
基于GEO数据库对胆管癌潜在分子靶点的筛选及生物信息学分析
2022-02-24 06:05刘树业
世界华人消化杂志 2022年3期
于 珍, 吴 静, 张 磊, 刘树业
于珍, 吴静, 张磊, 刘树业, 天津市第三中心医院检验科, 天津市重症疾病体外生命支持重点实验室, 天津市人工细胞工程技术研究中心, 天津市肝胆疾病研究所 天津市 300170
于珍, 博士, 主要从事消化系统肿瘤的靶向治疗研究.
0 引言
胆管癌是一种异质程度极高的恶性肿瘤, 多原发于胆管上皮细胞. 其发病率逐年增加, 患者5年生存率较低, 小于10%. 胆管癌侵袭性强, 其发生可能与胆汁淤积、感染及炎症相关, 但具体的分子发病机制尚不完全清楚. 胆管癌的早期症状隐匿, 不易发现, 多数患者确诊时已是晚期. 对于晚期胆管癌患者而言, 治疗方法有限, 预后较差. 使用标准化疗方案(吉西他滨和顺铂)治疗的患者中位总生存期仍不到1年. 靶向治疗可有助于改善晚期肿瘤患者的治疗效果, 提高患者生活质量, 因而获得广泛关注. 而目前, 用于胆管癌的靶向分子疗法尚未见批准. 开发有前景的分子靶点, 对于提高胆管癌靶向治疗的应用前景至关重要.随着高通量测序技术的不断完善和发展, 利用生物信息学技术挖掘胆管癌基因组潜在分子靶点成为可能. 本研究中, 我们利用基因表达综合(gene expression omnibus, GEO)数据库中的2组胆管癌表达谱芯片数据进行一系列生物信息学分析, 获得胆管癌与正常组织的差异表达基因, 探讨枢纽基因在胆管癌发生发展中的作用, 为胆管癌的靶向治疗提供潜在分子靶点.
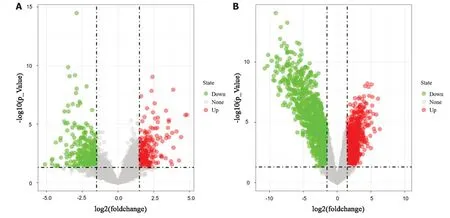
图 1 差异基因火山图. A: GSE26566差异基因火山图; B: GSE45001差异基因火山图.
1 材料和方法
1.1 材料 胆管癌数据集来源于GEO数据库, 编号分别为GSE26566和GSE45001. GSE26566包含104例胆管癌组织样本和6例正常胆管组织样本. GSE45001包括10个胆管癌组织样本和10例非肿瘤组织样本.
1.2 方法
1.2.1 差异表达基因筛选: 2组原始数据集采用GEO2R (http://www.ncbi.nlm.nih.gov/geo/geo2r/)在线分析工具进行数据处理, 处理后数据进行差异基因筛选. 调整后的P值(adj.P)<0.05和|logFC|>1.5作为纳入标准. 运用SangerBox在线软件作火山图. 用Venny 2.1.0对上述2个数据集的差异基因绘制Venn图取交集.
1.2.2 差异基因功能分析: 使用DAVID(http://david.ncifcrf.gov)在线分析对筛选出来的158个差异基因进行GO富集分析, 包括生物学过程(biological process, BP)、细胞组分和定位(cellular component, CC)以及分子功能(molecular function, MF), 同时进行KEGG信号通路分析.设置<0.05.
1.2.3 蛋白质互作网络与枢纽基因分析: 使用STRING 11.5数据库(https://sring-db.org/)预测胆管癌差异基因编码蛋白间的蛋白质互作网络(protein protein interaction network, PPI). 通过Cytoscape 3.7.0软件MCODE插件获得PPI中最显著的核心模块, 设置参数degree cutoff = 2、node score cutoff = 0.2、k-score = 2及max.depth = 100.利用CytoHubba插件Degree算法筛选出核心模块中前10位有较高连接度的枢纽基因.
1.2.4 枢纽基因表达验证: 使用GEPIA数据库(https://gepia.cancer-pku.cn/)对枢纽基因的mRNA在胆管癌组织及正常组织中实际表达水平进行验证.
采用Wilcoxon检验分析胆管癌组织和正常组表达数据.<0.05差异有统计学意义.
2 结果
2.1 差异表达基因筛选结果 经GEO2R分析, 分别从数据集GSE26566和GSE45001中提取到差异基因433个和1660个, 绘制火山图(图1), 其中绿色为下调基因、红色为上调基因. 取2个数据集差异表达基因的交集, 制作Venn图(图2), 得到相同差异表达基因158个, 其中上调基因53个, 下调基因105个.
2.2 差异基因GO分析和KEGG分析 GO富集分析结果显示这些差异基因主要参与细胞对锌离子反应、增长的负调控, 并且参与细胞粘附、代谢以及蛋白质聚合等生物过程; 主要细胞成分位于外泌体、胞外区、弹性纤维、细胞器膜基质及细胞外基质; 主要分子功能与结合肝素、半胱氨酸型内肽酶抑制剂活性、蛋白质同源二聚化、受体结合及磷酸吡哆醛结合等相关(表1). KEGG信号通路分析显示, 这些差异基因主要参与矿物质吸收、碳代谢、丙酸代谢、代谢途径、PPAR信号通路、脂肪酸降解、细胞色素P450对异生物质的代谢、化学致癌、ECM-受体相互作用和糖酵解/糖异生等通路(表2).

表 1 胆管癌差异基因的GO富集分析

条目 通路 基因数 P值hsa04978 矿物质吸收 7 2.01E-05 hsa01200 碳代谢 9 1.05E-04 hsa00640 丙酸代谢 5 4.41E-04 hsa01100 代谢途径 30 5.77E-04 hsa03320 PPAR信号通路 6 0.001749 hsa00071 脂肪酸降解 5 0.002098 hsa00980 细胞色素P450对异生素的代谢 6 0.002715 hsa05204 化学致癌 6 0.003812 hsa04512 ECM-受体相互作用 6 0.005455 hsa00010 糖酵解/糖异生 5 0.01122
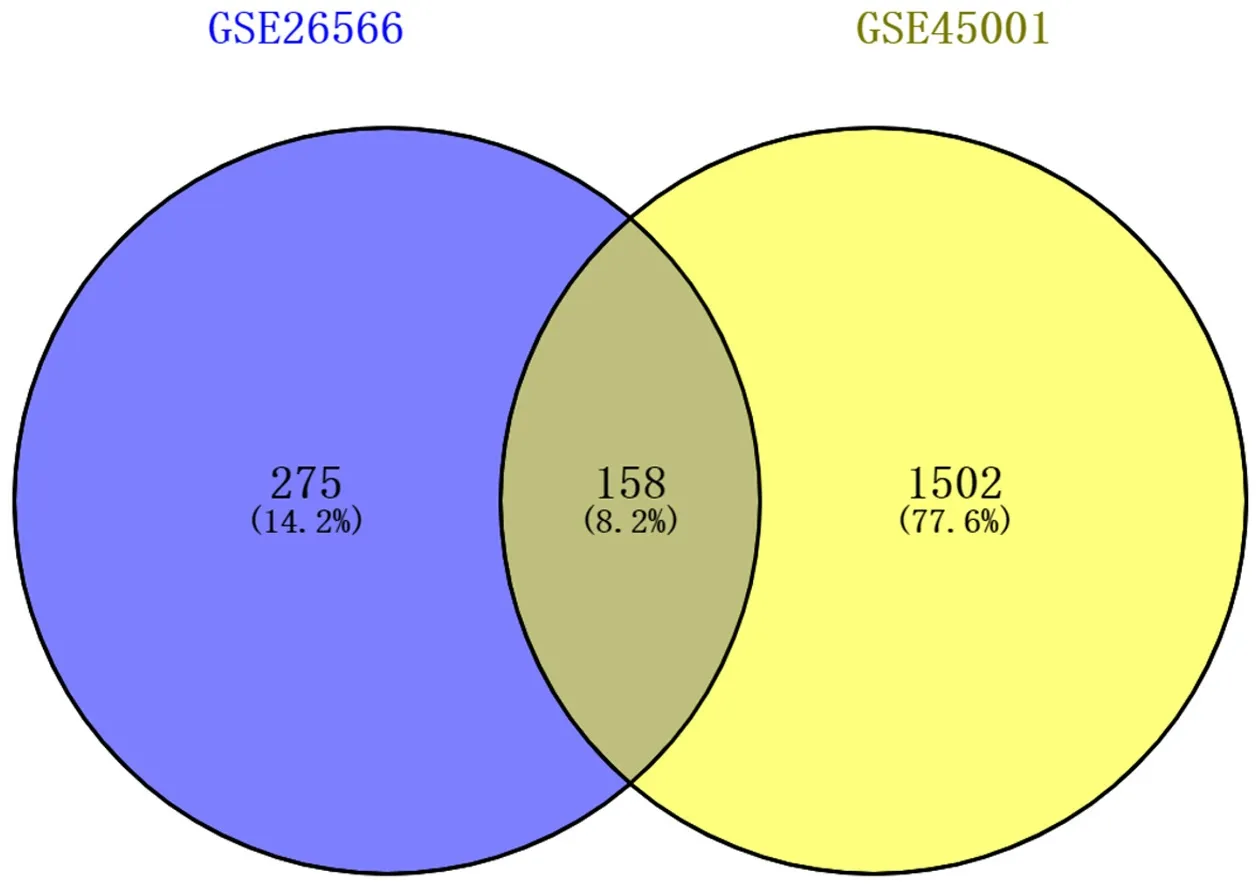
图 2 GSE26566和GSE45001差异基因Venn图.
2.3 蛋白互作网络与枢纽基因分析 除去游离的蛋白节点, 共有130个蛋白质节点连接形成382条复杂的PPI网络(图3).最显著模块由17个重要节点及127条边的网络构成(图4). 通过Cytohubba插件Degree算法确认枢纽基因, 结果如图5, 显示枢纽基因分别为NUSAP1, TOP2A, RAD51AP1, MCM4, KIAA0101, CDCA5, TYMS, ZWINT, 皆为上调基因.
2.4 枢纽基因转录表达水平验证 通过GEPIA数据库验证8个枢纽基因mRNA的表达水平. 如图6所示, 胆管癌组织中NUSAP1, TOP2A, RAD51AP1, MCM4, KIAA0101, CDCA5, TYMS和ZWINT的表达较正常组织均显著增加.
3 讨论
胆管癌是仅次于肝细胞癌的第二大常见肝脏原发性恶性肿瘤, 也是胆道最常见的恶性肿瘤, 根据解剖位置可分为肝内胆管癌、肝门周围胆管癌和远端胆管癌. 胆管癌发病早期症状隐匿, 晚期恶性程度高, 预后差, 所以寻找有助于治疗胆管癌的分子靶点至关重要.

图 3 共同差异基因的蛋白质互作网络图.
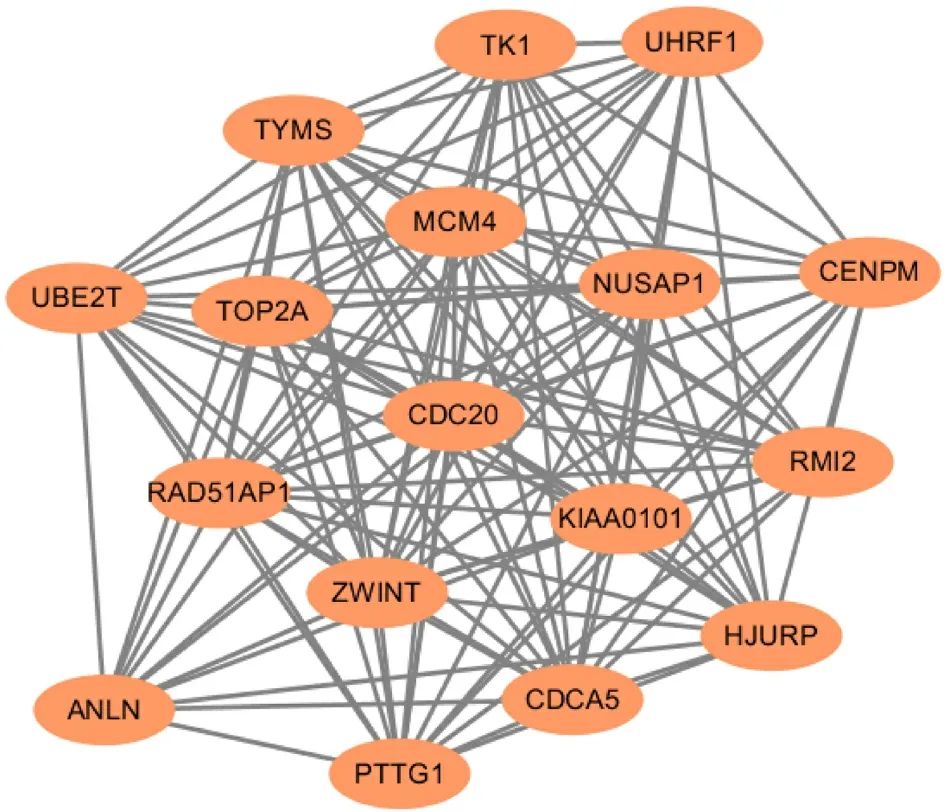
图 4 蛋白质互作网络图中的核心网络图.
本研究中, 我们使用GEO2R分析了GEO数据库GSE26566和GSE45001两组数据集.获得差异基因158个, 其中上调基因53个, 下调基因105个. 通过DAVID在线工具对共同差异基因做GO富集分析, 结果显示差异基因主要参与细胞对锌离子反应、细胞增殖的调控及细胞粘附、代谢等生物过程, 富集于外泌体、胞外区、弹性纤维等区域, 主要分子功能与结合肝素、半胱氨酸型内肽酶抑制剂活性、蛋白质同源二聚化、受体结合及磷酸吡哆醛结合等相关. KEGG信号通路分析表明, 差异基因主要参与矿物质吸收、代谢、PPAR信号通路及脂肪酸降解等过程.

图 5 枢纽基因模块.
经Cytoscape软件筛选, 本研究共得到8个枢纽基因, 分别是NUSAP1, TOP2A, RAD51AP1, MCM4, KIAA0101, CDCA5, TYMS, ZWINT, 皆为上调基因. 经验证, 这8个枢纽基因在胆管癌组织中的表达较正常组织高. NUSAP1编码核仁和纺锤体相关蛋白, 参与调节纺锤体的组装和维持正常的胞质分裂, 因此是细胞有丝分裂和增殖的重要调节因子. 研究发现, NUSAP1在多种肿瘤中存在过表达, 包括肝癌、肺腺癌和肾细胞癌. 另外, NUSAP1的上调与黑色素瘤和口腔鳞状细胞癌的恶性进展有关. 因此, NUSAP1可能是一个潜在的治疗靶点.DNA拓扑异构酶Ⅱ-α(TOP2A)基因参与调节核酸空间结构的动态变化, 其表达与肿瘤侵袭、复发以及不同癌症的预后相关, 包括乳腺癌、睾丸畸胎瘤、膀胱移行细胞癌、脑膜瘤、神经胶质瘤、肝癌和子宫内膜癌. RAD51AP1(RAD51-associated protein 1)基因编码一种DNA结合蛋白, 在RAD51介导的同源重组中至关重要. Obama等研究表明, RAD51AP1参与肝内胆管癌的细胞增殖和DNA修复, 其沉默有助于抑制胆管癌细胞的生长. MCM4是微染色体维持蛋白家族的成员之一, 具有复制解旋酶活性, 参与DNA复制和DNA解旋. 有研究显示, MCM4的高表达会增加多种肿瘤发病风险, 如肝癌、食管癌、乳腺癌等. KIAA0101编码一种保守的增殖细胞核抗原结合蛋白, 与DNA聚合酶竞争结合以调节DNA复制和损伤修复. 据报道, KIAA0101基因在乳腺癌和非小细胞肺癌组织中存在表达上调, 敲低KIAA0101基因可抑制乳腺癌细胞的生长, 另外KIAA0101高表达也与非小细胞肺癌患者术后复发相关. 细胞分裂周期相关蛋白5(CDCA5), 也称为Sororin, 在确保姐妹染色单体的准确分离方面发挥着关键作用. 最近的研究表明, CDCA5的高表达与多种癌症的肿瘤发生和组织侵袭相关, 包括口腔鳞状细胞癌、非小细胞肺癌、尿路上皮细胞癌和胃癌. TYMS编码胸苷酸合酶, 通过催化dUMP的甲基化产生dTMP, 在DNA复制中发挥重要作用. TYMS的高表达与许多实体瘤的发生存在关联, 包括肺癌、乳腺癌、胃癌、结肠直肠癌和肾细胞癌等. ZWINT编码一种调控着丝点分裂的动粒蛋白, 在维持有丝分裂周期过程中起着至关重要的作用. 越来越多的证据表明, ZWINT 在许多人类癌症中通常高度表达, 并且与预后不良和早期复发有关.
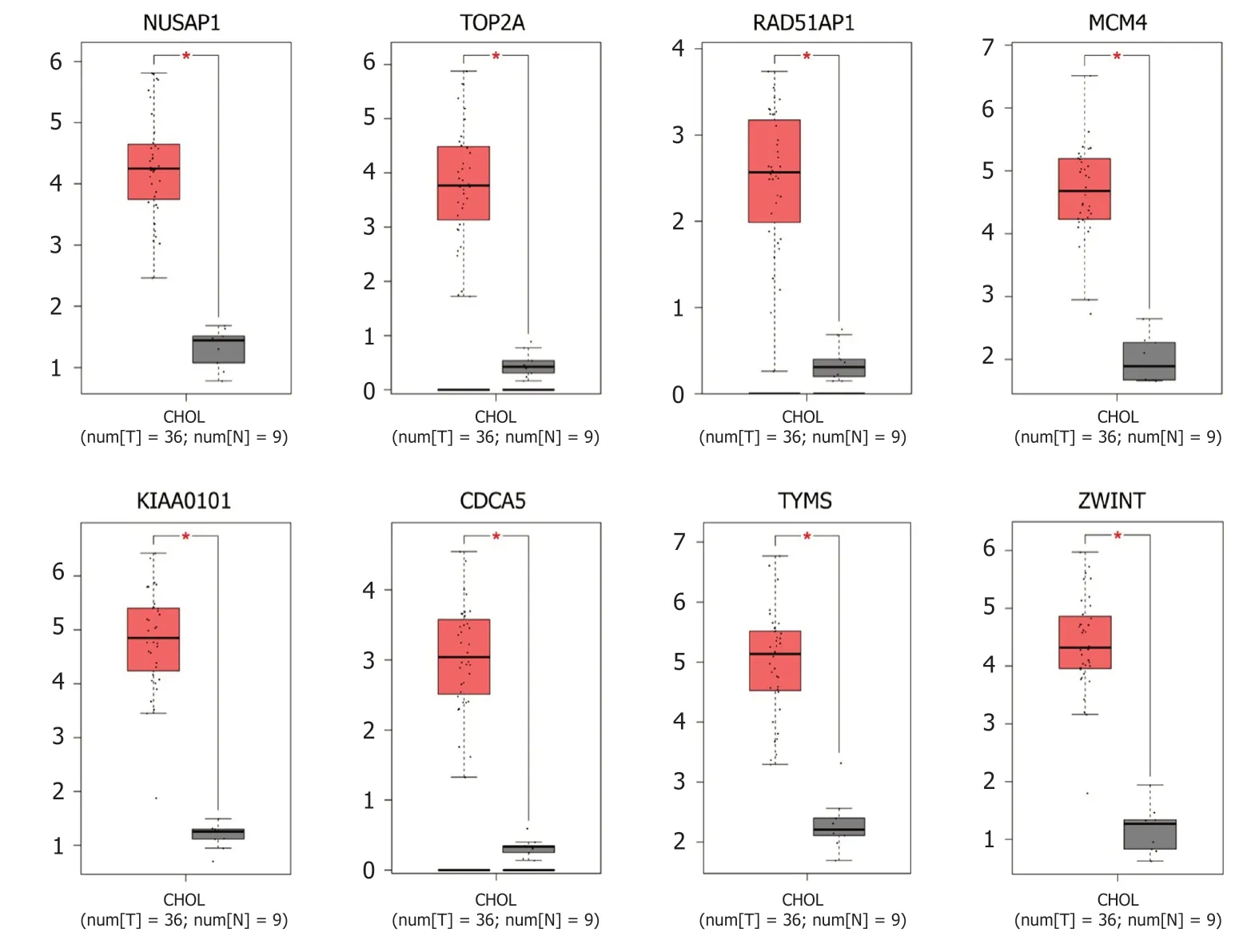
图 6 枢纽基因在肿瘤组织和正常组织中的表达验证.
4 结论
本研究通过生物信息学方法从胆管癌组织中筛选出8个枢纽基因, 与正常组织相比, NUSAP1, TOP2A, RAD51AP1, MCM4, KIAA0101, CDCA5, TYMS, ZWINT在胆管癌中的表达均上调. 结果提示, 这8个枢纽基因可能在胆管癌的发生发展中起到关键作用, 为胆管癌的靶向治疗提供了理论基础, 但这些基因在胆管癌中的具体机制和作用仍需要进一步实验验证.
文章亮点
胆管癌是胆道常见的恶性肿瘤, 预后不佳. 靶向治疗是改善胆管癌治疗效果的重要手段, 探索新的分子靶点对于胆管癌靶向治疗的研究十分必要.
寻找影响胆管癌发生和进展的枢纽基因对于开发胆管癌靶向治疗的靶点具有重要意义.
通过生物信息学方法探索胆管癌靶向治疗的潜在分子靶点.
利用GEO数据库分析2组胆管癌数据集, 得到共同差异表达基因. 通过GO富集分析与KEGG通路分析共同差异基因主要参与的生物学过程与信号通路. 基于String数据库构建蛋白质互作网络图, 经过Cytoscape软件筛选枢纽基因, 通过GEPIA数据库验证枢纽基因在胆管癌与正常组织中的表达量.
2组胆管癌数据集共包含158个共同差异表达基因. GO富集分析及KEGG通路分析结果显示, 差异基因主要参与细胞对锌离子反应, 细胞增殖与粘附等生物学过程, 涉及矿物质吸收、代谢、PPAR信号通路及脂肪酸降解等信号通路. 最终筛选得到了8个上调的枢纽基因, 在胆管癌组织中表达量显著高于正常组织.
本研究通过生物信息学方法获得了与胆管癌发生与进展息息相关的8个枢纽基因, 分别为NUSAP1, TOP2A, RAD51AP1, MCM4, KIAA0101, CDCA5, TYMS, ZWINT, 可能为胆管癌靶向治疗的潜在分子靶点.
这8个枢纽基因对胆管癌的影响和机制需进一步通过实验验证, 有望为胆管癌靶向治疗分子靶点的开发提供新思路.
猜你喜欢
传染病信息(2022年2期)2022-07-15
肝博士(2022年3期)2022-06-30
体育科技文献通报(2022年3期)2022-05-23
安徽医科大学学报(2022年4期)2022-05-12
幸福家庭(2021年3期)2021-03-30
世界中医药(2021年22期)2021-01-03
中国中药杂志(2017年15期)2017-08-30
中国中药杂志(2017年15期)2017-08-30
江苏农业科学(2017年5期)2017-04-15
湖北农业科学(2014年3期)2014-07-21
