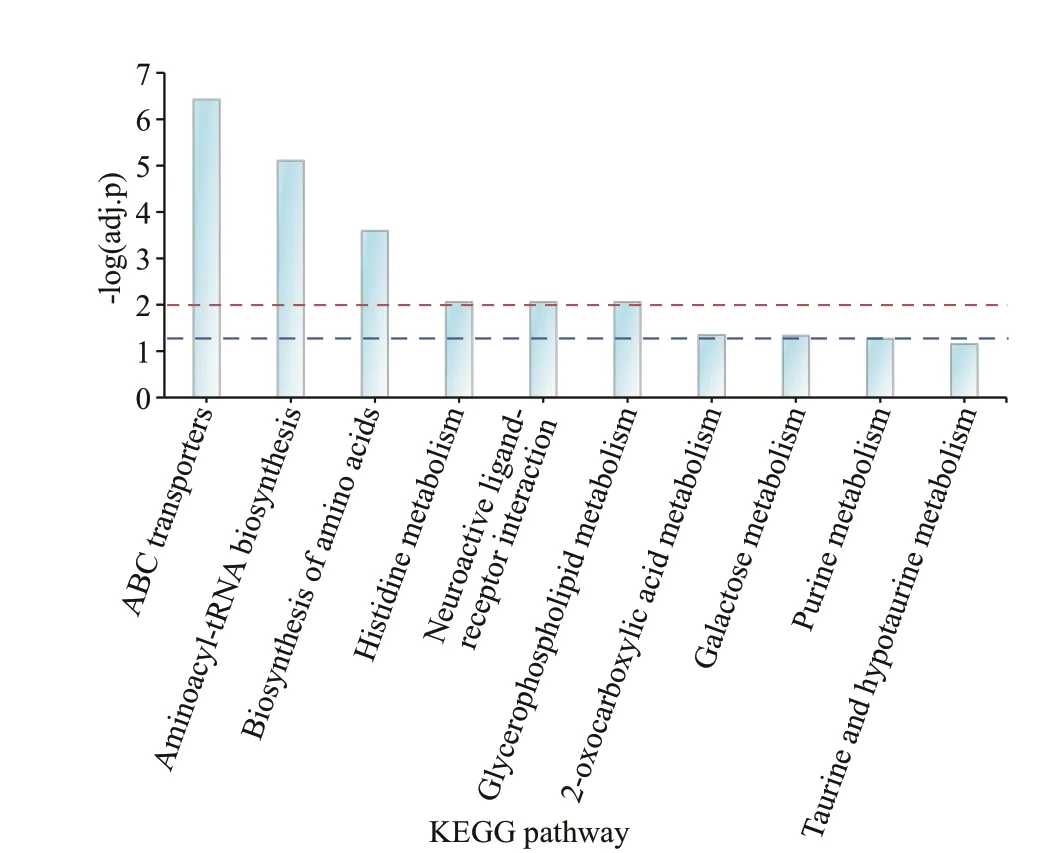
Fig.4 KEGG pathway enrichment with signif icantly changed metabolites
Berking et al. (2005) has proposed a model that points to the role of an oxidant defense system in strobilation ofAureliaaurita, namely, there was a feedback loop among ROS, iodide, tyrosine, and unknown strobilation control substance (SCS) to regulate the induction of strobilation. Polyps could always produce iodine from the absorbed iodide via the oxidation of ROS, which mostly reacted with tyrosine and diff used into the seawater as iodiferous tyrosines including monoiodotyrosine, diiodtyrosine and thyroxin. However, when tyrosine levels were reduced in polyp tissue, iodine could enter another route to react with SCS and give rise to the iodinated form of this substance (ISCS), whose production caused the onset of strobilation (Berking et al., 2005).Consistent with this proposal, it was found that tyrosine was signif icantly decreased in the strobila in the present study (Supplementary Table S3).Moreover, monoiodotyrosine and diiodtyrosine were also detected with slightly increased abundance in the strobila (Supplementary Table S1).
3.3 Pathway enrichment of signif icantly changed metabolite abundances
KEGG pathway analysis revealed that the 149 diff erential metabolites were involved in 46 metabolism pathways (Supplementary Table S4), and eight pathways were found to be signif icantly enriched(adj.P<0.05) (Fig.4). ABC transporters were the most signif icantly enriched, followed by processes such as aminoacyl-tRNA biosynthesis, biosynthesis of amino acids, histidine metabolism, neuroactive ligand-receptor interaction, and glycerophospholipid metabolism.
In the histidine metabolism pathway of the early strobilae, the L-histidinol signif icantly decreased while the L-histidine and histamine both signif icantly increased. It is clear that histamine plays a wider and critical role as pro-inf lammatory mediator in inf lammatory reactions (Bachert, 2002). This result is consistent with our hypothesis that oxidative stress and inf lammation play particular roles in triggering strobilation.
In the neuroactive ligand-receptor interaction pathway, four neurotransmitters including L-glutamic acid, histamine, taurine, and leukotriene D4 signif icantly increased. This indicates the presence of neuronal activity associated with strobilation. In previous studies, it was also shown that neurosecretory compounds might play a role in strobilation induction or progression (Crawford and Webb, 1972; Loeb and Hayes, 1981). Loeb and Hayes (1981) found that inC.chesapeakeipre-strobila polyps and early strobilae but not in young polyps, the calyx neurons secreted neurosecretory material, which was deposited into the surrounding tissues. Crawford and Webb (1972)observed that neurosecretory granules were most concentrated in the cell body, particularly in the axons of the segmenting strobilae, during the neck-formation stage. However, these were largely absent during the later strobilation stages.
4 CONCLUSION
Using an untargeted metabolomic analysis approach, we obtained comprehensive metabolome prof iles of the polyp and early strobila in the jellyf ishR.esculentum. We identif ied and provided a list of potential key metabolites and metabolic pathways for strobilation induction or progression. Of particular importance was the indole and indolizine-containing metabolites, phospholipids and eicosanoids, which provided new insights into the endogenous metabolism responses of strobilation. The functions of those key metabolites in strobilation still need further investigation. Our results provide valuable reference for further investigations of the molecular mechanisms ofR.esculentumstrobilation and other scyphozoan species.
5 DATA AVAILABILITY STATEMENT
The raw LC-MS data that support the f indings of this study are available from the corresponding author upon reasonable request.
6 ACKNOWLEDGMENT
We thank Shanghai Luming Biological Technology Co. Ltd. for the help with the LC-MS/MS analysis.
 Journal of Oceanology and Limnology2022年1期
Journal of Oceanology and Limnology2022年1期
- Journal of Oceanology and Limnology的其它文章
- The adjoint-based Two Oceans One Sea State Estimate(TOOSSE)*
- Structure and formation of the South Yellow Sea water mass in the spring of 2007*
- Lagrangian eddies in the Northwestern Pacif ic Ocean*
- Seasonal variability in dissolved oxygen in the Bohai Sea,China*
- In-situ experiments reveal mineralization details of porphyry copper deposits
- Chemical composition and Pb(II) binding of dissolved organic matter in a hypersaline lake in China*