
喜德县中坝村火烧迹地植被恢复初期优势种生态位与种间联结*
2022-02-19 03:27刘鲁光朱兆棋陈曦刘守江
西部林业科学 2022年1期
刘鲁光,朱兆棋,陈曦,刘守江
(1.西华师范大学 生命科学学院,四川 南充 637009;2.西华师范大学地理科学学院,四川 南充 637009;3.西华师范大学 四川省干旱河谷土壤侵蚀监测与控制工程实验室,四川 南充 637009)
植物群落是一定区域内植物与环境的有机结合体,群落各物种间的相互关系决定着群落的结构、功能及演替方向[1]。对群落物种生态位的研究主要涉及生态位宽度与生态位重叠两个方面[2]。生态位宽度是指生物在群落中所能利用的资源总和,生态位重叠反映的是不同物种对同一资源利用的共性,是判断物种间生态相似性与竞争强度的重要参考[3]。种间联结可以表达不同物种之间在空间分布上的相互关系[4],是一种静态的描述[5]。主要有基于物种是否存在的二元数据的定性描述与基于物种数量数据的定量描述两种方式[6]。基于生态位和种间联结的研究,可以了解群落物种间的相互作用、揭示群落物种与环境因子间的关系,在分析群落结构与功能[7]、测度群落稳定性[8]、判断群落演替趋势[9]等方面也得到了广泛应用。生态位与种间联结研究也因此成为当今研究的热点[10]。
林火是对植物群落结构和演替过程影响最大的干扰之一[11],也是当今人类所面对的全球性问题之一。林火会破坏植被,导致生态系统退化,加重当地的水土流失[12]。凉山州是四川省土壤侵蚀监测与恢复治理的重点地区,土壤侵蚀现象十分普遍,生态环境较为脆弱[13]。近年来,凉山州山火频发导致区域内植被严重退化,进一步加剧了相关区域的生态脆弱性。植物群落在保持土壤水分、水土保持、调节区域的小气候以及改善土壤质量等方面有重要作用[14]。同时,火后植被的恢复过程是一个长期复杂的动态过程[15],植被恢复状况对土壤侵蚀强弱起着决定性的作用[16],因此该区域的植被恢复成为灾后生态修复与水土保持工作的重要内容。已有大批学者对干热河谷[17]、黄土高原[18]、砒砂岩区[2]、喀斯特地貌区[9]等生态脆弱区植物群落的生态位及种间联结进行了研究,这些研究均指出生态位与种间联结研究对生态脆弱区的生态恢复与植物保护具有重要的实践意义。
2020年5月7—12日,喜德县鲁基乡中坝村发生森林火灾,过火面积约28.6 hm2。林火导致中坝村植被受损严重,生态系统严重退化,并于6月4日由强降雨引发泥石流次生自然灾害,严重威胁附近居民的生命财产安全。大批学者对火烧迹地群落恢复过程中的物种组成、植物多样性、群落稳定性等指标进行了研究[19],但现有研究中对林火干扰后植物群落生态位与种间联结的关注还较少。本研究于2020年8下旬对喜德县中坝村火烧迹地进行调查,以凉山州喜德县火烧迹地恢复初期植物群落的优势物种为研究对象,利用物种分布的一元和多元数据,对群落优势种的生态位宽度、生态位重叠、种间联结的状况进行分析。以此来量化物种之间及物种与环境间的适应与竞争状况,进而揭示火烧迹地植被恢复初期群落物种的空间分布特征、群落结构、生境异质性及演替趋势等状况,以期为火烧迹地后续的生态恢复与植物保护实践提供理论参考。
1 材料与方法
1.1 研究区概况
研究区位于四川省凉山州喜德县(102°11′31.20″~102°43′1.19″E,27°53′6.72″~28°31′8.76″N),山脊呈东西走向,土壤为酸性土,总面积2 210.428 km2。该区域属于亚热带季风气候区,气候受区位因素的影响,有低纬度高海拔的特点。夏季凉爽多雨,冬季干燥温和。年平均气温14.1 ℃左右,1月最冷,月平均气温5.5 ℃左右;7月最热,月平均气温21 ℃左右。多年平均降水量1 006 mm,降水的季节分布不均匀,80%以上的降水量发生在5月至10月。火灾发生前乔木以赤松(Pinusdensiflora)、青冈(Cyclobalanopsisglauca)为主,赤松在林火中损毁严重,调查时未发现自然更新的乔木幼苗。
1.2 数据来源
在对喜德县中坝村火烧迹地全面探查的基础上,于火灾后3个月即2020年8月,在火烧迹地选择未发生明显侵蚀的山体、侵蚀沟、堆积体3种当地常见地形区分别设置17个、9个、4个规格为5 m×5 m的固定植物群落样方,便于后期进行持续跟踪观测。由于原有乔木在山火中损毁严重,调查时的恢复状况极差,因此本调查未设置乔木样方。对样方中灌木、草本植物的物种名、植株数、盖度、最大高度等进行调查记录,同时测定样方的海拔、坡度、坡向,样方的详细分布见图1。
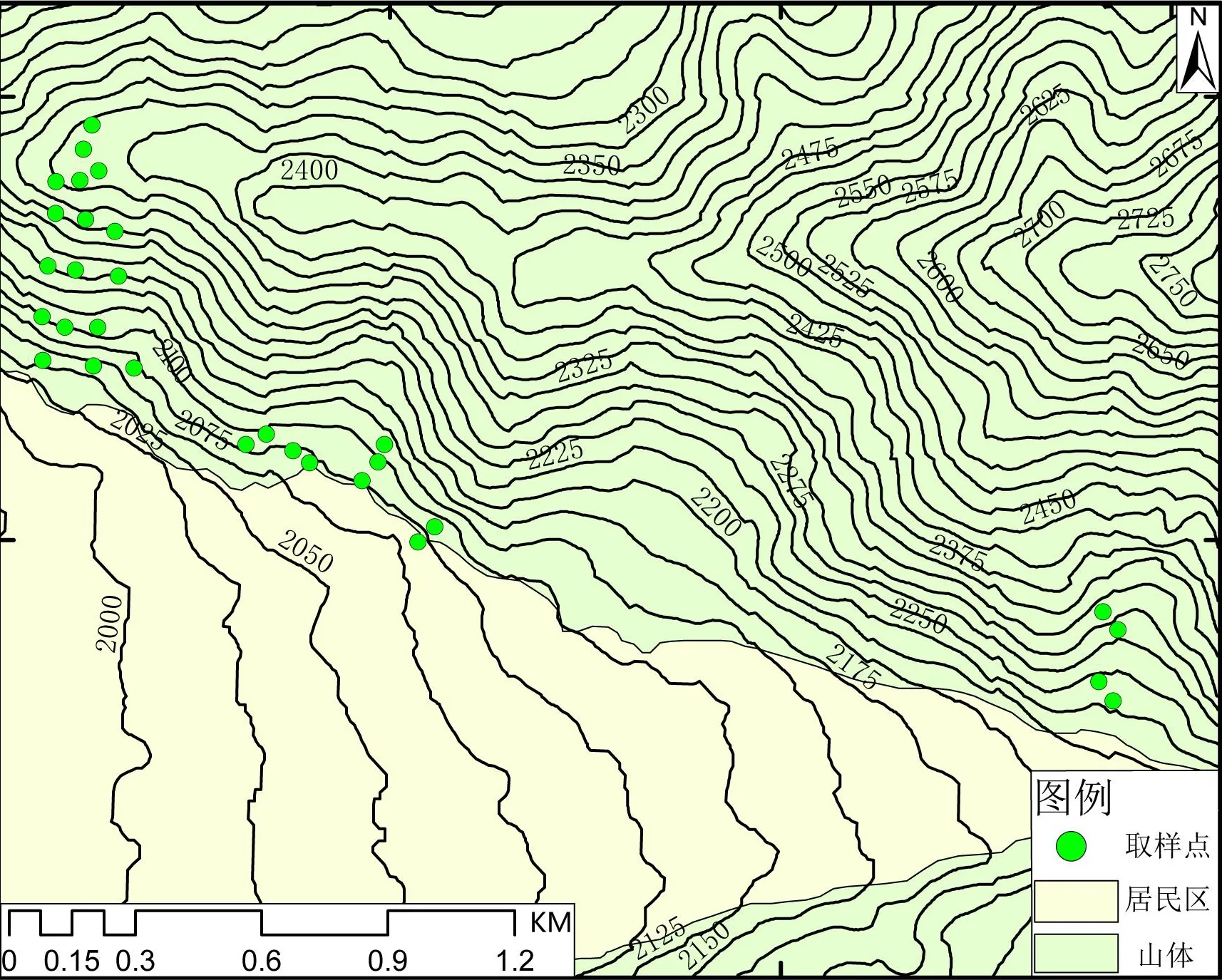
图1 研究区域样点分布Fig.1 Overview of the study area
1.3 数据处理
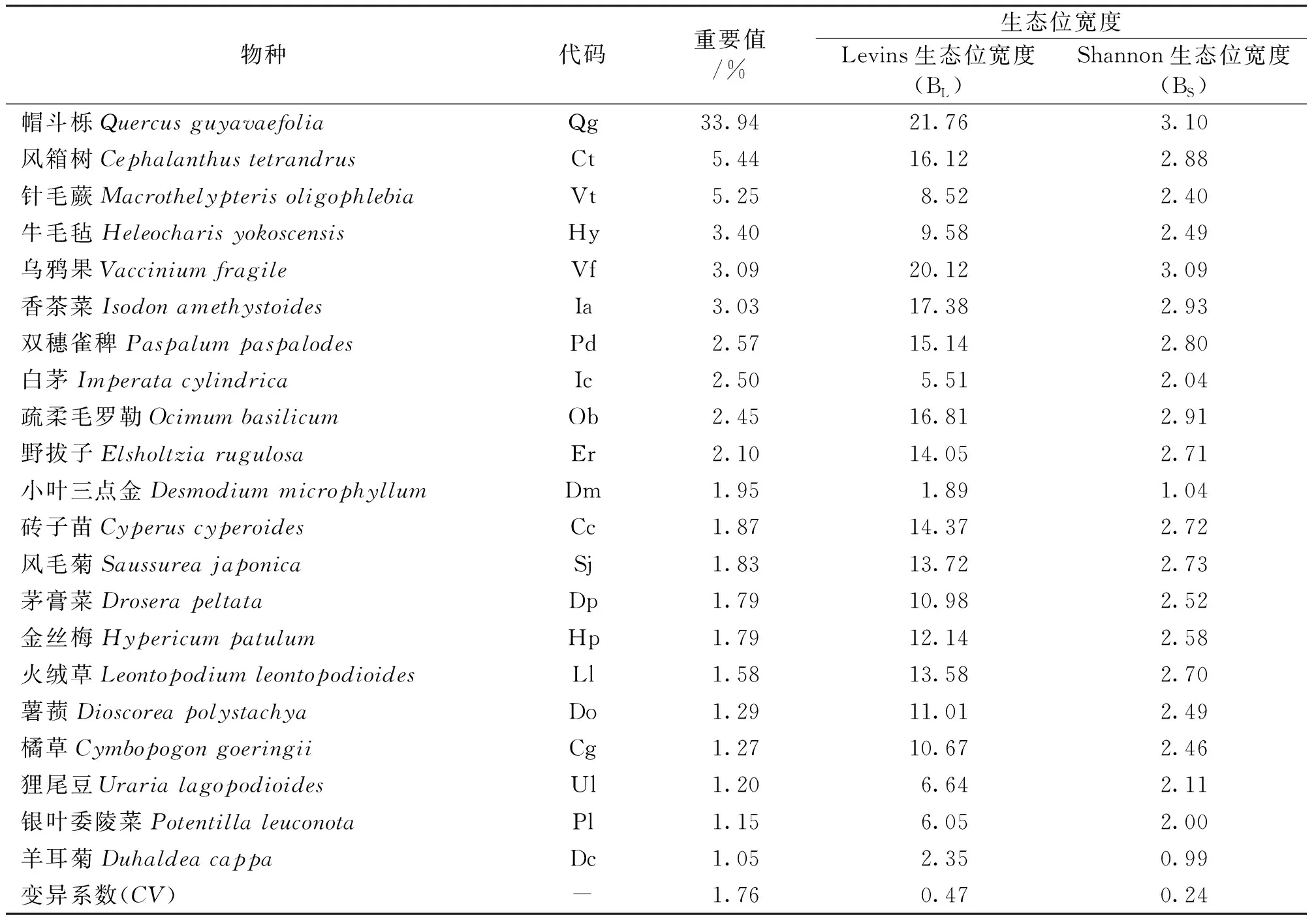
根据实地踏查及计算结果,结合前人的经验[20-21],选取重要值大于1%的21个物种(重要值之和占全部物种的80%)作为植被恢复初期群落的优势种进行生态位与种间联结研究。采用Excel 2016对野外调查的数据进行整理及重要值计算,使用R软件中的种间关联分析spaa包[22]进行群落物种的生态位及种间联结计算,使用R软件中corrplot[23]及arcgis 10.6进行绘图。
1.3.1 重要值计算
重要值(IV)可表示物种在群落中的重要程度,公式如下[24]:
IV=(相对密度+相对频度+相对盖度)/3
1.3.2 生态位计算
式中,Pij和Pkj分别表示物种i和物种k在样方j中的重要值,N为样方总数。
1.3.3 群落总体联结性检验
采用Schluter提出的方差比率法(VR)来进行群落多物种间整体关联性测定,公式如下[10]。
W=N×VR
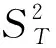
1.3.4X2检验
基于一元数据的物种间种间关联性计算使用经过Yates矫正的X2检验来进行计算,公式如下[24]。
式中,a为两物种共同出现的样方数,b、c为两物种中仅有一个出现的样方数,d为两物种均未出现的样方数。若ad=bc,则物种间无关联,若ad>bc,则呈正联结,若ad 1.3.5 Spearman秩相关与Pearson相关 利用多度数据的物种种间关联性采用Spearman秩相关(rs)与Pearson相关(rp)来进行计算,公式如下[24]: 式中,xij、xkj分别为物种i与物种k在样方j中的秩。 植被恢复初期群落中不同物种的重要值、BL、BS均存在较大差异(表1),变异系数分别达到1.76、0.47、0.24,这说明植被恢复初期各物种的分布不均匀,物种对环境的利用能力存在较大差异。其中,帽斗栎(Quercusguyavaefolia)BL、BS最大,分别为21.76、3.10,说明帽斗栎对火烧迹地环境的适应能力最强,在群落中占据优势地位。BL、BS两种生态位宽度的排序基本一致,但与重要值排序存在较大差异,如乌鸦果(Vacciniumfragile)的重要值的排序明显低于两种生态位的排序;针毛蕨(Macrothelypterisoligophlebia)的重要值排序明显高于两种生态位的排序。根据物种的生态位宽度和物种的习性可以将物种分为2类。第一类是对环境适应能力较强的泛化种,BL、BS分别介于(11.0,21.2)、(2.5,3.11),包括帽斗栎、乌鸦果、砖子苗(Cyperuscyperoides)等物种,这些物种的抗逆性强,能适应较大幅度的生境变化;第二类是生长发育对环境有特殊的要求的窄生态位物种,BL、BS分别介于(1.8,11.1)、(0.98,2.5),包括喜阴湿的牛毛毡(Heleocharisyokoscensis)、针毛蕨等,这些物种分布往往受某一个环境因子的限制或只能在特定的环境条件下才能生存。 表1 优势物种重要值与生态位宽度Tab.1 Importance value and niche breadth of dominant species 火烧迹地植被恢复初期各物种之间的Pianka生态位重叠(Oik)介于0~0.85之间(表2)。Oik=0的有8对,占总对数的3.8%,分别为风毛菊(Saussureajaponica)-小叶三点金(Desmodiummicrophyllum)、狸尾豆(Urarialagopodioides)-小叶三点金、狸尾豆-银叶委陵菜(Potentillaleuconota)、茅膏菜(Droserapeltata)-小叶三点金、乌鸦果-羊耳菊(Duhaldeacappa)、小叶三点金-羊耳菊、羊耳菊-针毛蕨、小叶三点金-白茅(Imperatacylindrica)。Oik≥0.8的有4对,占总对数的1.9%,分别为风毛菊-橘草(Cymbopogongoeringii)、金丝梅(Hypericumpatulum)-牛毛毡、帽斗栎-乌鸦果、风毛菊-疏柔毛罗勒(Ocimumbasilicum),这些物种可能具有较高的生态相似性,且物种间为相互竞争的关系。生态位重叠程度较低的0 表2 火烧迹地优势种的pianka生态位重叠Tab.2 Pianka niche overlap of dominant species in burned land 表3 火烧迹地优势种总体联结性检验Tab.3 Test on the overall association of dominant species in burned land X2检验结果显示(图2),植被恢复初期由群落21个优势种组成的210个种对中,有103对种间关系呈正联结,101对呈负联结,6对物种之间无明显的关联,分别占总种对数的49.05%、48.10%、2.86%。植被恢复初期群落物种间的正负关联比为1.02,只有极少数物种之间不存在相互关联,说明群落还不稳定,与VR检验的结果相符。所有种对中有32对的关联程度达到显著水平,占总种对数的15.2%。其中,有13对物种之间为显著正联结,6对物种之间为极显著正联结,分别占总种对数的6.19%和2.86%。呈极显著正联结的种对分别为火绒草(Leontopodiumleontopodioides)-野拔子(Elsholtziarugulosa)、金丝梅-双穗雀稗、金丝梅-野拔子、牛毛毡-针毛蕨、双穗雀稗(Paspalumpaspalodes)-针毛蕨、小叶三点金-银叶委陵菜。所有种对中负联结达到显著水平的有10对、达到极显著水的种对有3对,分别占总种对数的4.76%、1.43%。与风毛菊呈极显著负联结的均为牛毛毡、小叶三点金、银叶委陵菜这类植株矮小、喜阴湿环境的物种。植被恢复初期,多数样方的生境较差,抗逆性较强的风毛菊相对喜湿物种分布更加广泛,降低了相关物种出现在同一区域的概率;其次,即使出现在同一区域风毛菊的基生叶也占据了群落部分的下层空间,使其难于与地被植物共存。 图2 火烧迹地优势物种间的X2检验半矩阵Fig.2 X2 test semi matrix among dominant species in burned slash Spearman秩相关分析的结果显示(图3),火烧迹地21个优势种所构成的210个种对正负联结比为0.79。其中,呈正相关的有91对,占总种对数的43.3%;正相关达到显著水平的有14对,占总种对数的6.7%;正联结程度达到极显著水平的有26对,占总种对数的12.4%。所有种对中呈负联结的有115对,占总种对数的54.8%。其中,显著负联结(P<0.05)的种对有14对,占总种对数的6.7%;呈极显著负联结的种对有15对,占总种对数的7.2%。此外,有4对物种间呈无联结,占总种对数的1.9%。 图3 Spearman秩相关与Pearson相关检验半矩阵注:左下为Spearman秩相关,右上为Pearson相关。*为显著相关,**为极显著相关。Fig.3 Spearman rank correlation and Pearson correlation test semi matrix Pearson相关分析结果显示,正联结的有88对,负联结的有119对,分别占总对数的41.9%,56.7%,正负关联比为0.74。Pearson相关分析达到显著与极显著水平的有30对,占总对属的14.3%。其中,显著正联结的有8对,极显著正联结的有20对,分别占总对属的3.8%、9.5%;显著负联结的有2对,占总种对数的1.0%。此外,有3对物种间呈无联结,占总种对数的1.4%。 3.1.1 群落优势种的重要值和生态位特征 生态位宽度与物种的环境适应能力、资源竞争力密切相关。生态位宽度越大,物种对空间、养分等资源的利用能力越强[9]。本研究中,物种间生态位宽度的差异较大,帽斗栎、乌鸦果、香茶菜(Isodonamethystoides)等物种的生态位宽度较大,其抗逆性较强,能适应大幅度的环境变化,在植被恢复初期的群落建构中起着重要作用。其中,帽斗栎在植被恢复初期的群落中占据绝对优势地位,这可能与帽斗栎的生物学特性及林火的干扰有关。帽斗栎对环境的适应能力较强,在光照好的条件下可更好的生长发育,同时也是西南地区重要的乡土物种[25]。林火使乔木的林冠层受损严重,对林下植被的遮蔽大大减少。同时,树冠火对乔木种子库也造成了损害,进一步延长了乔木层恢复的时间,这给帽斗栎、乌鸦果等喜光物种的生长提供了有利的条件,导致火烧迹演替初期植物群落中的优势物种大多具有喜光的特点[26]。除此之外,火烧后该区域正处于一年中的雨季,降水为土壤水分提供了补给,在立地条件较好的区域物种表现出了喜湿的特点,如牛毛毡、双穗雀稗、针毛蕨等。这与调查时当地处于湿季存在一定的关联,帽斗栎等生长迅速的物种也对保持土壤水分起到了积极作用。一般来说,重要值大的物种生态位宽度也较大。中坝村火烧迹地植被恢复初期整体也呈现这样的趋势,但火烧迹地环境的异质性使两者之间的相关程度并不显著。植被恢复初期,受地形、海拔、坡度、坡向等自然因素的影响,资源在空间上的分布是不均匀的,各样方的小生境存在差异。针毛蕨、牛毛毡等喜湿物种,呈聚集分布,在湿度较大的样方中优势地位明显,提高了其在整体重要值中排名。但火烧迹地演替初期大多数区域坡度较大,缺少乔木的遮蔽,蒸发强烈,土壤含水量较低,使这些物种的频度较低。生态位宽度不仅与重要值有关,同时也受物种频度的影响,因此出现重要值与生态位宽度不一致的现象,这与刘润红等[4]、王彦阁等[27]的研究结果相一致。 3.1.2 群落优势种的生态位重叠 生态位重叠可以反映物种之间的生态相似性,具有相似生态需求的物种在资源不足的情况下会表现为竞争关系[28]。火烧迹地植被恢复初期,群落物种间的生态位重叠普遍不高,物种间竞争较缓和。一方面可能是林火短期内提高了土壤的养分[29],植被恢复初期物种对资源的利用还不够充分,部分资源还处于未被利用的状态;另一方面,部分物种之间由于竞争、生境异质性等因素产生了生态位分化[30-32],如香茶菜、针毛蕨、牛毛毡、这些物种生态习性相似,但物种间的生态位重叠较低,互补性较高。生态位宽度较大的物种对资源环境的利用能力更高,其分布更广,往往与其他物种形成较高的生态位重叠[27]。火烧迹地植被恢复初期物种间也呈现这样的特点,如生态位宽度较高的帽斗栎、乌鸦果、风箱树(Cephalanthustetrandrus)等与其他物种的平均生态位重叠均在0.4以上;生态位重叠为0的物种多为小叶三点金、羊耳菊、狸尾豆等生态位宽度较低的物种,这些物种在样方中的分布较少,不易与其他物种形成重叠。 3.1.3 群落优势种的种间联结 一般来说,次生演替初期群落物种间关系较分散,种对间的正负关联比较低,随着演替的进行,逐渐趋向于正联结,以求得多物种的共存[8]。中坝村火烧迹地植被恢复初期,群落整体呈不显著的正联结,与X2检验结果的正负关联比一致,而Spearman秩相关与Pearson相关的检验结果的正负关联比均<1,这说明群落还未形成稳定的群落结构,物种间也缺少稳定的搭配关系[33],群落处于不稳定阶段,演替到稳定群落还需要较长时间。本研究中,X2检验,Spearman秩相关,Pearson相关3种检验方法的结果基本一致,但也存在一定的差异。3种检验方法的结果均为呈显著正联结的种对比例大于显著负联结种对数,种对的显著率较低,说明植被恢复初期物种间竞争较弱,种间联结较为分散,这在一定程度上可以缓和物种间对资源和空间的竞争关系,有利于演替初期物种的共存。Spearman秩相关检验种对的显著率明显高于Pearson相关与X2检验。这是因为Pearson相关对数据的正态分布有要求,而野外采集的数据难以保证呈正态分布。X2是基于二元数据的检验结果,虽能够较好地反应物种间关联的显著程度,但在反映物种间联结大小方面存在不足,因此X2的检验结果需要结合Spearman秩相关及Pearson相关进行分析[34]。演替初期,群落物种间普遍存在相互关联[35],仅有极少数的物种间完全无关联,这可能是林火干扰后具有相似生境需求的物种在适宜区域聚集分布,使物种间普遍存在相互关联。火烧迹地植被恢复初期群落生态位重叠较大的物种种间关系多呈显著的正联结[7],如风毛菊与橘草、金丝梅与牛毛毡、帽斗栎与乌鸦果,这也说明环境资源较为充足,可以支持具有相似生态习性的物种对同一资源的分割而达到共存。 火烧迹地植被恢复初期,群落优势种种间整体呈不显著的正联结,有正向演替的趋势。群落优势物种的生态位宽度差异较大,帽斗栎是植被恢复初期群落的建群种。植被恢复初期物种之间普遍存在生态位重叠与种间联结,但重叠与联结程度均较低,种间关系分散,竞争较缓和。火烧迹地现有资源能够支持物种的生长发育,环境异质性现象明显,有利于后续的植被恢复与物种多样性的增加。当前火烧迹地乔木受损严重,群落结构和功能还不完善,距离稳定阶段还需较长时间的演替,在乔木受损严重的区域可以适当地进行人工植被修复。
2 结果与分析
2.1 优势物种的重要值与生态位宽度
2.2 优势物种间的生态位重叠

2.3 群落总体联结性


2.4 X2检验
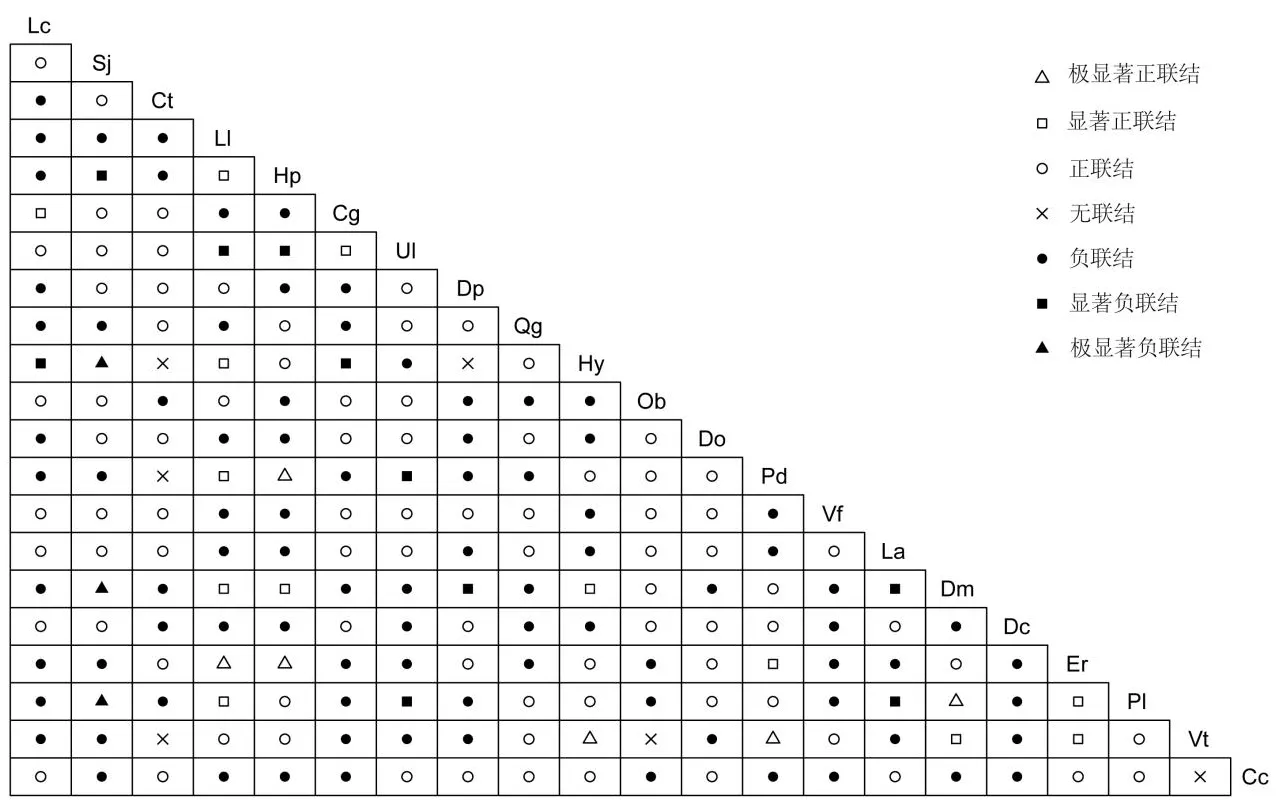
2.5 Spearman秩相关与Pearson相关分析

3 讨论与结论
3.1 讨论
3.2 结论
猜你喜欢
草业科学(2022年6期)2022-06-29中国农业科学(2022年10期)2022-06-28广东蚕业(2021年12期)2022-01-18中学生理科应试(2021年10期)2021-12-07语数外学习·高中版中旬(2021年12期)2021-03-09语数外学习·高中版上旬(2020年8期)2020-09-10南方周末(2019-06-27)2019-06-27小学生作文(低年级适用)(2018年12期)2018-03-23速读·中旬(2017年8期)2017-09-04新高考·高一数学(2016年10期)2017-07-06
