
肾形肾形虫干预下甜菜幼苗的光合响应
2022-02-21 11:04崔晶晶窦骞瑶韩卓君潘恒艳宋柏权周建朝王秋红
中国农学通报 2022年36期
崔晶晶,王 丽,窦骞瑶,韩卓君,潘恒艳,宋柏权,周建朝,王秋红
(1黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2哈尔滨师范大学生命科学与技术学院,哈尔滨 150025)
0 引言
肾形虫属于单细胞原生动物,拥有着不少的生物量,仅次于细菌和真菌[1-2]。其广泛地分布在根系周围,其身体构造简单、且易于采集,是土壤生物量、活性和多样性的重要部分,因此在土壤生态功能中占据重要作用。土壤原生动物种类繁多,且它们的活动对根际或根围中土壤的理化性状影响的同时也直接影响了植物的生长和发育[3-6]。一些前者的研究[7-11]也表明原生动物不仅可以使土壤中养分物质进行矿化以及分解碳氮磷元素,也能够促进植株的生长以及生物量的积累。JENTSCHKE等[12]和BONKOWSKI等[13]在研究中发现原生动物会显著增加植株的生物量。BONKOWSKI&BRANDT[14]证明了原生动物对豆瓣菜幼苗有强烈的生长刺激作用,增加植株的生物量。此外,光合作用的提高可以促进植株生物量的积累,影响植物生长发育的过程[15-18]。ANDRALOJC等[19]研究也发现柳树的光合能力和其生物量是呈正相关,并且影响其生长。MU等[20]和URIBELARREA等[21]等发现光合作用与叶片的氮含量和叶片内氮在不同氮组分中的分配密切相关。刘晓明等[22]和李鸿妹[23]等发现适当的光照强度可以改变植物氮代谢相关酶酶活性,从而影响植株生长以及生物量的积累。刘德明等[24]也发现植株的氮效率会受到光合作用和干物质积累的影响。
甜菜(Beta vulgaris L.)是藜科,甜菜属二年生草本植物,主要分布在新疆、黑龙江、内蒙古等地[25]。甜菜是中国主要的糖料作物,也是主要的经济作物。目前甜菜的价值已经向多元化发展,不仅可作糖料,还可作生产燃料和饲料等用途[26]。随着研究人员对土壤生物学不断深入研究,发现甜菜的优质高产依赖于完善的种植技术及适宜的田间管理,但同时也会受到自然生态环境的影响。在甜菜根系与根际土壤的互作及调控下,挖掘和利用甜菜本身的遗传潜力,有目的地定向调控甜菜生长的环境条件,已成为研究的关注点。
有研究表明土壤动物提高植物的生物量及植株生长[27-28],但原生动物肾形虫对甜菜生长的相关指标评价分析方面研究较少。通过对比接种肾形肾形虫与未接种肾形肾形虫幼苗形态、生物量积累、光合特性、叶绿素和酶活性等相关性指标差异,分析肾形肾形虫对甜菜幼苗生长发育的影响,从而对甜菜根系与根际土壤、根际生物间的对话,以及根际生态微环境的深入研究提供参考,促进甜菜优质、高糖、高产的产业发展。
1 材料与方法
1.1 试验材料
甜菜土壤原生动物:肾形肾形虫(Colpoda reniformis)是肾形虫属(Colpoda)原生动物。
实验所用的原生动物活体肾形肾形虫是从甜菜根际土壤中筛选获得,并由哈尔滨师范大学生命科学与技术学院原生动物学研究室所研究分析出的肾形虫属中新物种。
甜菜:701(中国农业科学院甜菜研究所育种品系)。
1.2 试验设计
1.2.1 培养土壤原生动物 活体肾形肾形虫,浓度10000只/50m L。(哈尔滨师范大学生命科学与技术学院原生动物学研究室分析研究所得出最适浓度)
根据王香[11]方法对土壤浸出液中出现的活性原生动物进行观察鉴定6天之后,选择提取活体肾形肾形虫置于一塑料六孔板中,以米粒浸出液喂食,在20~25℃的恒温恒湿培养箱中进行大量培养。每24 h清理一次代谢废物。
1.2.2 甜菜种子预处理 采用王秋红等[29]方法。
1.2.3 营养液 稀释10倍的改良后的霍格兰(Hoagland)营养液[16]。
由于对比稀释5倍、10倍、20倍、30倍的改良后的霍格兰(Hoagland)营养液。在25℃恒温下培养,发现肾形肾形虫在稀释10倍的Hoagland营养液中生长情况及存活量最好。故选择稀释10倍的改良后的霍格兰(Hoagland)营养液。
1.2.4 培养方法 将处理过的甜菜种子均匀播种在装有蛭石的育苗盘中,每天定量浇蒸馏水,待种子出苗后,将幼苗移至50m L离心管中培养。在移苗之前用移液器吸取六孔板中培养的活体肾形肾形虫,分别加入每个离心管中10000只以及稀释10倍的改良后的霍格兰(Hoagland)营养液50m L,用封口膜在植株地上部和地下部交界处将离心管封口。3月31号开始处理,光照14 h,夜间黑暗10h。由于管中营养液的损耗及为了保持管中植物根系生长的液体环境的基本一致性,故每隔3~4天得更换营养液及肾形肾形虫。培养40天后分别取样。用不加肾形肾形虫的作对照,每种处理40个重复。
1.3 测定项目及方法
1.3.1 形态和生物量指标的测定 用直尺准确测量各植株的叶长、叶宽、地上部分高、根长等基础表型指标,然后用根系分析系统WinRHIZO(Seiko Epson Corp)扫描植株幼苗的总根长、根表面积、根尖数、分枝数等指标。将根、叶60~85℃烘干至恒重,用电子天平(0.01 g)称干重。计算叶长叶宽比、叶长叶柄比、根冠比。
1.3.2 叶绿素色度比值 用叶绿素仪(型号SPAD-502)测定单株功能叶SPAD值,各测10个重复,每片功能叶重复测定3次。
1.3.3 叶片光和作用测定 用美国LI-COR公司Li-6400便携式光合系统分析仪测定甜菜功能叶片的净光合速率(Pn)、胞间 CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr),每一叶片测定3次,每个处理测定10株。
1.3.4 叶绿素的测定 采用王学奎、黄见良[30]的方法。称取叶片0.2 g提取叶绿素,每个叶片3次重复,于分光光度计663、645 nm处测得吸光度,并计算叶绿素a、叶绿素b和总叶绿素的含量。
1.3.5 相关酶活性的测定 采用王学奎、黄见良[30]的方法,测定硝酸还原酶(NR)、谷氨酰胺合成酶(GS)。
1.4 氮效率相关指标计算

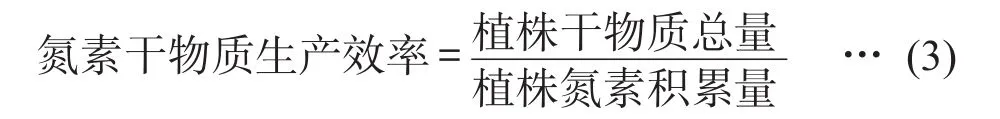
1.5 数据处理
用SPSS Statistics 25进行数据分析,Origin 2021作图,进行显著相关性分析。
2 结果与分析
2.1 接种肾形肾形虫对甜菜幼苗基本表型的影响分析
2.1.1 接种肾形肾形虫对甜菜幼苗形态特征及生物量的影响 表1表明,与未接种肾形肾形虫相比,接种肾形肾形虫对甜菜幼苗的总植株高、叶面积、根长都有显著增加,特别是叶片面积,达到了极显著水平(P<0.001),数据值为22.67 cm²,增加了15.49%。总植株高、根长也有显著影响(P<0.01),数据值分别达到为39.15 cm、26.84 cm,相对于对照,分别增加了14.14%、20.09%,但是叶片数、地上部高无明显变化。

表1 接种Colpoda reniformis对甜菜幼苗地上部形态性状的影响
图1可知,接种肾形肾形虫增加了甜菜幼苗生物量的积累。与对照组相比,地上部分干质量、地下部分干质量、和总干质量分别增加了10%、33.3%、14%,地下鲜质量增加了24.32%,但是接种肾形肾形虫对幼苗地上部鲜质量生物量的积累无显著影响。从图中看出,与对照相比,根冠比(鲜)增加了18.18%,根冠比(干)增加了20%。
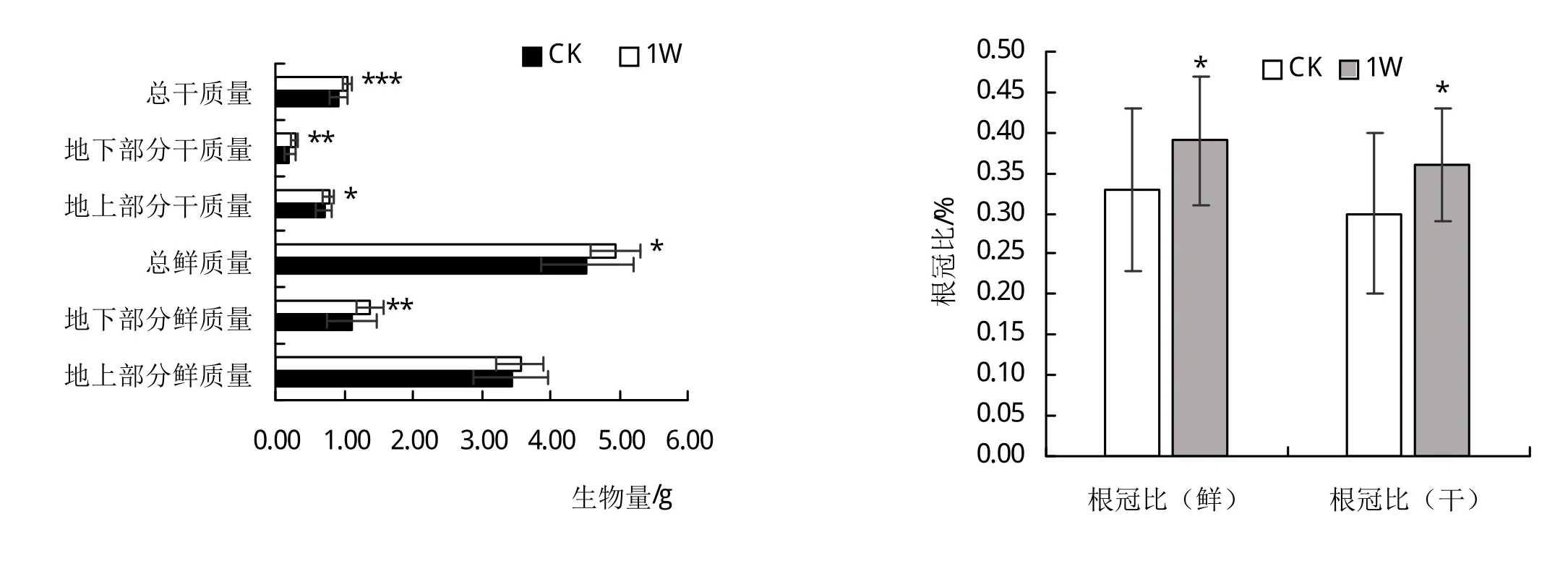
图1 接种Colpoda reniformis对甜菜幼苗生物量积累和根冠比的影响
2.1.2 接种肾形肾形虫对甜菜幼苗光合特征相关生理指标的影响 SPAD值可以反应植株叶片叶绿素的浓度。图2-a可知,接种肾形肾形虫的SPAD显著高于对照组,增加了33.2%。图2-b可知,接种肾形肾形虫后甜菜叶绿素的含量明显提高。与未接种肾形肾形虫处理相比,叶绿素a、叶绿素b和总叶绿素分别增加了31.58%、31.82%、31.72%。
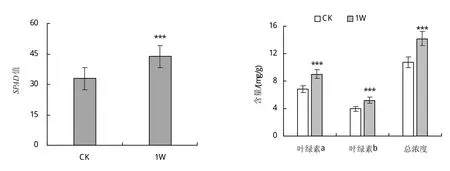
图2 接种Colpoda reniformis对甜菜幼苗SPAD值和叶绿素含量的影响
由表2可知,接种肾形肾形虫显著促进了甜菜幼苗的光合作用。与对照组相比,Pn、Gs分别增加了43.91%、40.56%,数据值为18.78、59.44;但Tr、Ci增加不显著。
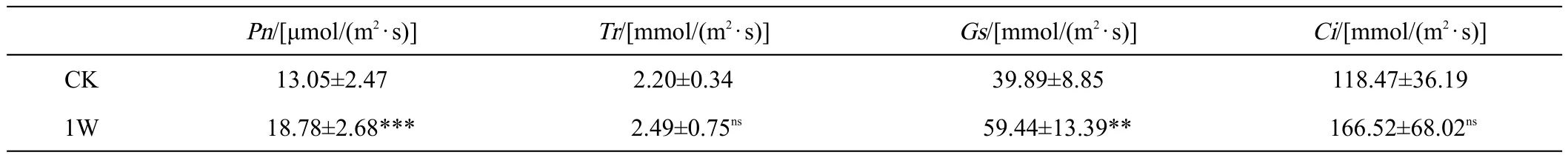
表2 接种Colpoda reniformis对甜菜幼苗光合特性的影响
2.1.3 接种肾形肾形虫对甜菜NR和GS相关酶活性的影响 由图3可知,相比对照,接种肾形肾形虫显著促进了甜菜幼苗的GS活性,增加了23.52%,但是NR活性差异不显著。
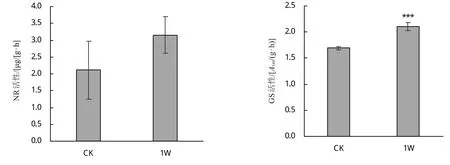
图3 接种Colpoda reniformis对甜菜幼苗NR和GS相关酶活性的影响
2.1.4 接种肾形肾形虫对甜菜幼苗氮素干物质生产效率和氮累积量的影响 从图中可以看出,相比对照,接种肾形肾形虫显著提高了甜菜幼苗的氮素干物质生产效率和氮素累积量,地上和地下分别增加6.3%、8.8%和5.3%、19.7%,说明接种肾形肾形虫的处理能够显著提高甜菜干物质累积量。
2.2 甜菜幼苗各种基础表型特征间相关性分析
通过对接种肾形肾形虫的甜菜与对照组的相关指标的比较分析,发现肾形肾形虫的干预对甜菜的生长发育是有影响的。但是甜菜各指标之间的联系与关系难以直观呈现,因此还需要对甜菜的综合指标进行相关性分析、聚类分析、主成分分析。
2.2.1 综合指标相关性分析 对甜菜幼苗的指标进行Pearson综合相关性分析(表3),结果如下:叶绿素a、叶绿素b以及总浓度是呈显著正相关(P<0.01),表明当叶绿素a呈值且越大,叶绿素b和总浓度就越大。地上部高、叶片面积、叶长、总生物量与Pn呈正相关,表明Pn越大越强,则有利于甜菜的生长以及植株生物量的积累。Pn与Gs呈正显著相关(P<0.01),由表可知,Gs越大则光合强度越强。氮效率与SPAD、Gs、Ci呈正相关,说明光合强度越大,氮效率越高。SPAD与NR呈正相关,但与GS呈负相关。NR与GS是显著负相关的。
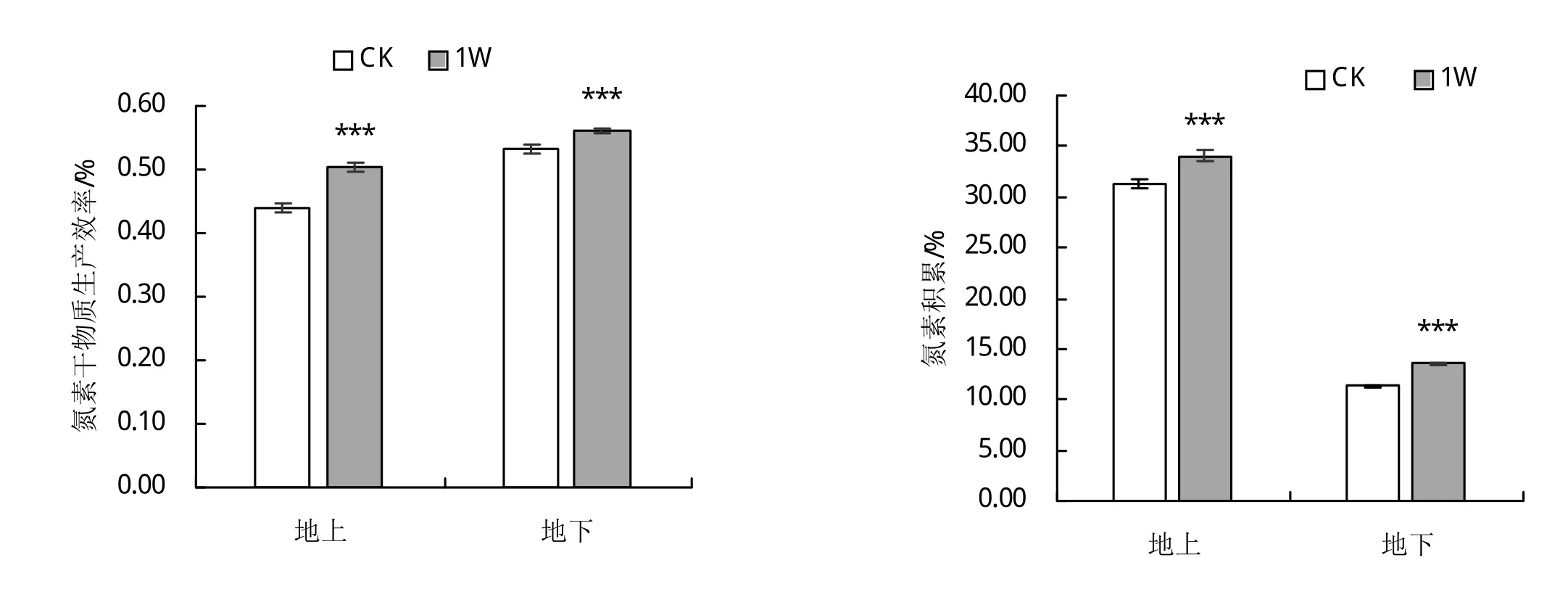
图4 接种Colpoda reniformis对甜菜幼苗氮素干物质生产效率和氮累积量的影响
2.2.2 聚类分析 甜菜各表型特征存在较复杂的联系,还要通过聚类分析,获得相关指标间相关性或差异性。
以欧氏距离和最长距离法对甜菜相关指标成分进行聚类分析,如下图5所示,当欧氏距离为0.7时,可将指标19种分成5类群。其中A类群包括4个指标,占总指标成分的21.05%,分别为叶绿素a、叶绿素b、叶绿素总浓度以及Tr。B类群包括3个指标,占总指标成分的15.78%,分别为根长、叶宽、地上生物量。C类群只有GS一个指标,占5.26%。D类群有9个指标,占总指标成分的47.36%,该类群的指标成分含量高于其他类群,分别包括地上部分高、Pn、Gs、叶面积、叶长、根冠比、地下生物量指标。E类群包括两个指标,分别为SPAD、NR。
综上所述,5个类群构成了整个植株的生长以及养分运输的整个过程。其中,从图5、表3可以看出,A、B、C类群与D、E类群呈负相关关系,且D、E类群占主导,表明光合特性的相关指标、氮效率以及植株的叶面积在甜菜生长过程中发挥重要作用。再者,从A类群可以看出叶绿素亦是不可缺少的指标。聚类分析较好的体现了不同指标间相关差异性,为研究甜菜生长过程中主要影响植株生长指标提供了一定的数据和理论支撑。
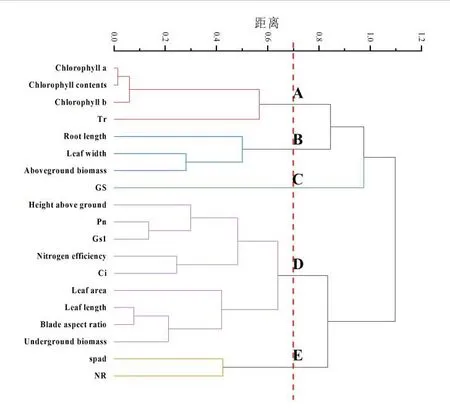
图5 聚类分析图
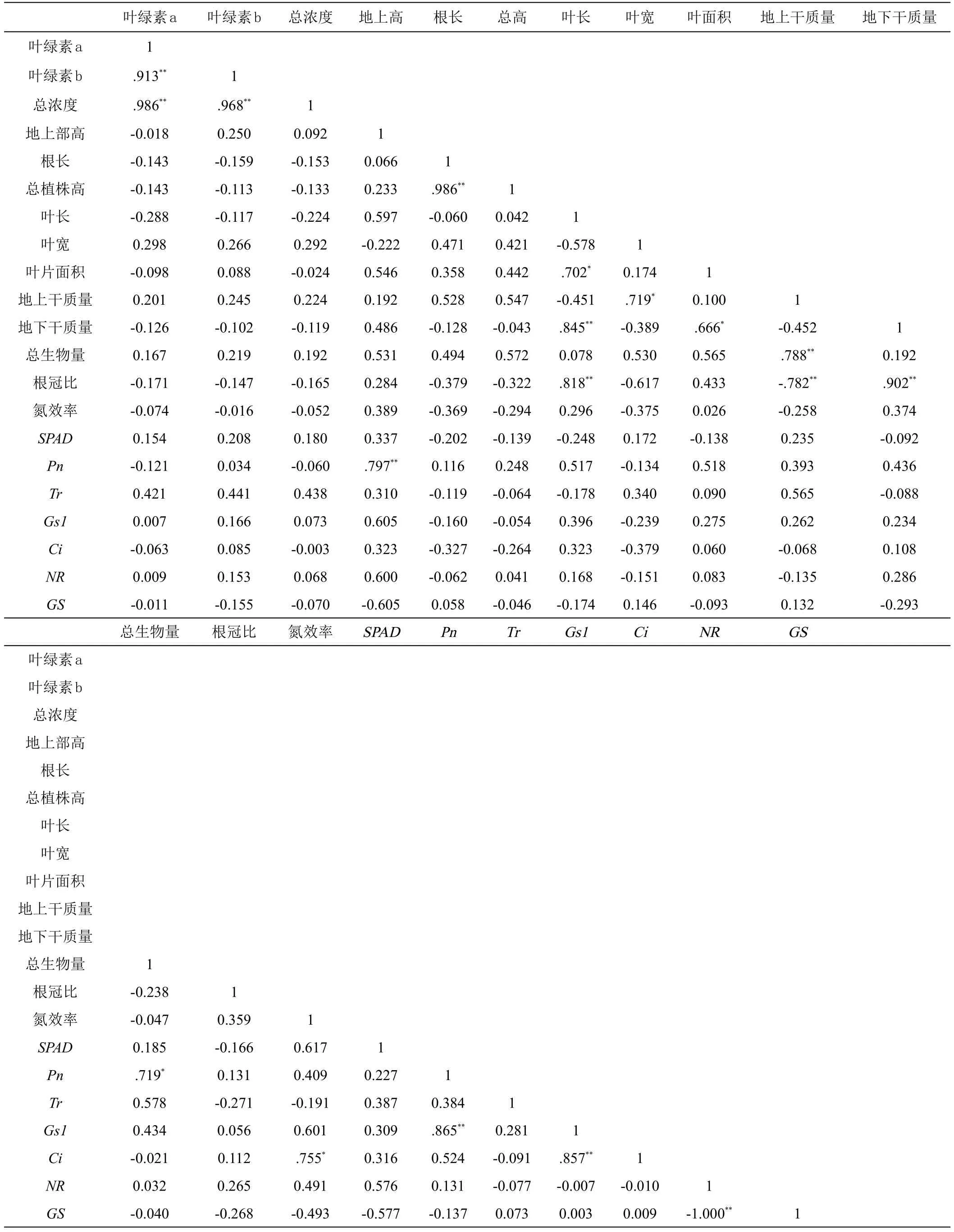
表3 接种Colpoda reniformis下甜菜幼苗的综合指标相关性分析
2.2.3 主成分分析 为了了解相关指标在甜菜生长发育过程中起的贡献作用,对甜菜幼苗的基表型特征指标及相关理化指标进行主成分分析,主成分见图6,6个成分特征值大于1,所以提取出6个主成分,累积贡献率为93.43%,其分别为30.65%、21.79%、16.19%、10.15%、9.9%和4.4%,这6个成分基本代表了这甜菜19个指标的绝大数信息。
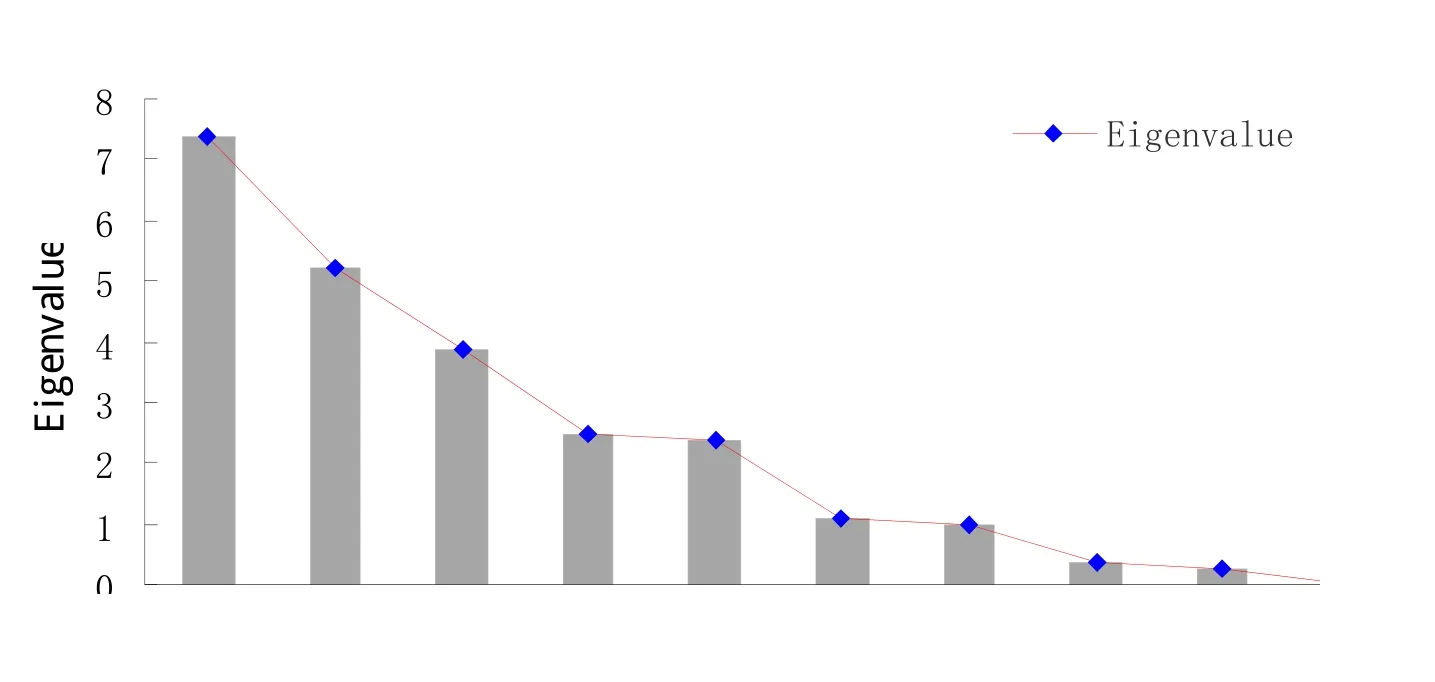
图6 主成分分析
由图7-a、7-b可知,第一主成分贡献率为30.65%;叶长、叶片长宽比、含氮量、地上部高、根冠比、地下部分干质量、氮效率、叶长叶柄比、根冠比、Pn、Gs的因子荷载值占比最高,所以第一成分中可对接种肾形肾形虫的甜菜生长情况、地下干质量积累以及光合特性进行初步的了解。第二主成分贡献率为21.79%;总生物量、地上部分干质量、Tr、叶宽的因子荷载值占比最高;结合表4,由于总生物量值与地上部分质量值呈显著正相关(P<0.01),且地上部分质量与叶宽呈正显著相关二者(P<0.05),所以第二主成分主要是植株生物量的积累。第三、四主成分贡献率分别为16.19%、10.15%;见图7-c、7-d,根长、植株高、总浓度、SPAD、叶绿素a、叶绿素b、叶片面积的因子的荷载占比最高,所以这两主成分概括了光合色素的含量、叶面积、根长及植株的高度的情况。第五主成分贡献率为9.9%,由图e、f可知,Gs、NR、GS、Ci荷载占比高,从表格看出,Gs与Ci显著性相关,NR与GS显著性负相关,所以第五成分可以看出叶片的呼吸强度及其酶活性的情况;第六成分贡献率是6个成分中最少,为4.4%,其中根长指标荷载值占比较高。
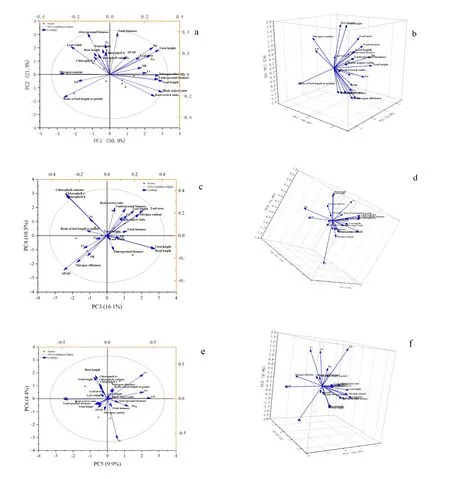
图7 接种Colpoda reniformis下甜菜幼苗所有指标主成分分析
3 讨论与结论
原生动物的活动不仅影响根际生物种群的代谢活性和繁殖发展,而且促进养分循环[6],从而影响植株的生物量、根系形态、植物激素等植物生理机制[31],更有利于植物的生长和生态系统的发展[32]。研究表明原生动物能够加强植株根系对土壤物质养分的吸收进而增加生物量的积累,促进植株的生长[33]。本研究发现接种肾形肾形虫后甜菜幼苗的总叶面积、根长、地上部分生物量、地下部分生物量以及总生物量显著增加,分别增加了15.49%、20.09%、10%、33.3%及14%,这也与BONKOWSKI等[13]和GUO等[34]的研究结果类似。通过提高植株的光合能力从而积累更多的生物量使得植株更好地生长[35]。本研究中,接种肾形肾形虫增加了叶绿素的含量,增加了叶面积,从而提高了光合效率[36],促进了植株生长以及生物量的积累[18,19]。
另外,本研究发现接种肾形肾形虫后甜菜的根长显著增加,并且根干质量和根冠比显著增加(图1),这表明肾形肾形虫对甜菜幼苗地下部分的贡献较大。甜菜根部接种肾形肾形虫后,肾形肾形虫的取食活动会对根系的生长产生刺激作用[37],且促进根系形态结构的发展[38],为植物获取了更多的养分以供植物的营养生长[39],因而增大植株地上部的叶面积,从而增强植株光合作用,获得更多的同化产物向地下转运来增加植株块根发育和物质积累[40]。
与形态和生物量变化相适应,光合作用在农作物干物质积累和产量方面具有重要作用,是必不可少的[41],且作物产量与光合的强度是息息相关的[33],所以两者之间具有一定有的相关性[42]。本研究结果发现甜菜的叶片面积、Pn、Gs在接种肾形肾形虫后效果明显增加,分别增加15.49%、43.91%、40.56%,并且叶绿素a、叶绿素b、总叶绿素也分别增加了31.58%、31.82%、31.72%。另外,SPAD值也得到了增加[43],此类观点在左亚男探讨蚯蚓粪对草莓植株的生长的研究有点类似[44]。叶绿素在植物光合中起着主导的作用[16],其含量的变化不仅影响植株的物质积累,也对植株的光合能力产生直接影响[45],其亦是植物光合能力及植株生理状况的一个参照因子[46]。叶绿素a在光合色素中的占据主导,反映作物对长波光的吸收程度,兼具吸收和转化光能的作用,大部分叶绿素a用于吸收光能[47]。叶绿素b在整体中起到辅助作用[48],其是在叶绿素a的基础上扩大了植物吸收光能的范围[16,49]。本研究中看出叶绿素含量的提高的确可以加强植物进行光合作用的能力,并且也发现接种肾形肾形虫后的甜菜叶片面积明显增大,达到了22.67 cm²,相比对照组增加了15.49%。叶片面积增大,则其对光的吸收面积亦变大,这也就进一步加大了甜菜植株相应的同化能力和物质积累。从研究结果来看,甜菜幼苗的形态、生理特征在肾形肾形虫的一定干预下是可以得到促进作用的,并且植株的生物量也得到了显著增加。
此外,接种肾形肾形虫后,甜菜的谷氨酰胺酶(GS)的酶活性显著增强,增加了23.52%,且氮素的干物质效率及氮累积量也显著提高,植株的地上部分和地下部分分别增加6.3%、8.8%和5.3%、19.7%。这表明GS的提高促进了甜菜的氮代谢的合成与转运,这在周晓明[50]和熊淑萍等[51]的研究的结果得到证实。谷氨酰胺合成酶(GS)在氮素代谢途径中必不可少,对植物氮素吸收和利用有直接作用,其对植物的生长发育和产量亦产生重要作用[52]。OLIVEIRA等[53]将大豆的GS1基因超表达到烟草中提高了转基因株系的生物量,并且发现GS酶活力和植株生物量是明显相关的。孙永健等[54]研究发现GS活性可作为植株各生育期氮素积累量指标的判断。其次在本研究中发现NR的酶活性的增加并不是太显著,可能培养的时间太短,这些结论仍然需要进一步研究和证明。
综上所述,肾形肾形虫可以干预甜菜幼苗的生长发育和生理情况,且具有促进作用。与未接种肾形肾形虫处理相比,接种肾形肾形虫明显增强了甜菜的光合作用,增加了植株的生物量积累,并且肾形肾形虫的干预对甜菜地下部分生长的影响是比较明显的。这些结果说明接种肾形肾形虫后对甜菜幼苗地上部分和地下部分都有一定积极影响,表明原生动物的活动干预可能在刺激植物生长中发挥重要作用,对以后甜菜在生物型肥料方面的增产、高效提供理论依据,具有重要的生态意义。未来的农业实践亦可能旨在专门提高植物有益的原生生物,或者将这些原生生物作为新型的、可持续的生物肥料来应用,实现农业的可持续发展。
猜你喜欢
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
今日农业(2020年19期)2020-12-14
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
中国糖料(2016年1期)2016-12-01
中国糖料(2016年1期)2016-12-01
中国科技信息(2015年2期)2015-11-16
食品工业科技(2014年6期)2014-05-10
