
驱动人体下肢运动的脑肌电相干同步方法
2022-03-02 14:16:20孙沁漪张小栋李存昕李瀚哲
西安交通大学学报 2022年2期
目前,下肢助力外骨骼技术已广泛应用于工业物资搬运、军事作战装备等领域,成为研究热点。人-机智能融合方法可以充分发挥人和外骨骼机器人的优势
。其中,针对下肢外骨骼中人体运动意图的准确快速识别又是其核心研究内容,亟需高稳健性及精准度的识别方法。目前主流的外骨骼人体运动意图的识别方法主要采用生物电信号或力学信号。基于生物电信号的意图识别可以很好地解决基于力学信号识别带来的控制滞后的问题,更加真实反映人体运动意图
。利用脑电信号及肌电信号各自的优势进行脑肌电融合识别下肢运动意图,相比于单一生物电信号,可以极大地提高识别稳健性和准确性,实现对更多运动模式的精准预测。
根据驱动下肢运动的神经传导通路及运动控制理论,脑肌电传导路径的不同导致来自同一运动意图的脑肌电存在信息上的差别,将影响脑肌电融合识别下肢运动意图的性能,为下肢外骨骼操控者的安全性带来较大隐患。有效估计这一传导时间可以保证脑肌电信息的一致性,提升脑肌电融合准确度,同时对神经肌肉传导通路病理学研究、外设控制优化研究有着重要的意义。
目前广泛用于识别时延的方法是基于估计信号交叉谱密度相位的斜率
,此种方法只能在典型相干频域内进行可靠的时延估计,不能用来估计窄频带相干信号的时延,而窄带相干常出现在人体生理数据上。
另一种方法基于大脑与肌肉间关系变化来评估脑肌电信息时延,即皮质-肌肉相干性(cortico-muscular coherence,CMC)。皮质-肌肉相干性是大脑运动感觉皮层与对侧肌肉的同步振荡现象,可以有效量化大脑皮质与肌肉间的耦合程度,已有大量的研究证明在肌肉进行稳态力收缩或等距收缩中,存在较强的皮质-肌肉相干现象,可从对应大脑及肌肉采集获得的数据进行量化评估,如脑电图(EEG)、脑磁图(MEG)、肌电图(EMG)等。采用基于CMC的分析方法,可以有效确定驱动下肢运动中大脑及肌肉信息交流的典型相干中心频率,并利用其估算信号间的时延。文献[5-6]设计了等距肌肉收缩实验,发现采集得到的感觉运动皮层MEG信号与对侧肌肉EMG信号存在beta频带的显著相干性(13~35 Hz)。针对上肢肌肉运动,皮质-肌肉相干性存在显著对侧现象,如文献[7]中,利用多元自回归模型(multivariate autoregressive,mvAR)对原始数据进行拟合就可以观测到这一现象。文献[8]设计了下肢胫骨前肌的稳态收缩运动,当持续输出大于30%最大自主收缩力(maximum volumetric contraction,MVC)时,可以发现脑电信号与肌电信号在14~30 Hz时具有显著相干性。基于以上发现,文献[9]基于皮质-肌肉相干性分析成功计算了Rossler系统时延,并运用于脑电与肌电时延估算。文献[10]利用皮质-肌肉的时延反映皮质脊髓束的传导效率,并设计了上肢稳态等距收缩时延来估算脑电与肌电的时延。文献[11]采用短时傅里叶变换计算皮质-肌肉相干性,并根据最大化相干性法则(maximising coherence principle,MCP)进行时延估计。现有的时延估计方法大多考虑脑电与肌电单一通道的相关,未考虑通道间耦合
。
针对上述问题,本文采用mvAR-MCP进行脑肌电相干性分析以实现脑肌电信息同步化。基于mvAR模型拟合下肢运动过程中的多通道脑电及肌电信号,组合单一被试的所有实验及通道信号数据,将认知过程作为整体来考虑,囊括脑肌电信号存在的通道之间耦合的同时,可以在较短时间间隔内高精度估算CMC;基于CMC确立脑电及肌电典型相干中心频率,利用MCP量化驱动下肢运动中脑电及肌电的时延,最后通过实验对本方法的性能进行验证。
1 驱动人体下肢运动的脑肌电传导机制
人体下肢运动是通过对大脑的控制和骨骼肌的动作实现,大脑和骨骼肌的活动产生了脑电和肌电信号。脑电是从头皮或颅内记录到的局部神经元电活动及神经细胞群突触后电位的总和。大脑运动中枢首先产生下肢运动意图,这一神经冲动向上传递经颅骨、头皮检测得到脑电信号,其产生机理如图1所示。
1.1 带状整地是呈长条状翻垦造林地的土壤。在山地带状整地方法有∶水平带状、水平阶、水平沟、反坡梯田、撩壕等;平坦地的整地方法有∶犁沟、带状、高垄等。
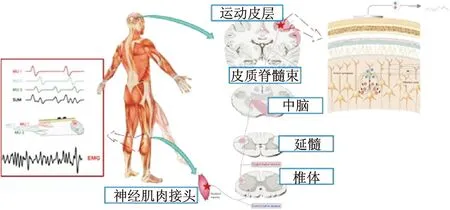
驱动下肢运动的意图主要产生于Brodmann大脑皮层分区的4区及6区。运动皮层存在有典型的对侧支配与倒立分布的特点,负责下肢运动的皮层又分布于运动皮质的顶部中心,对应前额叶顶部中心区域。在10-20国际脑电图标准中,与下肢运动相关的脑电通道为C3、C4、CP1、CP2和Cz通道。
电子商务的便捷、快速、实惠等等优势使得实体经济黯然失色,面对日新月异的网络销售,实体经济该如何发展才能走出困境呢?
运动中枢产生的神经冲动一方面向上传递至头皮产生脑电,一方面以电脉冲的形式沿皮质脊髓束向下传递产生肌电:沿中脑、延髓、椎体,最后经神经肌肉接头传递至下肢肌纤维处,如图1所示。肌肉纤维被不断激活,引起肌肉收缩、下肢动作。动作电位序列的相互叠加,经由皮肤表面的综合效应形成肌电信号。
式中:
为脑肌电
、
通道的相干性矩阵;
为利用模型系数估算得出的功率谱矩阵。
2 基于mvAR-MCP的脑肌电时延分析
2.1 mvAR模型拟合的脑肌电相干性分析
相干性分析广泛运用于研究两个同时测量信号之间的频域相关性,可用于检测两信号之间的时延并进行同步分析。传统的基于信号相干性的相位谱估计方法仅在不同频率上进行信号相干性分析,无法获得时间维上信号的相干性变化,因此本文基于时窗划分进行脑肌电时频相干性分析。
自回归(AR)模型广泛用于随机离散过程的描述
,但其只能用于一维时间序列的拟合。为了更加准确描述同步采集的多通道、多次实验的脑肌电信号,本文使用一种基于AR模型提升的多变量模型。考虑一个
维通道、
次实验的脑肌电数据同步采集过程,原始数据可以重构为集成多次实验的
×
维高维模型
,此高维数据可利用
阶mvAR模型进行拟合,如下式所示
El Nio事件:Nio3.4指数大于等于0.5 ℃持续5个月或以上记为一次El Nio事件。断点的处理:若指数大于等于0.5 ℃间断两个月或以上,则认为两次事件不连续;若间断一个月,并且该月的Nio3.4指数的3个月滑动平均值大于等于0.5 ℃则认为前后事件连续,否则不连续。El Nio爆发时间:Nio3.4指数第一次大于等于0.5 ℃的月份的前一个月。El Nio事件分类:SP型:爆发时间在2—5月;SU型:爆发时间在6—9月。CPC计算ERSST.v4资料得到的Nio3.4指数,如图1所示。

(1)
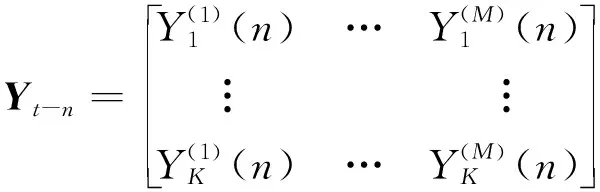
(2)
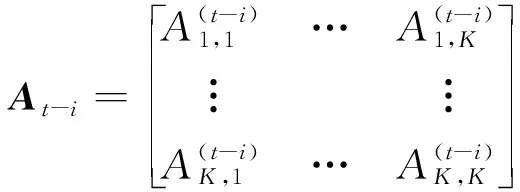
(3)
式中:
为重构后高维脑肌电数据矩阵;
为脑肌电数据采样点,
=1,2,…,
;
()
为第
次实验的脑肌电数据;
为脑肌电数据通道总数;
为维零均值白噪声矩阵,噪声方差矩阵为
;
(-)
为第
-
次实验的模型系数;
-1
,
-2
,…,
-
为
维mvAR模型系数矩阵,
为mvAR模型阶数。
2.2 AIC/BIC准则的mvAR模型阶数确定
(1)确定mvAR模型系数矩阵
如下所示
(
)=2
-
ln(det(
(
)))
(4)
式中:
为所用到构建模型的时间序列样本量;
为模型阶数;本文采集
次实验数据,每次实验数据样本量为
,
=
。
BIC与AIC两个模型均引入惩罚项控制模型参数,但BIC指标惩罚项较AIC指标更大,如下
(
)=
ln(
)+2ln(det(
(
)))
学生的提问涉及学生的理解能力,根据学生的提问进行分层解决,不同组的学生对问题的理解能力不一样,需要保证控制在学生的认知水平范围内进行分层提问,并针对学生的回答及时给予评价,从赏识教育的角度出发,在回答的问题上,多挖掘学生的闪光点,让其对积极思考,回答问题产生兴趣,久而久之,对学习就培养成一种积极探究的精神,深入思考的意识,逐渐能提高自己的学习能力,解决问题的能力,从而达到学习效果的提升,质量的提高。在分层异步教学中对整个班级涉及不一样难易程度的问题进行讲解,让学生自己选择,充分调动学生的积极性。
(5)
AIC和ABIC两个准则在选择模型时均兼顾模型的简洁性及精确性,在选择模型时取
及
达到最小时的模型阶数。
2.3 带遗忘因子的递推最小二乘算法的模型系数确定
(2)确定mvAR模型数据矩阵
如下式所示
在利用mvAR模型进行脑肌电数据拟合前,首先需要确定mvAR模型的阶数。可以根据赤池信息量准则(Akaike information criterion,AIC)及贝叶斯信息量准则(Bayesian information criterion,BIC)进行确定。AIC建立在熵的概念上,提供一种权衡估计模型的复杂度及拟合程度的标准。运用在本文mvAR模型选择上的AIC可以表示为模型拟合精度及参数个数的加权函数
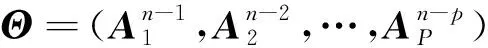
(6)
式中:
为mvAR模型阶数;
为当前采样点;
-
计算如式(3)所示。
确定模型阶数后,采用带遗忘因子的递推最小二乘算法估算拟合脑肌电数据的mvAR模型系数,过程如下。
=(
-1
,
-2
,…,
-
)
(7)
式中:
为最终量化的脑肌电时延;
为脑肌电典型相干的中心频率。
陈颐磊也知道,这种骂战对战局的形势发展毫无影响,顶多给自己打打气罢了。目前,全军弹药还能坚持几天,但八天来伤亡几千人已让他捉襟见肘,调兵到了山穷水尽的地步。假如鬼子再发动一次南门这样的攻势,城破或许只是一瞬间的事。
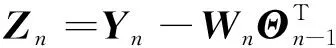
(8)
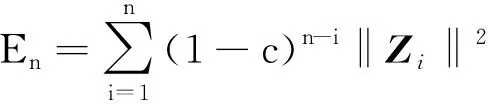
(9)
式中:
为瞬时误差估计矩阵,给出期望输出
与估计输出之间的误差;
为当前采样点;
为累积误差;
为遗忘因子,0<
<1确保遗忘距离当前时刻较远的脑肌电数据。
(3)用递归算法迭代计算
|min(
),确定最终mvAR模型系数矩阵
,根据mvAR模型瞬时误差估计矩阵可计算噪声协方差矩阵
,如下式所示
瞬变电磁波场变换与合成孔径成像技术是近年来发展相对较快的数据处理技术[13-14],该技术能够提取出瞬变电磁数据中所包含的电性界面的信息[14],降低体积效应的不利影响,为煤矿井下废弃巷道的探测提供了思路[15-18]。笔者针对工作面内部废弃巷道的探测问题,开展了基于波场变换的瞬变电磁合成孔径成像探测技术研究与工程应用。
其次,对于计算机数据的保护,我国刑法仅对获取这一行为进行处罚。而《公约》对于数据的保护规定了两个罪名:非法干扰计算机数据罪和非法拦截计算机数据罪。“获取”与“干扰”“拦截”相比,显然后者规定的范围广,因此更有利于对计算机数据的保护。

(10)
式中:
为当前采样点;
为实验次数。功率谱矩阵如下式所示
旁遮普天津技术大学机械工程专业应用型人才培养体系研究…………………… 徐超辉,赵 巍,RAMMIZ Ul-hassan,ASMA Bakht,MUHAMMAD Farhan-ejaz(71)
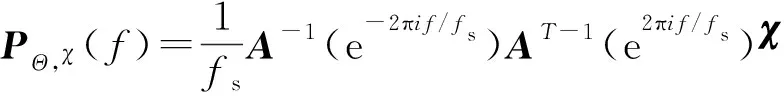
(11)
式中:
为当前脑肌电相干频率;
为脑肌电采样频率;
为mvAR模型系数矩阵;
为拟合噪声协方差矩阵,迭代计算方法如式(10)所示。
根据采集得到的脑电数据及肌电数据,选择4~16阶mvAR模型对重构脑肌电数据进行拟合,并根据模型参数个数及拟合精度计算
及
,可以获得4位受试者左右腿mvAR模型的
及
随模型阶数变化曲线,受试者S1的
、
随模型阶数变化曲线如图5所示。
右美托咪定联合舒芬太尼静脉自控镇痛用于食管癌患者术后镇痛的效果及对机体免疫功能的影响 …… 高彦东等(24):3427

(12)
由驱动下肢运动的脑肌电传导机制可知,经由头皮采集得到的脑电信号及下肢肌肉表面采集得到的肌电信号具有不同的传导机制,同时采集得到的肌电信号蕴含的下肢运动意图信息滞后于脑电信号,这一时延将减弱脑肌电相干性
,最终影响脑肌电融合的下肢运动意图识别准确率。同时驱动下肢运动中的脑肌电涉及大脑运动皮层区域的多通道脑电信号及下肢表面肌肉的多通道肌电信号,仅使用一对一通道相干性分析并不能完整反映人体神经传导及运动控制的机制,应采用高维模型进行脑肌电数据拟合,分析多通道间耦合更加符合脑肌电传导机制的生理基础,因此本文采用mvAR-MCP方法进行驱动下肢运动的脑肌电相干性分析及时延量化,进而谋求信息同步化方法,以完成驱动下肢运动的脑肌电信号同步化。
根据以上计算,可得到脑肌电显著相干出现的中心频率
及时刻
。
2.4 基于MCP的脑肌电时延估计
传统时延估计方法主要基于相位谱分析
,采用两时间序列互谱密度函数的相位谱斜率预估时间序列间的时延。但是,此种方法只能在信号的典型相关频带内进行可靠的时延估计,不能用来估计窄频带相干信号的时延。驱动下肢运动神经传导通路的不同导致采集脑电及肌电的不同步,根据mvAR模型获得的显著相干中心频率可以视作皮质与肌电信息交流最显著的频带,时延
的存在导致两相干序列的相干性下降,在显著相干时刻及频带处人为地将滞后信号随时间点向前移动,序列相干性将显著提升
,当
=
时,相干性达最大值,此方法即为MCP方法,通过人为移动脑肌电序列可量化脑肌电之间的时延,如式(13)所示
=max(
(
))
(13)
式中:
-
为重构的脑肌电数据,如式(2)所示;
为当前采样点。
2.为错题本标好页码,并在摘抄错题时按顺序为每道题标记题号,方便在做相关题分析、总结关联时,可以更快知道某道错题与错题本上相关的错题有哪些和在哪一页;
3 数据采集与处理
3.1 实验系统
为验证本文提出的脑肌电同步化方法有效性,本文设计人体下肢稳态力输出实验和抵消脑肌电时延后下肢运动意图识别实验并搭建相应平台,以人体下肢稳态力输出实验为例,进行实验并验证方法。实验硬件系统如图2所示,包括肌电采集电脑A、脑电采集电脑B、打标盒A、打标盒B、压力检测设备、WIFI、脑电帽及肌电仪。其中脑电帽为博睿康公司生产的32导脑电采集设备,用于采集脑电数据。肌电仪为博睿康公司生产的16导肌电采集设备,用于采集肌电数据。均自带24位AD转换器,共模抑制比为100 dB,采样频率为1 000 Hz。实验开始时,采集获得的肌电数据及脑电数据经WIFI模块分别无线传输至肌电采集电脑A及脑电采集电脑B,同时打标盒对脑电及肌电进行数据对齐,打标盒A及打标盒B分别同时对肌电采集电脑A及脑电采集电脑B进行打标,标记实验开始及结束时刻。与此同时压力检测设备采集受试者足底压力,传输至脑电采集电脑B并进行实时显示。
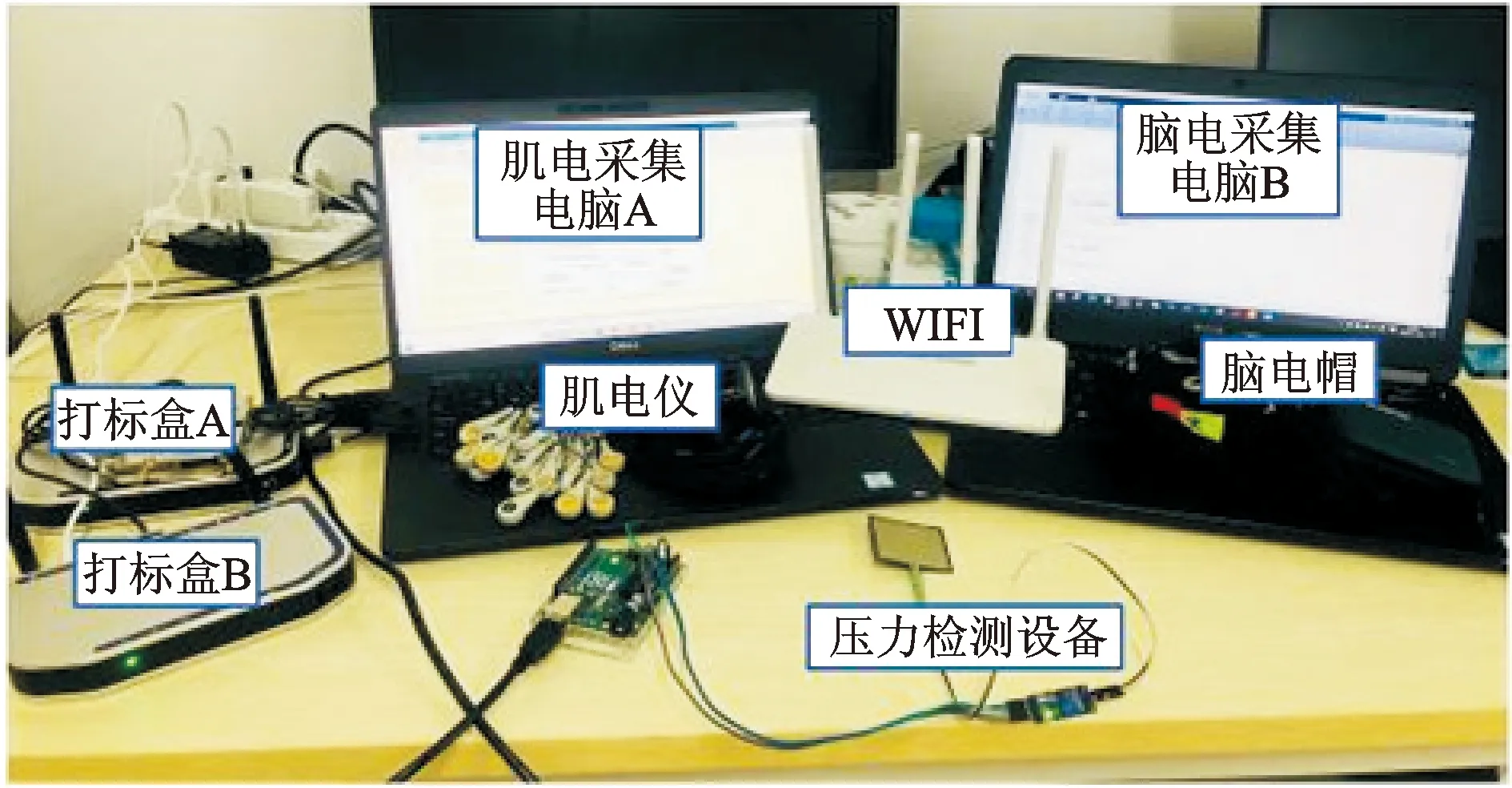
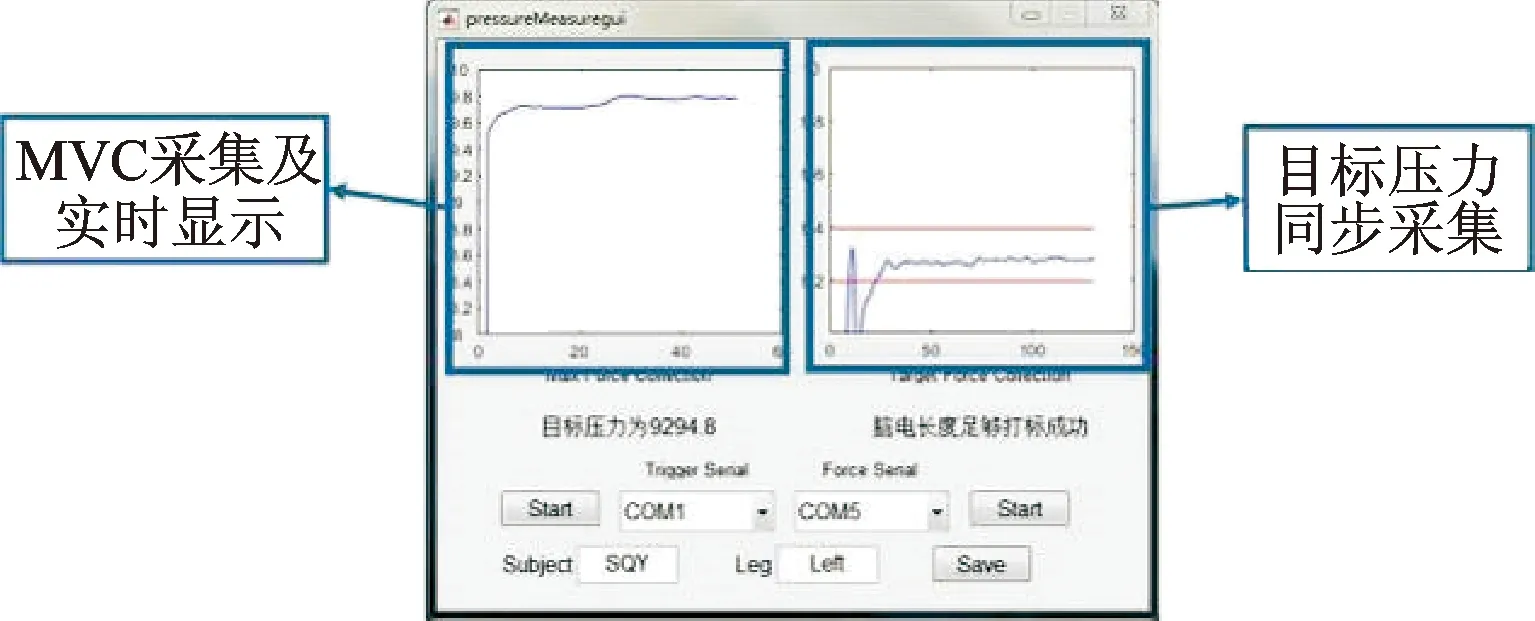
稳态力输出实验的实验软件系统如图3所示,左边界面显示最大自主收缩力
的采集流程及目标压力为0.8
,右方界面实时采集实验中受试者足底压力并进行显示,上下红色区域为根据最大自主收缩力确定的目标压力区间,采集结束后进行数据确认,显示“脑电长度足够且达标成功”意味着本次实验数据有效,可进行后续的数据保存等步骤。
初中生具有强烈的自我表现欲,为了充分发挥这种欲望,同时也为了突出英语教学的实践性和交际性,我们经常让学生排练课本剧,学生对于笔者这种教学方式也很喜欢。每逢我们排练节目时,他们都能积极完成教师所布置的各项任务。如在笔者的一次公开课上,学生自编自导自演的外星人驾驶飞碟来到地球这一剧情,扮演外星人的学生那夸张怪异的造型以及含混不清的外星语言,加上其他人物的巧妙配合,这一剧情被他们表演得十分滑稽有趣,获得了听课同行的一致好评。
3.2 受试者及实验范式
挑选S1~S4这4位受试者(1位女性、3位男性,平均年龄为(25±1.9)岁)进行下肢稳态力输出实验,其中3位有脑电及肌电相关实验经验;挑选S5~S12这8位受试者(2位女性、6位男性,平均年龄(26±1.5)岁)参加脑肌电融合识别实验。以上所有受试者均无任何肢体或神经系统疾病。实验中同步采集受试者的脑电及肌电数据,脑电电极按照国际标准10-20系统放置,采集受试者32导脑电通道信号。肌电采集共计16通道,左右腿各8个通道,所采集的肌肉分布在大腿及小腿上,包括股直肌、股外侧肌、股内侧肌、股二头肌、半腱肌/半膜肌、胫骨前肌、腓肠肌外侧、腓肠肌内侧在内的8块下肢表面肌肉。
下肢稳态力输出实验中,受试者双目直视屏幕,呈坐姿,臀部接触椅子且大腿悬空,大腿及小腿呈90度,双腿分开同肩宽,足底压力传感器放置于脚后跟底面,实验场景如图4a所示。正式实验开始前,采集每位受试者左右腿的最大自主收缩力,受试者保持踝部背伸,用脚后跟底面尽力下压足底压力传感器并保持至少10 s,获得左右腿的下肢足底的最大自主收缩力。根据左右腿的最大自主收缩力可以获得左右腿的目标足底力输出区间,由于足底压力传感器受到除了受试者施加的压力还包括下肢自身重力,因此目标区间设置较高,为0.8
±10 N。受试者需完成单腿各10次实验,每次实验时长总计20 s。实验中的脑肌电数据同步由打标完成,实验开始前倒计时3 s,开始时声音提示受试者进行10 s准备阶段,在准备阶段,受试者调整相应腿的力输出,使其位于目标区间范围内,10 s后进入正式脑电及肌电数据采集,受试者尽力保持10 s同一力输出至实验结束,5次重复实验后休息5 min。实验流程如图4b所示。

3.3 脑电及肌电数据预处理
脑电及肌电数据采集频率均为1 000 Hz,脑电数据经1~100 Hz滤波后,去均值,采用自适应滤波去除50 Hz工频干扰;肌电经1~200 Hz滤波后,去均值,采用自适应滤波去除50 Hz工频干扰。选择分布在额叶顶部的初级运动皮层处C3、C4、Cz、Cp1、Cp2脑电通道,以及下肢股直肌、股外侧肌、胫骨前肌、腓肠肌外侧、腓肠肌内侧等肌电通道的数据,单个受试者10次实验共11通道的数据重构成原始高维数据
。
4 结果与讨论
4.1 mvAR模型阶数确定及脑肌电时频相干性
(4)利用mvAR模型拟合的脑肌电数据
之间的相干性矩阵可用功率谱得出,如式(12)所示
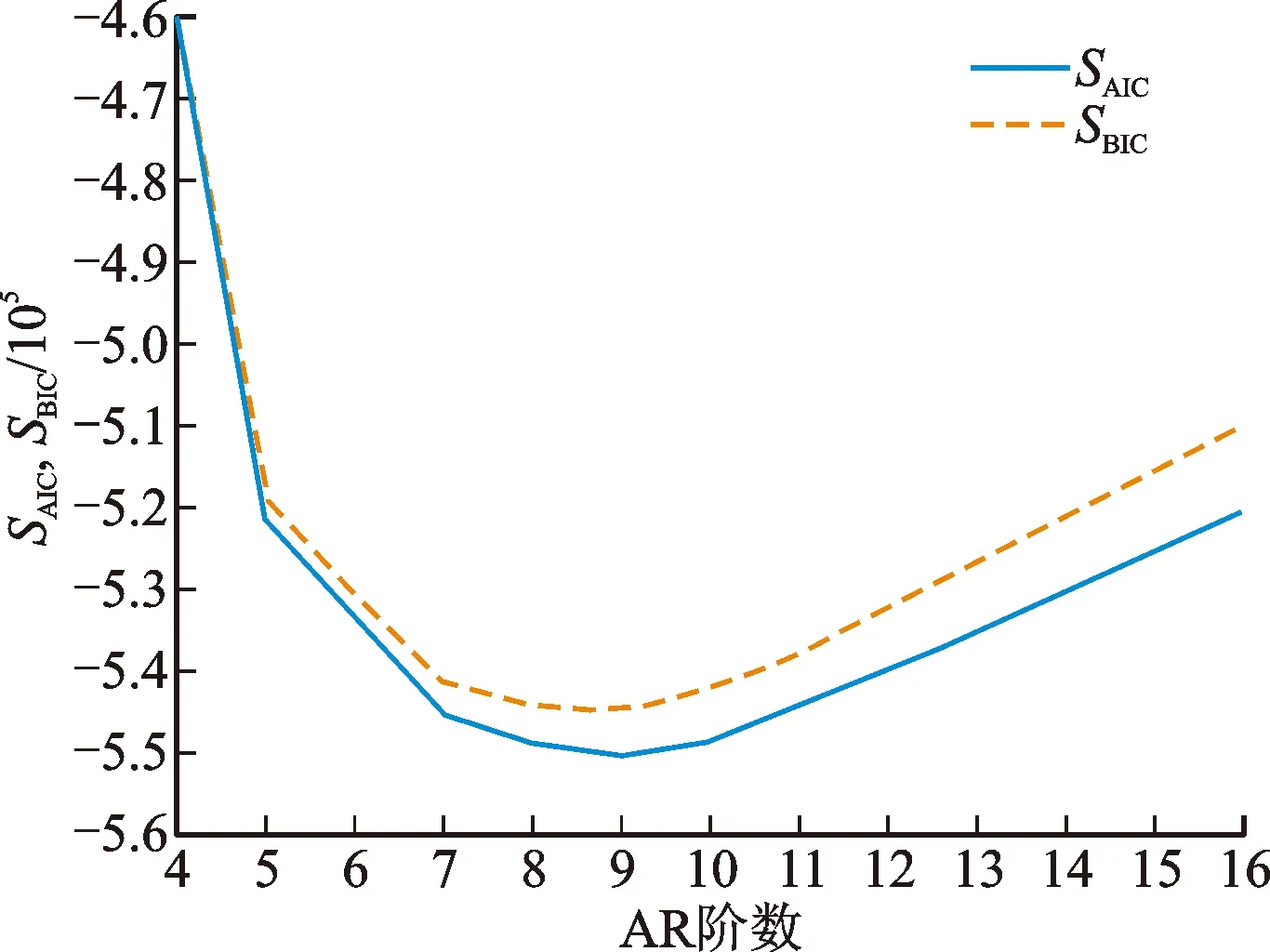
由图中结果可知,当模型阶数从4增大至16时,模型
及
具有类似的变化规律,且都会在4~16中间取得最低值,找到
及
最低时对应的模型阶数,即可获得每位受试者左、右腿的模型最终选择阶数,如表1所示。

根据以上最优模型阶数,可以对受试者左右腿分别建立mvAR模型进行拟合,并进行脑肌电相干性分析。取500 ms汉宁窗、400 ms重叠,频率分辨为0.5 Hz,选取时长为5 s的准备阶段及其后达到目标输出且保持下肢稳态力输出5 s的数据段进行相干性分析,可以获得每位受试者的脑肌电时频相干谱。分别对比各受试者脑肌电时频相干性,可以获得具有显著相干的脑肌电通道,如图6所示。受试者S1在左腿的下肢稳态力输出实验中,Cz脑电通道与大腿股直肌、股外侧肌、股内侧肌等肌电通道具有显著相干,同时随稳态力输出保持时间的增加,相干也逐步增强。显著相干频带分布于20~40 Hz之间,即对应脑电beta频段的主要分布区间,图中前5 s为准备阶段数据,从0时刻起,后5 s为输出力达到目标输出区间内的有效数据,S1在2.4 s左右时,Cz脑电通道与肌电通道间的相干性达到峰值。计算得到每位受试者所有脑电及肌电通道间的时频相干谱,并将相关性低于0.1部分置0,可以确定具有显著相干的脑肌电通道及对应的显著相干中心频率和时刻,如图6所示。

从受试者显著相干通道间时频相干谱中可以发现:各受试者在下肢稳态力输出实验中,呈显著性相干的脑电与肌电通道,脑电通道大部分为Cz通道,分布于前额叶顶部初级运动区,为控制下肢运动的运动感觉区域,与大脑皮层的运动控制理论一致;各受试者的显著相干频带,即皮质-肌肉耦合最强的频带,均分布在20~50 Hz范围内,出现时刻大多在0时刻以后,具体峰值相干出现的中心频率与时刻如表2所示。
充分发挥诉前程序作用。牢牢把握“诉讼不是目的、维护公益才是目的”的价值目标,将工作的重点放在诉前督促整改环节,督促行政机关履职。通过发出检察建议、约谈、走访、诉前圆桌会议、诉讼风险提示等方式,督促行政机关整改纠错,全面履行法定职责。三年来,全省检察机关共向行政机关发出督促履职检察建议6577件,各地政府职能部门积极整改、主动履职,诉前整改率从2016年的69%上升到目前的93.4%。
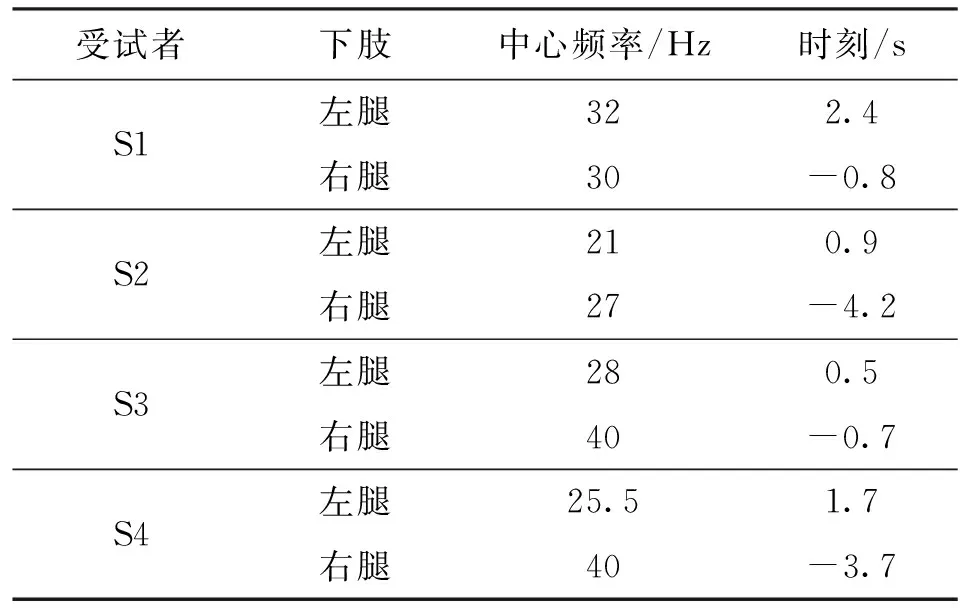
由表2可见,各受试者脑肌电显著相干中心频率大分布在20~30 Hz之间,S2受试者在-4.2 s时出现显著相干,推测为该被试在准备时间内即已达到目标区间力输出,因此早于其他受试者出现显著相干。所得到的的显著相干中心频率分布区间与现有研究一致
,例如早在2000年,Gross等就已发现人体执行等长收缩运动时,在20 Hz为中心的频带存在较强的皮质肌肉相干现象
。Möller等使用mvAR模型发现脑电中的相干性
。2012年,Muthuraman等在beta频带发现较强的脑肌电相干性
。基于表2各受试者通道的显著相干频率及时刻,可以使用最大化相干性法则进行脑肌电时延估算。
4.2 最大化相干性法则的时延估算
确认各受试者显著相干出现的中心频率及时刻后,使用最大化相干性法则对脑肌电间时延进行估算。根据人体神经传输及运动控制等先验知识,肌电的产生滞后于脑电,因此将每位受试者显著相干出现的中心频率及时刻处的脑电及肌电进行50 ms数据点人工平移,计算相干性并多次实验进行平均后,可得到相干性变化曲线。图7为S1的脑肌电相干性变化曲线。

图7中,横坐标0时刻点对应未进行平移时,受试者该次单腿稳态力输出实验中,脑肌电的峰值相干性,横坐标负向代表将滞后肌电数据向滞后方向平移,正向代表将滞后肌电数据向提前方向平移。根据图7所示的脑肌电相干性变化曲线,可以发现受试者的脑肌电相干性在正向提前方向平移后逐步增加到达峰值后下降;向滞后方向平移肌电数据时,脑肌电相干性将显著降低。这意味着当脑肌电间的滞后时延被抵消时,脑肌电间相干性将达到峰值,而当时延人为增大时,这一相干性又会降至谷值。进行受试者确认峰值相干性对应时刻即可获得每位受试者单腿的脑肌电时延估算结果,如表3所示。

计算得到受试者的脑肌电时延均分布在20 ms左右,左腿肌电整体平均滞后于脑电(23.3±11.4) ms(平均值±标准差),右腿平均滞后于脑电(19.8±4.8) ms。结果表明,各个受试者之间的脑肌电时延指标并无显著性差异,具有普遍存在性。
在获得每位受试者脑肌电时延后,为了验证时延估计的有效性,选择S5~S12共8位受试者进行了抵消时延后的脑肌电融合识别下肢运动意图实验。实验中的8位受试者采用脑肌电平均时延进行补偿,获得8位受试者的识别准确率如表4所示。最后,为验证本算法的稳健性,增加滑动时窗长度进行脑肌电融合识别,减少抵消时延部分与脑肌电数据的占比,可得到最终结果如表5所示。
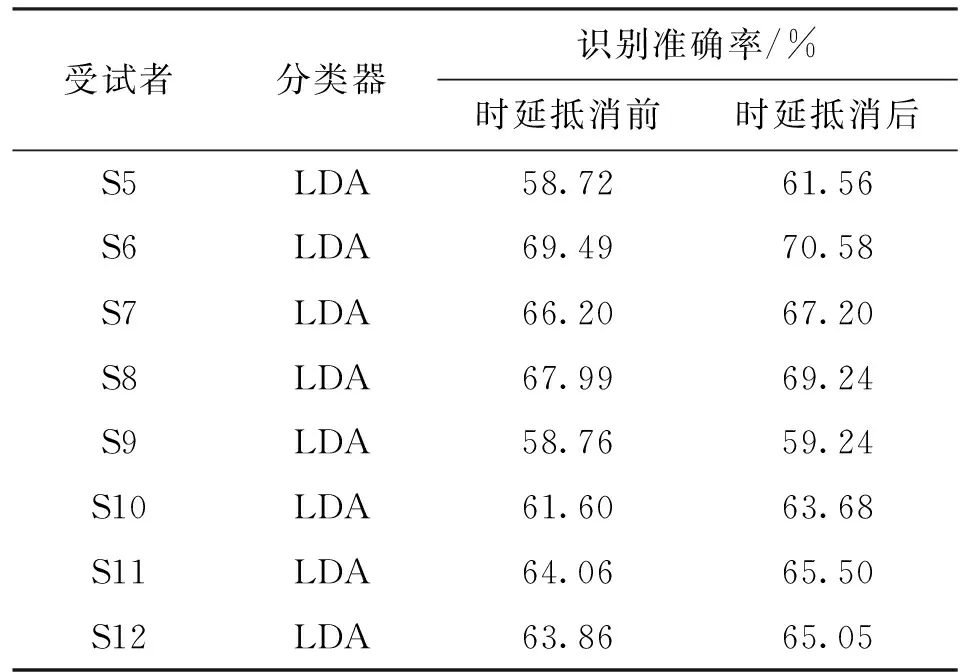
结合表4和表5可知,基于脑肌电特征融合下肢运动意图在抵消脑肌电时延后准确率有部分的提升,在50 ms时窗划分数据进行识别时,S5~S12这8位受试者的意图识别准确率平均提升了1.42%±0.68%(
=0.48),受试者脑肌电总体时延越大,时延补偿后提升效果越显著。当分类的分割时窗长度变长时,抵消时延部分占比减小,提升效果有所降低,但仍可提升原始脑肌电包含的驱动下肢运动信息,考虑到抵消时延数据量相对用于整体识别的脑肌电数据量较小,带来的部分准确率提升可视为有效。时窗长度变化后,本方法均可提升脑肌电融合识别下肢运动意图的准确率,抵消时延数据量相对用于整体识别的脑肌电数据量越大,提升效果越明显,因此本文所提出方法可有效同步脑肌电中的下肢运动意图信息,同时提升脑肌电融合识别稳定性。
教学内容是开展体育课堂教学的前提和基础,因此构建生活化的课堂教学,自然要引进生活化的教学内容。对于体育课堂教学,教师不能一成不变,要积极寻求新的亮点,寻找新颖和新奇,激发学生的学习兴趣,吸引学生的眼球,让学生积极踊跃参与体育运动。在小学的体育教学中,有很多基础知识对于小学阶段的学生来讲理解比较困难,因此教师就要引进生活化的教学内容,来进一步提高小学生的学习自信心。

对比现有的针对皮质肌肉相干性展开的研究,Muthuraman等基于离散傅里叶变换进行脑肌电相干性时频分析,确定了多通道脑肌电的典型相干频带
,但采用离散傅里叶变化分析精度有限。Anirban Chwodhury等采用手部运动中脑肌电相干性作为控制外设的信号源,采用信号互谱密度函数直接计算通道信号相干性
,此种方法可以快速进行两通道信号相干性的分析,但缺失了时间分辨率。Xu等采用短时傅里叶变换进行脑肌电相干性分析,并使用最大化相干性法则进行了脑肌电时延估算,得到脑肌电时延估算值约为25 ms的结果
,与本文估算结果类似,但短时傅里叶变换无法同时保证高精度的时间分辨率及频率分辨率。以上研究大多针对上肢所展开,且大部分只进行了脑肌电相干性的定性分析,并未进行脑肌电的时延量化。本文所提出的方法使用高维模型拟合同一被试所有实验及通道的脑肌电数据,可以在高精度的时间分辨率及频率分辨率的前提下进行脑肌电相干性分析,并在此基础上实现了脑肌电的时延量化。
目前,国内外采用脑肌电融合进行意图识别的研究仍处于信号组合的简单应用阶段,且缺乏理论研究。但已有研究证明了脑肌电中蕴含的运动意图信息不一致这一现象,如Liu等利用肌电进行下肢运动的标记,使用脑电检测运动意图,在运动真实发生的时刻以前即可采用脑电检测到运动意图信息
。同样在Eileen Lew、Mads Jochumsen等多个团队的研究中同样发现了脑肌电的这一特点
。由于同一运动意图诱发的脑肌电在时间上不一致,导致直接使用同步采集得到的脑肌电进行运动意图识别在理论基础上存在缺陷,而通过本文的研究证明了脑肌电的响应时间差,虽然这一时延并不大,但是在肌肉或精神疲劳状态下,这一时延带来的影响同样会增大,那么消除二者之间的时间差所带来的性能提升就会被放大。综上所述,本文所提出的方法实现了脑肌电运动意图信息的同步,最终使整体融合识别正确率有部分提升,在时窗变化时仍然保持了提升效果,不仅有限效增加了脑肌电融合识别的稳定性,也同时提升了脑肌电融合识别这一方法的逻辑性。
5 结 论
针对下肢运动意图驱动下肢运动中脑肌电信息不同步、现有的下肢运动特定脑肌电时延研究不充分的缺点,本文提出一种采用mvAR模型拟合驱动下肢运动多通道脑电及肌电信号,并基于计算获得的CMC采用MCP量化脑肌电时延的方法。通过脑肌电时延抵消实现了驱动下肢运动脑肌电信息同步化,并设计实验对方法的性能进行研究和验证,得出以下主要结论:①mvAR模型可以有效拟合多通道多次实验的脑肌电数据,进行高精度的时频相干性分析;②人体在进行稳态力输出时,脑肌电在beta频段具有显著相干现象。实验结果表明,S1~S4这4名受试者的左腿平均时延为(23.3±11.4) ms,右腿平均时延为(19.8±4.8) ms;③抵消脑肌电时延可以有效同步化脑肌电运动信息同时也增加了识别系统的稳定性。当采集时窗为50 ms时,S5~S12这8位受试者抵消时延后的脑肌电较未抵消时延的识别准确率提升了(1.55±0.86)%,且随时窗长度变大,仍可保持提升效果。本文方法及结论可提升脑肌电融合稳定性,同时为驱动下肢运动的神经肌肉传导通路研究、以脑肌电作为信息源的外设控制优化研究奠定基础。
由于伪迹信号以及信号的时变、微弱特性,脑肌电融合识别下肢运动意图的准确率依旧较低,还需要更加精准的滤波方法以及识别算法进行性能上的提升。同时,还需验证本文方法在不同类型受试者(如士兵、下肢功能障碍患者等)上的稳健性。以上研究内容将在研究的下一阶段中进行补充完善。
:
[1] 张腾, 张小栋, 张英杰, 等. 引入深度强化学习思想的脑-机协作精密操控方法 [J]. 西安交通大学学报, 2021, 55(2): 1-9.
ZHANG Teng, ZHANG Xiaodong, ZHANG Yingjie, et al. A precise control method for brain-computer cooperation with deep reinforcement learning [J]. Journal of Xi’an Jiaotong University, 2021, 55(2): 1-9.
[2] 李瀚哲, 张小栋, 李睿, 等. 利用运动准备电位的人体下肢自主运动意图预先感知方法 [J]. 西安交通大学学报, 2019, 53(10): 16-23.
LI Hanzhe, ZHANG Xiaodong, LI Rui, et al. A preperception method for voluntary movement intention of lower limb using readiness potential [J]. Journal of Xi’an Jiaotong University, 2019, 53(10): 16-23.
[3] BROWN P, FARMER S F, HALLIDAY D M, et al. Coherent cortical and muscle discharge in cortical myoclonus [J]. Brain, 1999, 122(3): 461-472.
[4] LINDEMANN M, RAETHJEN J, TIMMER J, et al. Delay estimation for cortico-peripheral relations [J]. Journal of Neuroscience Methods, 2001, 111(2): 127-139.
[5] CONWAY B A, HALLIDAY D M, FARMER S F, et al. Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man [J]. The Journal of Physiology, 1995, 489(3): 917-924.
[6] SALENIUS S, PORTIN K, KAJOLA M, et al. Cortical control of human motoneuron firing during isometric contraction [J]. Journal of Neurophysiology, 1997, 77(6): 3401-3405.
[7] XIFRA-PORXAS A, KOSTOGLOU K, LARIVIERE S, et al. Identification of time-varying cortico-cortical and cortico-muscular coherence during motor tasks with multivariate autoregressive models [C]∥2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. Piscataway, NJ, USA: IEEE, 2018: 1024-1027.
[8] HASHIMOTO Y, USHIBA J, KIMURA A, et al. Correlation between EEG-EMG coherence during isometric contraction and its imaginary execution [J]. Acta Neurobiologiae Experimentalis, 2010, 70(1): 76-85.
[9] GOVINDAN R B, RAETHJEN J, KOPPER F, et al. Estimation of time delay by coherence analysis [J]. Physica: A Statistical Mechanics and Its Applications, 2005, 350(2/3/4): 277-295.
[10]MENG Fei, TONG Kaiyu, CHAN S T, et al. Cerebral plasticity after subcortical stroke as revealed by cortico-muscular coherence [J]. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 2009, 17(3): 234-243.
[11]XU Yuhang, MCCLELLAND V M, CVETKOVIC Z, et al. Corticomuscular coherence with time lag with application to delay estimation [J]. IEEE Transactions on Biomedical Engineering, 2017, 64(3): 588-600.
[12]郑行. 上肢运动的皮层肌肉耦合及分析方法 [D]. 杭州: 杭州电子科技大学, 2019: 13-23.
[13]MIMA T, STEGER J, SCHULMAN A E, et al. Electroencephalographic measurement of motor cortex control of muscle activity in humans [J]. Clinical Neurophysiology, 2000, 111(2): 326-337.
[14]MIMA T, HALLETT M. Electroencephalographic analysis of cortico-muscular coherence: reference effect, volume conduction and generator mechanism [J]. Clinical Neurophysiology, 1999, 110(11): 1892-1899.
[15]崔冬. 多通道脑电信号建模及同步分析 [D]. 河北秦皇岛: 燕山大学, 2011.: 11-16
[16]杨芳梅. 基于神经肌肉功能耦合的脑肌电多尺度同步分析方法研究 [D]. 河北秦皇岛: 燕山大学, 2017: 13-24.
[17]MENG Fei, TONG Kaiyu, CHAN S T, et al. Study on connectivity between coherent central rhythm and electromyographic activities [J]. Journal of Neural Engineering, 2008, 5(3): 324-332.
[19]DING Mingzhou, BRESSLER S L, YANG Weiming, et al. Short-window spectral analysis of cortical event-related potentials by adaptive multivariate autoregres-sive modeling: data preprocessing, model validation, and variability assessment [J]. Biological Cybernetics, 2000, 83(1): 35-45.
[20]CHWODHURY A, RAZA H, DUTTA A, et al. A study on cortico-muscular coupling in finger motions for exoskeleton assisted neuro-rehabilitation [C]∥2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. Piscataway, NJ, USA: IEEE, 2015: 4610-4614.
[21]GROSS J, TASS P A, SALENIUS S, et al. Cortico-muscular synchronization during isometric muscle contraction in humans as revealed by magnetoencephalography [J]. The Journal of Physiology, 2000, 527(3): 623-631.
[22]MÖLLER E, SCHACK B, ARNOLD M, et al. Instantaneous multivariate EEG coherence analysis by means of adaptive high-dimensional autoregressive models [J]. Journal of Neuroscience Methods, 2001, 105(2): 143-158.
[23]LIU Dong, CHEN Weihai, LEE K, et al. EEG-based lower-limb movement onset decoding: continuous classification and asynchronous detection [J]. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 2018, 26(8): 1626-1635.
[24]LEW E, CHAVARRIAGA R, ZHANG Huaijian, et al. Self-paced movement intention detection from human brain signals: invasive and non-invasive EEG [C]∥2012 Annual International Conference of the IEEE Engineering in Medicine and Biology Society. Piscataway, NJ, USA: IEEE, 2012: 3280-3283.
[25]JOCHUMSEN M, ROVSING C, ROVSING H, et al. Classification of hand grasp kinetics and types using movement-related cortical potentials and EEG rhythms [J]. Computational Intelligence and Neuroscience, 2017, 2017: 7470864.
猜你喜欢
中国典型病例大全(2022年7期)2022-04-22 06:38:47
原子与分子物理学报(2021年2期)2021-03-29 07:31:38
测绘学报(2017年1期)2017-02-16 08:24:44
天津体育学院学报(2016年3期)2016-12-18 08:24:38
现代电生理学杂志(2016年3期)2016-07-10 12:10:32
现代电生理学杂志(2016年4期)2016-07-10 12:02:17
现代电生理学杂志(2016年4期)2016-07-10 12:02:17
现代电生理学杂志(2016年1期)2016-07-10 10:20:58
现代电生理学杂志(2015年1期)2015-07-18 11:02:16
体育科学研究(2015年4期)2015-02-28 17:06:29
