
夏季庐山常绿阔叶林4种常见植物根际AM真菌分布特征
2022-02-21 11:04吴佳海李金苗
中国农学通报 2022年36期
吴佳海,邹 芹,杜 曲,张 涛,李金苗,肖 斌,刘 玮,3
(1江西农业大学林学院/园林与艺术学院,南昌 330045;2嘉兴济兴环境科技有限公司,浙江 嘉兴 314000;3江西庐山森林生态系统定位观测研究站,江西 九江 332900;4江西庐山国家级自然保护区管理局,江西 九江 332900;5湛江科技学院,广东 湛江 524084;6中国林科院亚热带林业实验中心,江西 分宜 336600)
0 引言
庐山地理位置独特,常常被认为是一个重要的生物避难所,其森林覆盖率高,植被也较丰富,包括不少稀有、孑遗物种,具有较高的生态功能和价值[1]。建国前,由于人工采伐等原因,作为庐山低海拔较为常见的常绿阔叶林受到了严重的破坏,但是从20世纪40—50年代保护至今,庐山又逐渐恢复了以石栎(Lithocarpus glaber)、甜楮(Castanopsis eyrei)等为建群种的常绿阔叶林植被,既具有地带性特点,又具有地域性特征,这在中亚热带地区是极为少见的[2]。然而,在庐山常绿阔叶林中,人们对地下生态过程的重要参与者AM真菌,知之甚少。因此,对庐山常绿阔叶林土壤AM真菌群落及其分布特征的研究,不仅可以推动其土壤AM真菌的开发和利用,还可以推动人们对其地下生态过程的了解。AM真菌是一类古老的内生真菌类群,能与约80%陆地高等植物形成共生关系,在个体角度上,AM真菌可以促进植物生长并提高其抗病虫害、抗干旱等能力[3-6];在森林生态系统中,AM真菌可以提高树木幼苗的存活率,也可以促进树木生长与提高树木初级生产力,还可以推动退化森林的恢复与维持植被群落多样性[7-10]。自从1844年TULASNE兄弟通过AM真菌孢子形态特征描述开始,人们对AM真菌的种群及其分类、分布特征探究一直在延续,同时在前扑后继工作者的共同努力下,AM真菌的分类系统不断地得到扩充和完善[11]。2019年以来,基于孢子形态学分析法,研究AM真菌多样性和分布特征的文章也时有报道[12-13]。但是,目前以形态学方法探索庐山常绿阔叶林土壤AM真菌群落及其分布特征的研究较为罕见。结合形态学鉴定法,对庐山常绿阔叶林土壤AM真菌群落及其分布特征进行研究,以期为其土壤AM真菌资源开发与利用提供理论基础。
1 材料与方法
1.1 研究地点概况
庐山位于长江中下游鄱阳湖平原西北部,属断块山构造地貌、冰蚀地貌、流水地貌三位一体的复合型地貌景观,面积302 km2,海拔跨越40~1474m,属中亚热带东部湿润季风气候,同时又具有山地气候的特征。庐山年平均气温11.4°C,年平均降水量1929.2mm,降水主要集中与4—7月。
庐山常绿阔叶林样地位于庐山南山伴耳峰区域(115°54′23.39″E,29°31′05.28″N),样地大小为100m×100m,海拔402m。在庐山常绿阔叶林中,乔木建群种常有变化,像本样地中从东及西,壳斗科(Fagaceae)的甜楮(C.eyrei)、石栎(L.glaber),樟科(Lauraceae)的樟树(Cinnamomum camphora)均有所分布,但是分布并不均匀,并且这几种建群乔木根系分布范围较广,在山坡地上进行根际土取样较为困难。但老鼠矢(S.stellaris),檵木(L.chinensis),山胡椒(L.glauca),连蕊茶(C.fraterna)这4种植物常绿阔叶林样地中分布较为均匀、普遍,为常见树种。
1.2 取样及处理方法
1.2.1 取样方法 2018年6月,在常绿阔叶林样地中设置4个20m×20m的小样方进行取样,其中在每个20m×20m的小样方种选取常见的老鼠矢,檵木,山胡椒和连蕊茶4种植物,每种植物随机选择5株个体,共计80株个体,并在土壤表面清理枯枝落叶后随主根挖掘、采集其在0~30 cm土层的根际土样(1~2 kg)以及根系样品。另外,样品是以四方位取样法取得,其中土样样品分为2部分,一部分风干保存,另一部分置于-20ºC冰柜冷藏保存;其中根系样品则用流水冲净后置于-20ºC下冷藏保存。
1.2.2 AM真菌孢子形态鉴定方法
(1)使用改进的湿筛倾析法分离和获取土壤中的AM真菌孢子[14-15];
(2)孢子形态鉴定依据INVAM(http://invam.caf.wvu.edu/Myc-Info/)、http://www.zor.zut.edu.pl/Glomeromycota/index.html和《球囊菌门丛枝菌根真菌最新分类系统菌种名录》中AM真菌的分类与描述[16]。
1.2.3 AM真菌群落多样性指标计算方法 土壤AM真菌多样性特征采用下列指标代表:
(1)孢子密度(Spore density,SD)为10 g风干土中AM真菌的孢子数量、物种丰富度(Species richness,SR)、Pielou 均匀度指数(Pielou evenness index,J)等按照吴佳海等[15]的方法计算。
(2)相对多度(Relative abundance,RA)、分离频率(Isolation frequency,IF)、重要值(Importance value,IV)根据钟凯和董怡然的方法计算[17-18],其中IV<10%,则为稀有种,10%≤IV<30%则为常见种,30%≤IV<50%,则为广布种;IV≥50%,则为优势种。
(3)Shannon-Wiener指数(Shannon-Wiener index,H')根据春蕾和峥嵘的方法计算[19]。
1.2.4 土壤养分特征测定方法 测定的土壤因子有pH、氨态氮(Ammonium nitrogen,NH4+-N)、硝态氮(Nitrate nitrogen,NO3--N)、全氮(Total nitrogen,TN)、全磷(Total phosphorus,TP)、全钾(Total potassium,TK)。所有指标均按照鲍士旦的方法进行测定[20]。
1.2.5 植物根系AM真菌侵染特征测定 采用改进的醋酸墨水法测定植物根系AM真菌侵染特征,计算公式如式(1)所示[21-22]。

1.3 数据分析
采用单因素方差分析比较不同植物根际AM真菌孢子密度、物种丰富度以及土壤养分之间的差异,数据采用SPSS 22.0(SPSS Inc.,Chicago,IL)进行统计。相关性系数用于分析植物根际AM真菌孢子密度与物种丰富度之间的关系以及两者与土壤养分、水分之间的关系。Cannoco 5.0(CANOCO for Windows v.5.0)用于典范对应分析Canonical Correlation Analysis(CCA)分析,解释AM真菌群落分布特征与土壤养分特征之间的关系。
2 结果与分析
2.1 常见植物根系侵染特征
4种植物根系均有AM真菌侵染,其中侵染率从高到低依次为连蕊茶(24.14%)、檵木(23.65%)、老鼠矢(21.68%)、山胡椒(21.53%),且不同植物根系侵染率之间存在显著差异(图1)。
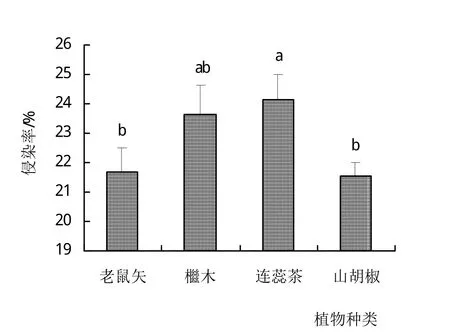
图1 庐山常绿阔叶林常见植物根系AM真菌侵染特征
优势种和广布种AM真菌的孢子密度与侵染率呈正相关,但均不显著(R=0.095,P=0.447;R=0.213,P=0.084);常见种和稀有种AM真菌的孢子密度与侵染率呈负相关,也均不显著(R=-0.090,P=0.467;R=-0.071,P=0.569)。优势种和广布种AM真菌的物种丰富度与侵染率呈正相关,但均不显著(R=0.058,P=0.646;R=0.201,P=0.103);常见种和稀有种AM真菌的物种丰富度与侵染率呈负相关,也均不显著(R=-0.122,P=0.324;R=-0.082,P=0.511)。
2.2 常见植物根际AM真菌群落组成
通过孢子形态鉴定方法,从庐山常绿阔叶林4种常见植物根际共分离出AM真菌7属44种,包括球囊霉属(Glomus)19种,无梗囊霉属(Acaulospora)16种,双型囊霉属(Ambispora)5种,根孢囊霉属(Rhizophagus)、近明球囊霉属(Claroideoglomus)、硬囊霉属(Sclerocystis)与多样孢囊霉属(Diversispora)各1种,其中球囊霉属(Glomus)和无梗囊霉属(Acaulospora)为优势属(表1)。同时,根据优势度可将44个AM真菌物种划分为4个优势种、5个广布种、12个常见种和23个稀有种,其中优势种均属于球囊霉属;而稀有种在7属AM真菌中均有分布(图2)。

表1 庐山常绿阔叶林常见植物根际AM真菌的分布特征 %
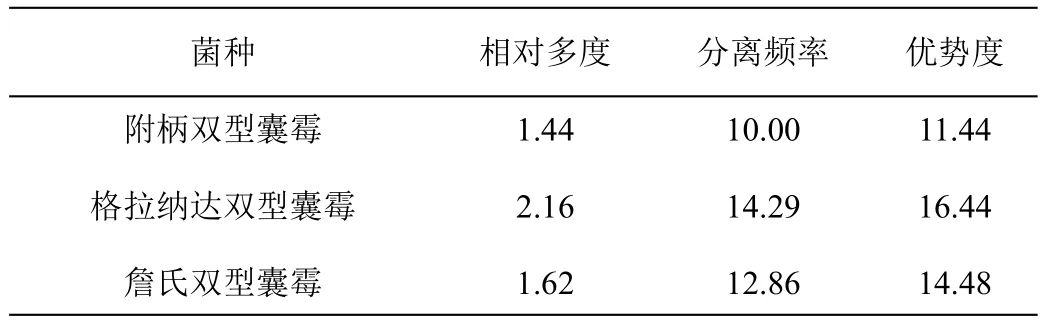
续表1
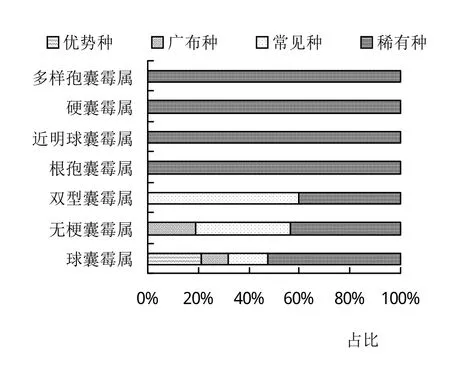
图2 常见植物根际AM真菌群落组成
2.3 常见植物根际AM真菌分布特征
4种植物根际AM真菌孢子密度不存在显著差异,但物种丰富度存在显著差异(表2)此外,优势种、广布种、常见种和稀有种AM真菌的孢子密度之间均存在负相关,但只有优势种AM真菌的孢子密度和常见种、稀有种AM真菌的孢子密度存在显著负相关(R=-0.273,P=0.023;R=-0.268,P=0.026);同时,优势种、广布种、常见种和稀有种AM真菌的物种丰富度之间均存在负相关,但均不显著(P>0.05)。

表2 常见植物根际土壤AM真菌分布特征
4种植物根际AM真菌相对多度和分离频率存在极显著相关(R=0.991,P<0.01)。
由图3和图4可知,在优势种AM真菌方面,只有檵木根际优势种AM真菌的孢子密度显著高于山胡椒,同时只有山胡椒根际优势种AM真菌的物种丰富度显著低于其他植物;在广布种AM真菌方面,只有老鼠矢根际广布种AM真菌的孢子密度显著低于连蕊茶和山胡椒;在常见种AM真菌方面,只有老鼠矢根际常见种AM真菌的物种丰富度显著高于山胡椒;在稀有种AM真菌方面,只有山胡椒根际稀有种AM真菌的孢子密度显著高于檵木和连蕊茶。
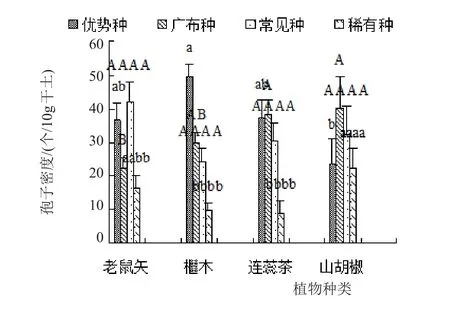
图3 植物根际不同优势度AM真菌孢子密度之间的差异
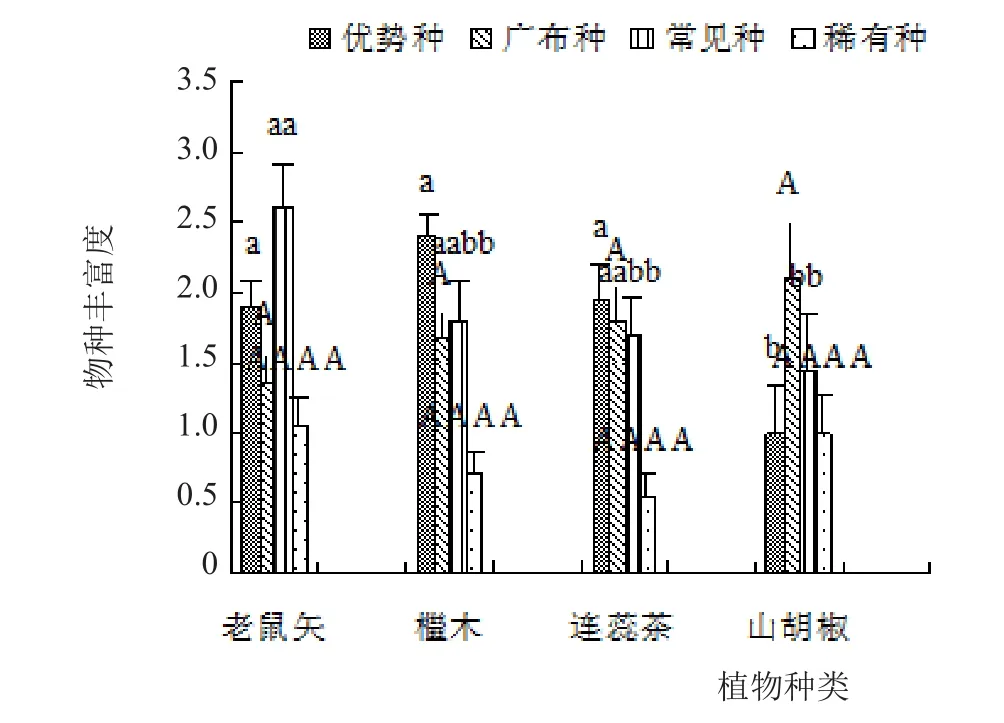
图4 植物根际不同优势度AM真菌物种丰富度之间的差异
除老鼠矢外,其他植物根际优势种和广布种AM真菌的孢子密度之和与物种丰富度之和均高于其根际常见种和稀有种AM真菌的孢子密度之和与物种丰富度之和(图3和图4)。
2.4 常见植物根际AM真菌群落影响因素
不同植物的根际土壤中,除了全氮、氨态氮和水分存在显著差异外,其他土壤养分均不存在显著差异(表3)。
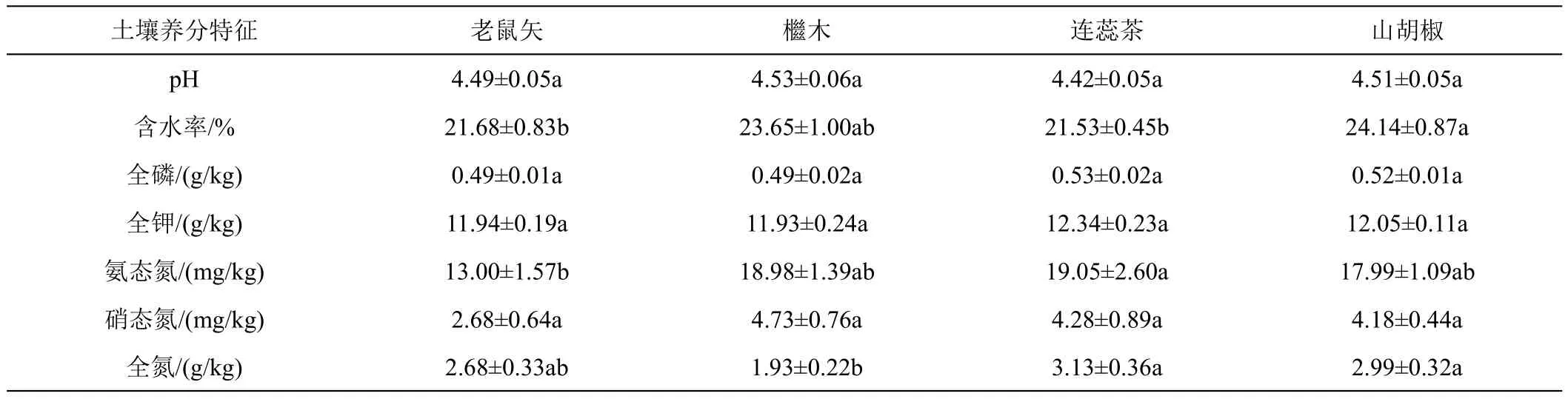
表3 常见植物根际土壤养分情况
图5为土壤养分、水分与AM真菌群落分布的典范对应分析图,其中第一主轴解释3.14%,第二主轴解释了5.63%,总共解释了8.77%。由图5可知,土壤养分(全氮、氨态氮、硝态氮、全磷、全钾)、pH和水分对AM真菌群落影响较弱;不同优势度AM真菌受土壤养分、pH和水分影响程度不同,受影响程度从高到低依次为稀有种、常见种、广布种和优势种。
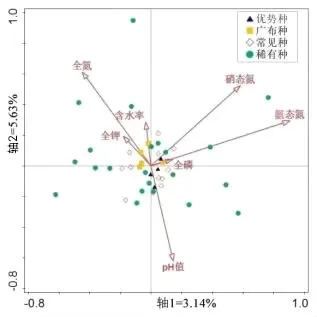
图5 庐山常绿阔叶林常见植物根际AM真菌分布对土壤养分、水分响应的典范对应分析图
3 结论
本研究在AM真菌形态学鉴定的基础上,引入了AM真菌优势度的概念,并将不同优势度AM真菌的分布特征为研究出发点,发现在夏季庐山常绿阔叶林中,常见植物与AM真菌的共生模式可能是以优势种AM真菌和广布种AM真菌共生为主,以常见种AM真菌和稀有种AM真菌补充共生的共生模式;在该共生模式下,植物根际AM真菌资源较为丰富,以球囊霉属和无梗囊霉属为优势属且分布不均;AM真菌与植物共生程度由不同优势度AM真菌共同决定;不同优势度AM真菌之间相互影响较弱;AM真菌分布受土壤养分(全氮、氨态氮、硝态氮、全磷、全钾)、pH和水分影响较小,但不同优势度AM真菌受土壤养分、pH和水分影响程度不同。
AM真菌也会受季节影响,因此,在春季、秋季和冬季中,常见树种与根际AM真菌的共生方式是否与夏季一致;同时,该共生模式仍需要进一步通过盆栽试验进行验证。
4 讨论
众所周知,AM真菌广泛分布于自然界中,能与陆地上约80%的高等植物形成互惠共生关系[6,23]。同时,AM真菌有着丰富的生态功能,在促进植物生长和植物群落演替等方面都起着重要作用[24-25]。同时侵染率是表示AM真菌与宿主植物共生程度的一个重要指标。然而关于侵染率,在研究探索过程中,不同学者确有不同看法,其中姜攀和宰学明[26-27]的研究均认为AM真菌孢子密度与AM真菌侵染率不存在显著相关,而贺学礼等[28]研究发现AM真菌孢子密度与AM真菌侵染率存在显著相关,而本研究的结果与姜攀和宰学明[26-27]的研究结果一致。可能产生的原因是:第一,有些AM真菌物种处于幼年阶段,还不具备侵染植物的能力;第二,不同优势度的AM真菌对侵染率的影响不同,有的AM真菌有利于提高植物根系侵染率,而有的AM真菌有利于降低植物根系侵染率,从而导致总的AM孢子密度与侵染率不存在显著相关。同时,本研究也发现优势种和广布种AM真菌的孢子密度和物种丰富度均与侵染率存在正相关,而常见种和稀有种AM真菌的孢子密度和物种丰富度则均与侵染率存在负相关。这一结论印证了上述第二点猜想,但是各优势度AM真菌的孢子密度和物种丰富度均与侵染率均不存在显著相关。原因可能跟上述的第一点猜想一致。同时,本研究还发现除了老鼠矢外,其他植物根际优势种和广布种AM真菌的孢子密度之和与物种丰富度之和均高于常见种和稀有种AM真菌的孢子密度之和与物种丰富度之和。因此,综合上述2个结果猜测,在夏季的庐山常绿阔叶林中,AM真菌与常见植物的共生模式可能是以优势种和广布种AM真菌为主共生,常见种和稀有种AM真菌为补充共生的共生模式。
AM真菌分布受到众多生物因素和非生物因影响的同时也受AM真菌自身产孢能力和传播能力等特性的影响[15,29]。本研究发现,球囊霉属和无梗囊霉属为优势属。这与袁腾等[30]和BONFIM等[31]的研究结果一致。球囊霉属与无梗囊霉属在各个生态系统中均较为常见,是AM真菌的两个大属,具有庞大的家族,还具有高的传播能力与高的适应能力[32]。因此,球囊霉属与无梗囊霉属AM真菌其庞大家族群落与较强的适应能力可能是其成为优势群落的原因。此外,本研究还发现,不同植物根际AM真菌孢子密度不存在显著差异,与陈志超等研究结果一致[33]。但植物根际不同优势度AM真菌的孢子密度却存在显著差异(常见种除外)。同时也发现优势种AM真菌的孢子密度和常见种、稀有种AM真菌的孢子密度存在显著负相关。因此,根据上述3个结果猜测,植物根际处于优势AM真菌的孢子密度可能被弱势AM真菌的孢子密度所稀释,最终导致AM真菌总孢子密度不存在显著差异。另外,本研究发现不同植物根际的AM真菌物种丰富度存在显著差异,与何斐等结果一致[34]。同时也发现不同植物根际优势种和常见种AM真菌的物种丰富度存在显著差异,并且不同优势度AM真菌物种丰富度之间不存在显著相关。所以,不同植物根际AM真菌物种丰富度的差异是由优势种和常见种AM真菌引起。常见种AM真菌分离频率和相对多度均较低,其产孢能力和传播能力可能较弱,同时常见种AM真菌可能更加容易土壤养分的影响。因此,常见种AM真菌能引起不同植物根际AM真菌物种丰富度差异。然而,优势种AM真菌既有高的分离频率又有高的相对多度,其孢能力和传播能力均较强,不易引起差异;同时本研究中优势种AM真菌的物种丰富度与其他优势度AM真菌的物种丰富度之间不存在显著相关,即优势种AM真菌对其他优势种AM真菌的影响程度较低。因此,优势种AM真菌不是通过自身特性和影响其他优势度AM真菌群落来影响宿主植物根际AM真菌总物种丰富度,那么优势种AM真菌是如何引起宿主植物根际AM真菌物种丰富度的差异,值得深思。
本研究中,土壤养分、pH和水分对不同优势度AM真菌分布的影响程度不同,影响从大到小依次为稀有种、常见种、广布种和优势种。这个结果证实了上述关于常见种AM真菌的第二个猜想。PORRASALFARO和DIEPENET研究发现氮可以影响AM真菌的营养过程,还可以改变AM真菌的群落结构[35-36]。在本研究中,土壤氮对AM真菌群落的影响相对较高,高于其他土壤养分、水分,这与XU的研究结果相似[37]。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
中国土壤与肥料(2021年5期)2021-12-02
中国瓜菜(2020年8期)2020-09-26
少儿美术(快乐历史地理)(2019年4期)2019-02-21
天然产物研究与开发(2018年3期)2018-05-07
岷峨诗稿(2017年4期)2017-11-25
中华诗词(2017年11期)2017-04-18
中国蔬菜(2016年8期)2017-01-15
中国烟草学报(2012年5期)2012-04-12
