
亚美马褂木无性系幼林生长性状变异及选择研究*
2022-02-19 03:32:24杨军贺傅爱平钟永达吴照祥李辉虎刘腾云杨爱红陈彩慧徐立安余发新
西部林业科学 2022年1期
杨军贺 ,傅爱平 ,钟永达 ,吴照祥 ,李辉虎 ,刘腾云 ,杨爱红 ,陈彩慧 ,徐立安 ,余发新
(1.江西省科学院生物资源研究所/江西省观赏植物遗传改良重点实验室,江西 南昌 330096;2.南京林业大学南方现代林业协同创新中心,江苏 南京 210037;3.永丰李山林场,江西 吉安 331504)
目前,杂交育种依然是林木育种中最主要和最有效的育种手段之一,它能够有效结合双亲的优良性状,获得优于亲本的杂种优势,进一步选育出优良单株[1-2]。获得的优株通过无性繁殖,建立无性系测定林,最终选育出良种。通过无性繁殖建立测定林,既避免了杂种优势的丢失,还大大缩短了育种周期,降低了育种成本,提高了人工林的生产力[3-5]。为进一步缩短优良无性系选育时间,研究人员对林木的早期选择进行了大量研究[6-8]。鹅掌楸属(Liriodendron)为第三纪孑遗树种,现自然界中仅存分布于中国的马褂木〔Liriodendronchinense(Hemsl.) Sarg.〕和分布于北美洲的北美鹅掌楸(L.tulipiferaLinn.)[9]。南京林业大学已故著名林木育种学家叶培忠教授于1963年首次以中国马褂木为母本,北美鹅掌楸为父本,通过人工控制杂交授粉获得亚美马褂木(L.sino-americanumP.C.Yieh ex Shang et Z.R.Wang),又称杂交鹅掌楸[10]。其综合了双亲的优点,经多地点长时间栽培实践,表现出明显的杂交优势,速生性、适应性和抗逆性、观赏性优势明显,而且在木材密度、纤维长宽等各方面都达到了纸浆材及板材用材标准。因此,亚美马褂木在园林绿化和用材林营造中具有广阔的应用前景[11-14]。已有研究表明,亚美马褂木作为速生杂交树种,其树高、胸径等生长性状间在未达到成熟年龄之前相关性较高,树龄3 a时即具有较高的选择效率[15]。本研究旨在探索亚美马褂木无性系早期生长和遗传变异规律,以期为亚美马褂木无性系良种选育和生产实践提供技术支持。
1 材料与方法
1.1 试验地概况
试验地位于江西省乐平市鸬鹚乡(28°54′N,117°19′E),属亚热带潮湿季风气候,年平均气温18.3 ℃,活动积温5 700 ℃ 左右,常年无霜期219~313 d,年平均降水量1 672 mm,年蒸发量1 220 mm,年平均日照为1 766 h;海拔71 m,矮山丘陵地;坡向为南坡和西南坡,坡度介于25°~45°;红壤土,厚度及肥力中等。
1.2 试验材料及方法
2003年4月,以南京林业大学树木园内5株北美鹅掌楸的混合花粉为父本,南昌市林科所茶园山林场和罗亭乡洪家村各1株成龄马褂木为母本进行人工杂交授粉。2004年用杂交种子播种育苗后获得F1子代苗1 000余株。2013年从中选择72株优良单株种植于江西省科学院资源与环境综合实验基地并建立采穗圃(位于南昌县黄马乡)。2018年春季以1 a生扦插苗营造无性系试验林,受限于林地面积及成活率,试验林包括30个无性系。采用常规林木无性系育种的方式[16-18],试验为随机区组设计,10株小区,3次重复,株行距3 m × 4 m,四周设保护行。造林苗出圃时以及2018—2020年每年冬季在亚美马褂木进入休眠期后进行苗高和地径每木调查,共4次。地径采用游标卡尺(0.01 mm)测量,苗高采用钢卷尺(0.01 m)测量。
1.3 数据分析
运用Excel、SPSS 22.0等软件对测量数据进行统计分析。方差分析线性模型如下。
Yijk=μ+Bi+Fj+eijk
式中:Yijk为第i区组内第j个无性系的第k个单株的观测值,μ为总体平均值,Bi为第i个区组的固定效应,Fj为第j个无性系的随机效应,eijk为随机误差。
依据方差分析结果进行苗高和地径的遗传参数估算,包括重复力、变异系数、遗传和环境方差、遗传和现实增益等[19-21]。
采用性状表现水平分析法和加权系数法(权重赋值采用等权法,即以各性状的表型标准差的倒数作为该性状的权重)2种方法分别对试验林参试无性系进行评价排序,以20%入选率分别筛选出得分较高的无性系,将通过2种计算方法均能入选前20%的无性系选为优良无性系。具体计算公式[16,22]如下。
性状表现水平 =(无性系平均值 - 无性系中的最小值)/(标准差×4)
2 结果与分析
2.1 生长性状变异分析及多重比较
从表1中可以看出:亚美马褂木无性系出圃苗以及在造林1 a、2 a和3 a时的苗高平均值分别为0.44 m、0.60 m、1.22 m和2.27m,地径平均值分别为0.89 cm、1.49 cm、2.46 cm和4.49cm;苗高的表型变异系数从出圃苗到造林2 a苗逐年下降,造林3 a时趋于稳定,依次为74.53%、52.31%、23.27%和25.04%;而地径的表型变异系数则小于苗高,呈现出先下降后上升的变化趋势,依次为37.07%、14.49%、17.25%和19.68%。生长量统计结果表明亚美马褂木无性系的生长性状在幼林时期较不稳定。
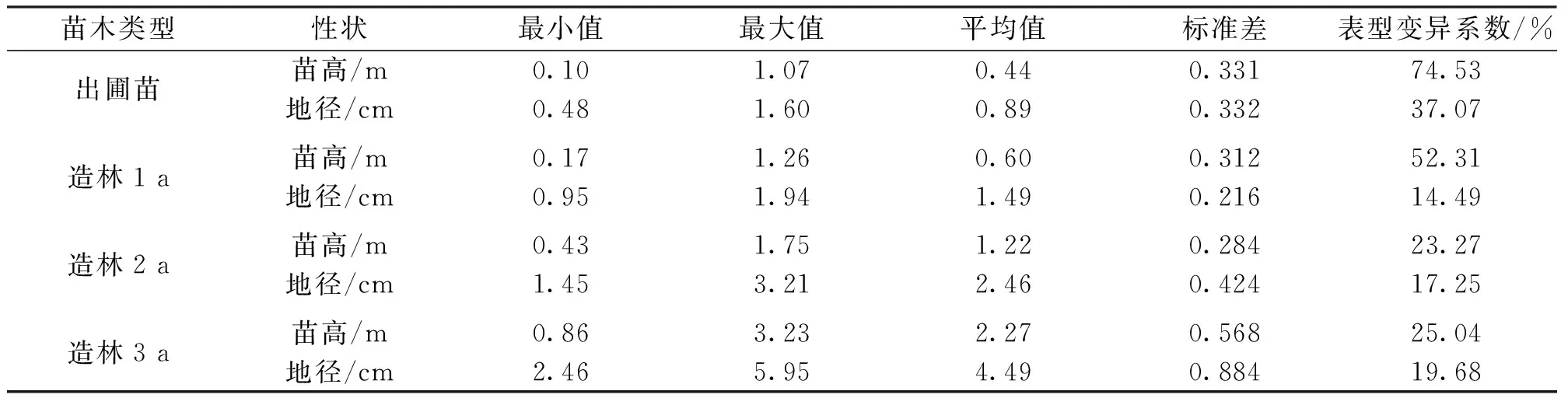
表1 亚美马褂木无性系的生长性状统计Tab.1 Statistics of growth traits of L.sino-americanum clones
对亚美马褂木造林3年时各无性系的生长性状进一步进行统计和多重比较(表2),结果发现:造林3 a时苗高高于总体平均值20%的无性系有7个,分别为无性系祥13、2、8、23、24、29和2-3-0,现实增益分别为26.03%、38.38%、23.89%、37.36%、25.86%、41.57%和35.20%;地径高于总体平均值20%的无性系有6个,分别为无性系祥13、2、8、23、24和29,现实增益分别为21.22%、31.87%、22.51%、31.18%、28.37%和30.08%;无性系祥13、2、8、23、24和29的苗高及地径均高于总体平均值20%;生长量最低的无性系为1-3-0,其苗高和地径仅为总平均值的37.79%和54.46%。
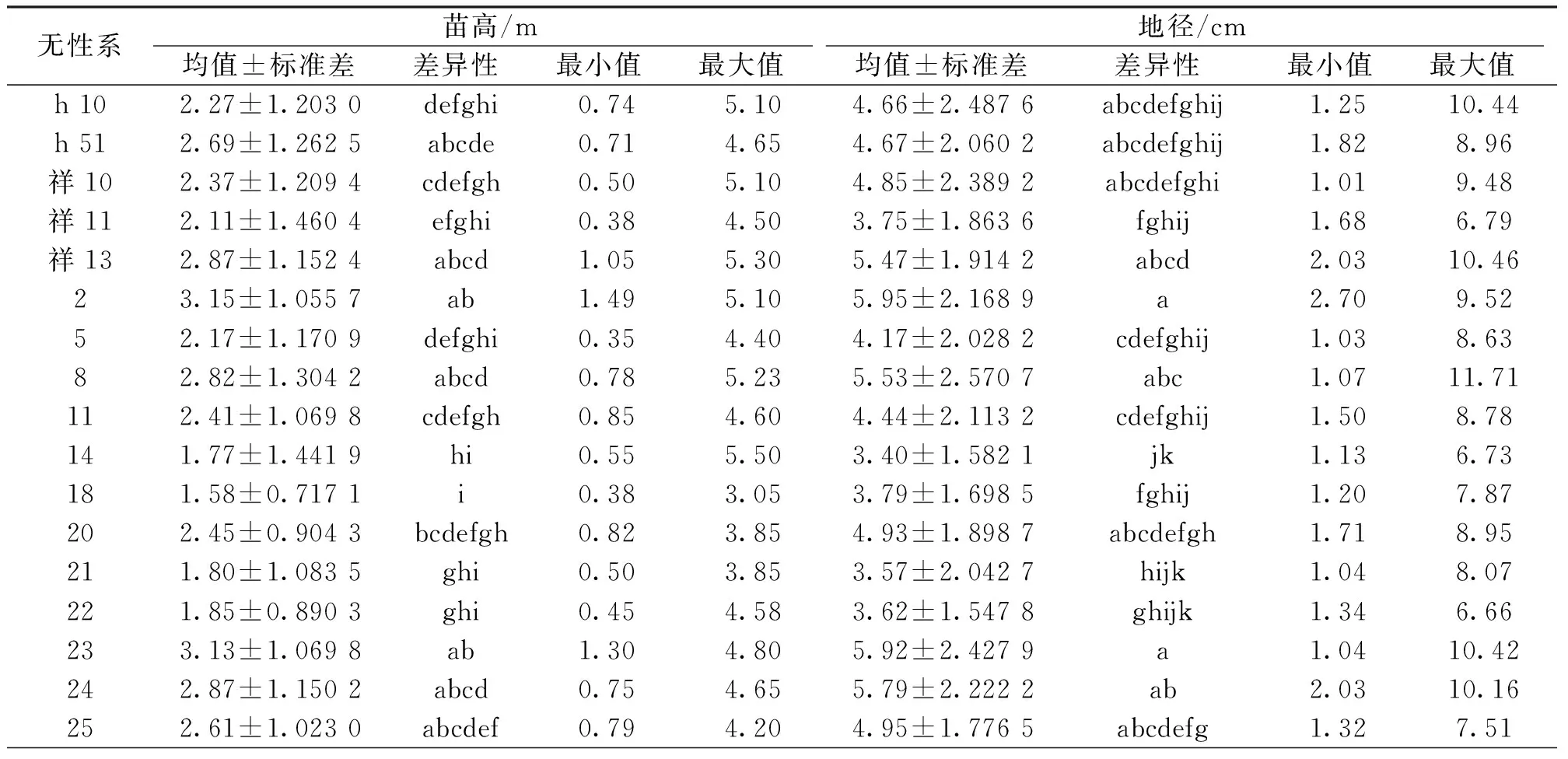
表2 3年生亚美马褂木无性系生长性状统计及多重比较Tab.2 Growth status and multiple comparison of 3-years-old L.sino-americanum clones
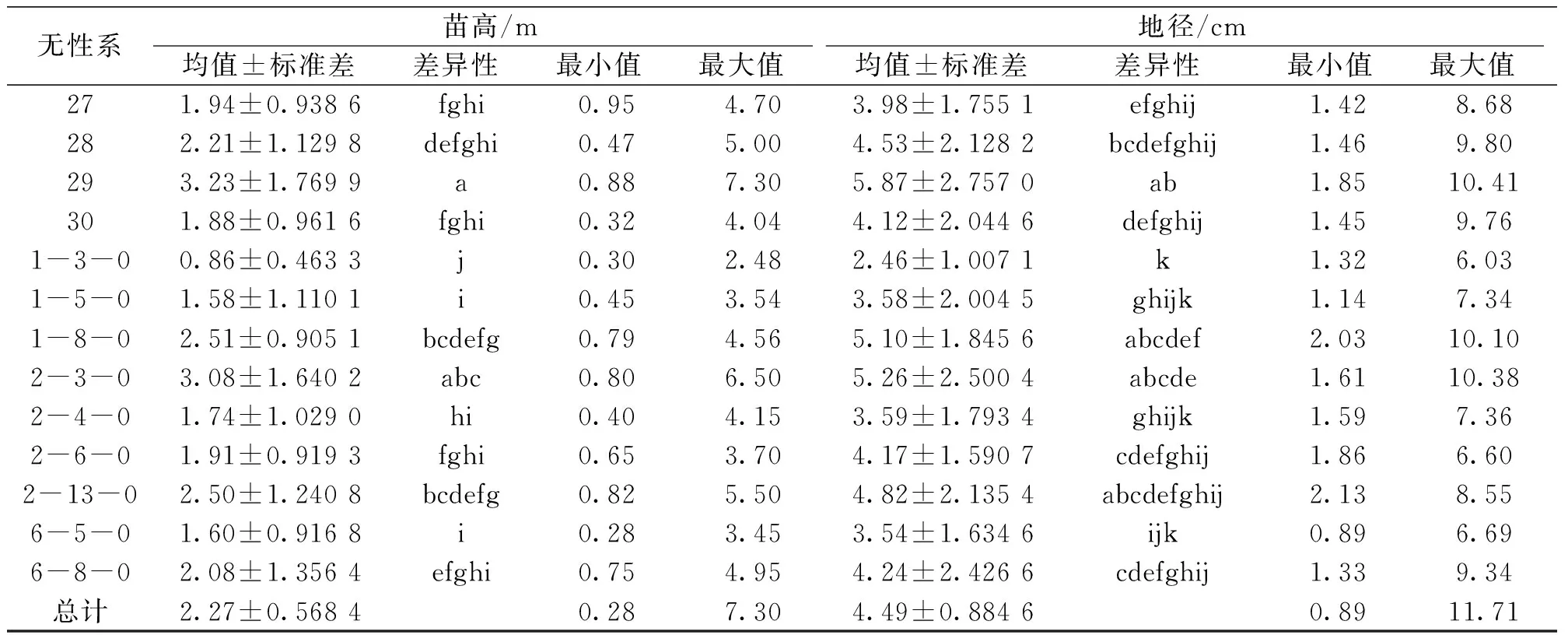
续表2
2.2 方差分析及遗传参数估算
对亚美马褂木无性系试验林出圃苗以及造林后的生长性状分别进行方差分析,从表3可以看出,除在造林2 a时地径性状在区组间存在显著差异(P<0.05)及造林3 a时地径性状在区组间差异不显著(P>0.05)外,其它生长性状在无性系间和区组间均存在极显著差异(P<0.01)。表明从这些优良单株无性系中进行良种选育是可行的。
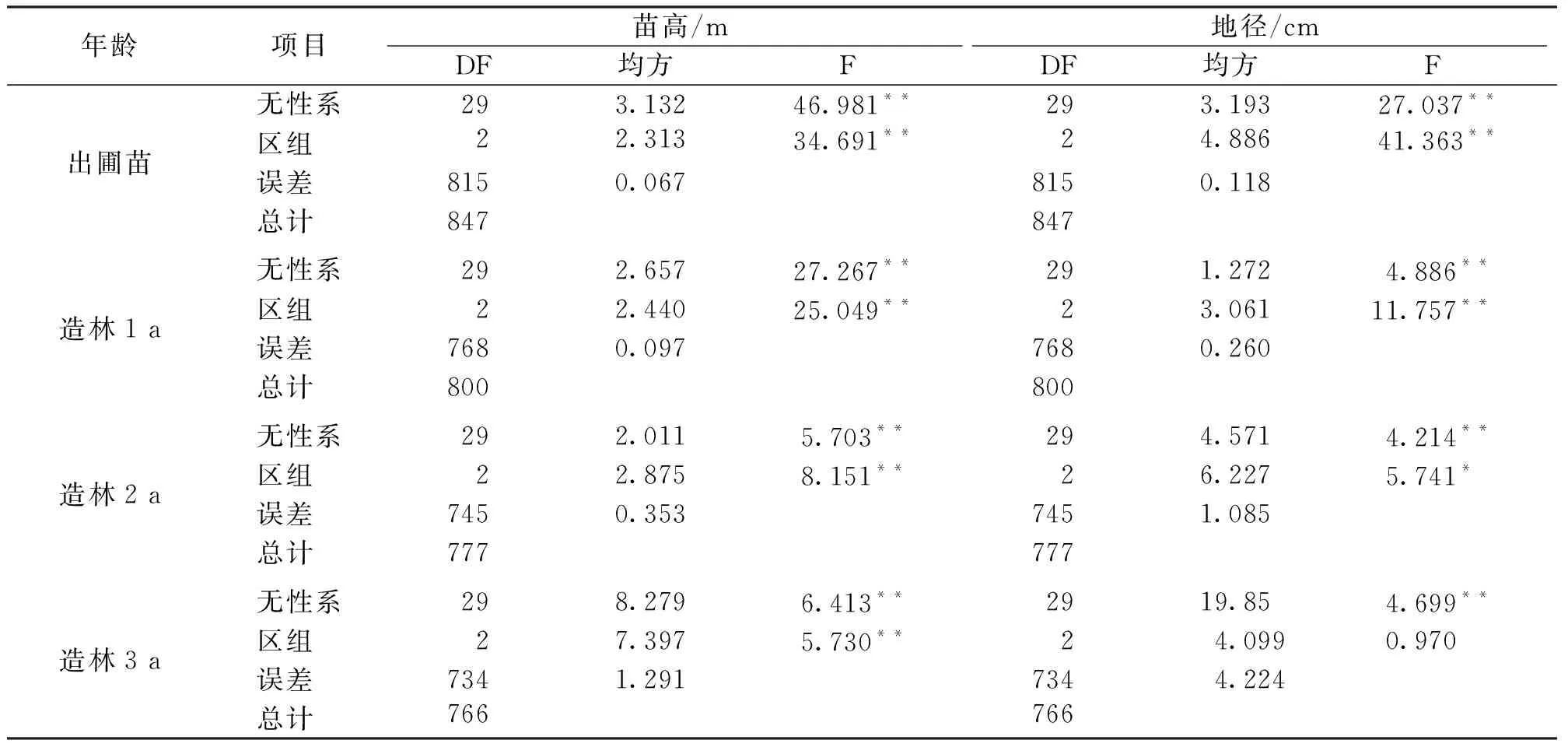
表3 亚美马褂木无性系生长性状方差分析表Tab.3 Variance analysis of growth traits of L.sino-americanum clones
亚美马褂木无性系试验林各年份生长性状的遗传变异系数(表4)均远高于表型变异系数(表1),且随着林龄的增加,遗传变异系数和表型变异系数的下降幅度大体一致,表明其产生变异的主要原因仍然是无性系间基因型的差异。出圃苗苗高和地径的遗传方差分量分别为1.022和1.025,而环境方差分量分别为0.067和0.118,并且苗高和地径的单株重复力和无性系重复力均在0.9以上,进一步说明亚美马褂木无性系在苗圃地内较理想环境下产生的丰富变异主要是由基因型差异所造成的[23]。在全部观测年份内,苗高和地径的无性系重复力均大于单株重复力,造林3 a时苗高和地径的无性系重复力依然可以达到0.84和0.79,表明无性系比单株具有更加明显的遗传效应。因此,在此试验林的无性系间开展选优工作具有更大可靠性。

表4 亚美马褂木无性系生长性状主要遗传参数Tab.4 Main genetic parameters of growth characters of L.sino-americanum clones
2.3 优良无性系初步选择及遗传增益
采用性状表现水平分析法和加权系数法分别对造林3年时无性系试验林进行综合评价和排序,结果见表5。

表5 亚美马褂木无性系综合评价及排序Tab.5 Comprehensive evaluation and sorting of L.sino-americanum clones
其中,加权系数法相关性状经济权重值的确定采用等权法,即以各性状的表型标准差的倒数作为该性状的权重[19],通过计算得出苗高和地径的权重值分别为0.6和0.4。采用性状表现水平法进行综合评价排序的结果中,无性系水平值超过无性系群体平均水平值的无性系有15个,分别超过群体平均水平值4.99%~41.28%,无性系水平值达到100以上的无性系有祥23、29、8和2-3-0,水平值分别超过群体平均水平值31.61%~41.28%,而无性系水平值最低的无性系为1-3-0,其无性系水平值仅有群体平均水平值的36.78%,为最高水平值的26.04%。采用加权系数法进行综合评价排序的结果中,I值超过无性系群体平均值0.80的无性系有15个,分别超过群体平均值1.56%~37.79%,而I值大于1的无性系有29、2、23、2-3-0、24、祥13和8,分别超过群体平均值23.92%~37.79%,I值最低的无性系同样是1-3-0,其I值仅有群体平均值的44.38%,为I值最大无性系的32.21%。
以20%的入选率,根据性状表现水平分析法选出的优良无性系为23、29、8、2-3-0、24和祥10,根据加权系数法选出的优良无性系为29、2、23、2-3-0、24和祥13。以此结果来看,虽然通过两种方法所得到的排序结果有一定差异,但入选的优良无性系差别不大,分别入选的6个优良无性系有4个是共有无性系,分别为23、29、2-3-0和24。共有优良无性系苗高和地径的平均值分别为3.08 m和5.71 cm,平均遗传增益分别为30.05%和21.38%,苗高和地径遗传增益最高的无性系分别是29和23,为35.62%和25.04%,苗高和地径遗传增益最低的无性系分别是24和2-3-0,为22.30%和13.48%;苗高和地径平均现实增益为35.60%和27.16%,同样,苗高和地径现实增益最高的无性系分别是29和23,为42.19%和31.82%,最低的无性系分别是24和2-3-0,为26.42%和17.13%(表6)。
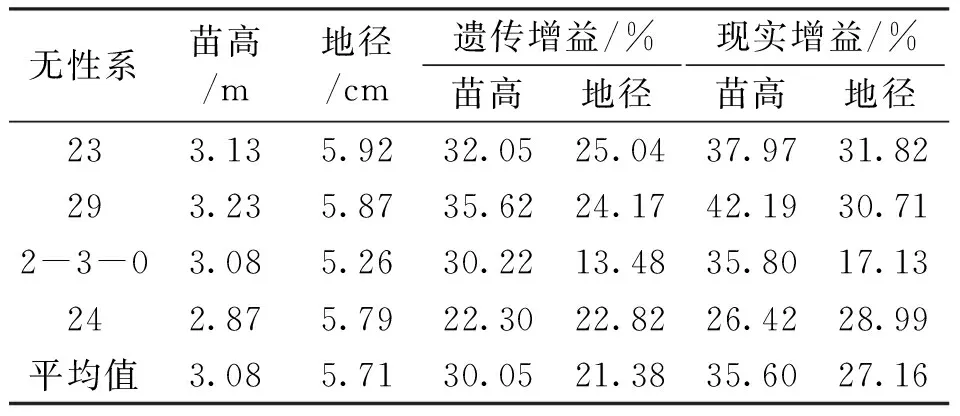
表6 入选亚美马褂木无性系生长性状值及遗传增益Tab.6 Growth characters and genetic gain of clones selected in L.sino-americanum
3 讨论与结论
为进一步研究亚美马褂木杂交优势,本研究以优良亲本杂交授粉获得F1半同胞子代实生苗,选择30个优良单株进行无性系扩繁,营建无性系子代测定林。通过对30个无性系生长性状的差异分析以及遗传参数估算,结果发现亚美马褂木苗高和地径在无性系间存在极显著差异(P<0.01),无性系较单株的遗传效应更加明显。袁晓峰[24]通过对两批亚美马褂木优良单株无性系苗期生长量进行分析,发现地径、苗高差异在无性系间均达极显著水平,表明开展亚美马褂木无性系选择是可行的,以苗高平均值高于参试无性系苗高平均值为指标,结合成活率,共复选出92个无性系。舒伟林[15]根据亚美马褂木无性系胸径、苗高、材积及重复力4个指标,通过综合指数选出了8个速生无性系,其单株胸径、苗高、材积年均生长量遗传增益分别达14.09%、11.97%及47.14%。李婷等[25]、徐瑢等[26]分别对白花泡桐(Paulowniafortunei)和白蜡树(Fraxinuschinensis)无性系进行了早期评价与选择,其研究结果均选择出了遗传增益较高的优良无性系,说明了早期选择的科学性。
亚美马褂木无性系从出圃苗到造林3 a时,苗高的遗传变异系数和表型变异系数均随着林龄的增大而降低,地径则呈现出先降后升的趋势,但升高不明显,而苗高和地径的无性系重复力以及单株重复力均随着林龄的增大而降低。张春回等[27]在研究亚美马褂木早期生长表现中也发现类似规律。对其它树种如尾叶桉(Eucalyptusurophylla)[28]的研究也有类似的结果,这表明环境对无性系试验林的影响不可忽略。王云鹏等[29]对木荷(Schimasuperba)的生长性状及材性性状的研究发现,其生长性状除遗传控制外,还受遗传与环境互作的影响,从而应开展环境与基因型互作研究,选择多个地点营建无性系试验林,以提高推广生产时的适应性[25,29-31]。本研究采用性状表现水平分析法和加权系数法分别对参试无性系进行综合评价排序[16,19,22],从评价结果来看,与王胤等[22]采用同样的两种方法对马尾松(Pinusmassoniana)无性系的综合评价选择结果不同,加权系数法各性状经济权重的设置和参试无性系数量不同可能是造成评价结果不同的原因。
虽然早期选择可大幅度缩短育种周期,但同时也因林木的生长周期较长而错过后期生长潜力大的无性系。因此,本研究初选出较高比例的无性系,通过连年观测试验林无性系生长性状,多阶段结合进行优良无性系选育[32-34]。后续研究宜增加材性性状作为选优指标,进行多性状综合选择以满足更多培育目标的要求并最大限度地提高选择增益。
综上所述,本研究对30个亚美马褂木无性系的生长性状进行差异分析以及遗传参数估算,结果表明苗高、地径的遗传效应表现为无性系较单株的明显,在无性系间进行良种选择是可行的;采用性状表现水平分析法和加权系数法综合初选出较高比例的无性系并从中优选4个优良无性系,分别为无性系23、29、2-3-0和24,其平均苗高和地径分别为3.08 m和5.71 cm,平均遗传增益和现实增益分别为30.05%、21.38%和35.60%、27.16%。研究结果可为亚美马褂木良种推广应用以及后续研究提供材料和技术参考。
猜你喜欢
法人(2023年9期)2023-12-01 14:50:22
科教新报(2022年13期)2022-05-23 05:16:50
武汉工程职业技术学院学报(2022年1期)2022-04-13 06:31:30
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
现代农业科技(2017年24期)2018-01-22 21:35:06
中学数学杂志(高中版)(2014年2期)2014-05-26 13:42:42
电测与仪表(2014年6期)2014-04-04 11:59:46
水动力学研究与进展 B辑(2012年4期)2012-08-22 08:31:49
