
植物分枝性研究进展及其在蔬菜育种中的应用
2022-02-13 12:57夏雨桐
中国蔬菜 2022年1期
夏雨桐 王 琛 郝 宁 武 涛*
(1 湖南农业大学园艺学院,园艺作物种质创新与新品种选育教育部工程研究中心,蔬菜生物学湖南省重点实验室,湖南长沙 410128;2 东北农业大学园艺园林学院,黑龙江哈尔滨 150030)
分枝是植物株型形态建成的重要农艺性状之一。分枝可影响作物产量,同时分枝状况也会影响作物的栽培密度和营养分配。较多的分枝会导致通风不良,极易使作物发生病害造成减产。此外,分枝也是观赏植物一个重要的株型性状,分枝的生长状态可影响观赏植物的造景效果。近些年,分枝逐渐成为育种家们较为关注的作物地上部性状之一。
植物分枝一般分为二叉分枝、假二叉分枝、合轴分枝、单轴分枝和分蘖等。单轴分枝的主要代表植物有拟南芥和黄瓜等;合轴分枝的主要代表植物是番茄;分蘖的主要代表植物为大多数禾本科植物,例如小麦和水稻;假二叉分枝的主要代表作物有茄子;而辣椒一般属于二叉分枝或者三叉分枝(巩鹏涛和李迪,2005)。
1 植物分枝的环境调控
植物的分枝受光照、温度、营养状态等环境因素的影响(Snowden &Napoli,2003),其中光照对植物分枝具有重要影响。研究表明,高强光可通过促进越桔侧芽生长从而促进分枝(Kawamura &Takeda,2002)。低光量子通量密度会减少拟南芥分枝(Su et al.,2011)。红光与远红光的比率降低时,分枝抑制因子()表达升高从而导致拟南芥分枝减少(Ballare,1999;Franklin &Whitelam,2005;Gonzalez-Grandio et al.,2013)。光合面积减少可促进分枝抑制基因的表达进而减少玉米的分蘖(Kebrom &Mullet,2015)。超过一定范围的光密度,会导致番茄中抑制分枝的激素独脚金内酯(strigolactones,SLs)含量增加和SLs 合成基因()的表达升高,从而抑制番茄分枝(Koltai et al.,2011)。此外,亮/暗(7 h/7 h)过渡频率的增加会抑制拟南芥腋芽的发育从而减少分枝(Jouve et al.,1998a,1998b)。温度也是影响分枝的重要因素之一,低温会诱导月季中的SLs 信号基因()的表达,促使其在茎节形成梯度进而影响侧枝的萌发率(Djennane et al.,2014)。低温也会促进SLs 受体基因()的转录从而抑制水稻的分蘖(Chen et al.,2018)。营养状态也可调控植物的分枝发育。研究表明,糖类的输入和输出对桃树腋芽的萌发有重要作用(Karine et al.,2004)。豌豆茎尖通过限制转移到腋芽的糖量来限制腋芽的生长(Michael et al.,2014)。缺糖导致了小麦突变体的少分蘖表型(Kebrom et al.,2012)。异麦芽酮糖可以促进芽的萌发从而促进分枝,蔗糖、果糖、帕拉金糖和果胶都能促进离体培养的玫瑰侧芽在光照下的萌发(Rabot et al.,2012)。磷和氮通过影响植物激素的合成及运输来调控拟南芥侧枝的生长发育(Evers et al.,2011)。缺磷通过抑制菊花腋芽的伸长来减少分枝长度,使其株型紧凑(郗琳,2015)。研究表明,氮肥对水稻的分蘖有促进作用(Li et al.,2016)。低氮处理可降低拟南芥中多分枝双突变体-/-的分枝数量(Madeleine et al.,2017)。
2 植物分枝的激素调控
植物激素是打破侧芽休眠和分枝持续生长必不可少的因素。外界环境的改变可使植物激素发生相应的变化,促使植物体内激素达到动态平衡,进而激发调控植物分枝基因的表达。
2.1 生长素
最早发现作用于植物分枝的激素是生长素(auxin,IAA)。提高植物体内IAA 水平后,侧枝数目会明显降低(Hall &Hillman,1975;Morris,1977;Greg et al.,1991;Romano et al.,1993)。研究表明,黄瓜茎尖中IAA 不能独立抑制腋芽的生长,需要与其他抑制腋芽生长的物质共同作用(徐庆华 等,2011)。对去除颖花的水稻喷施IAA后,水稻分蘖伸长受到抑制(Arite et al.,2007)。研究发现,IAA 含量处于最低值是腋生分生组织(AM)起始的关键(Gallavotti,2013)。IAA 合成途径(丙酮酸途径)中2 个重要基因()和()的突变体合成IAA 的能力不强,导致顶端优势受到抑制进而促进拟南芥分枝(Cheng et al.,2006)。玉米的同源基因功能缺失会导致AM 起始和侧部器官发育发生缺陷,从而减少分枝(Phillips et al.,2011)。黄瓜在驯化过程中侧枝抑制基因的表达量逐渐增高,在侧枝中直接抑制IAA 输出基因的活性,导致侧枝中IAA 过量积累,从而抑制黄瓜侧枝的生长发育(Shen et al.,2019)。IAA 信号转导关键基因、、的突变或缺失通过影响IAA 的信号转导进而促进番茄侧枝生长(Chaabouni et al.,2009;Deng et al.,2012)。
2.2 细胞分裂素
细胞分裂素(cytokinin,CTK)可通过促进细胞分裂来活化侧芽,促进植物分枝。直接对植物侧芽喷施CTK 可抑制顶端优势使侧芽生长。研究发现,拟南芥、等多分枝突变体中的CTK含量较高,说明CTK 正调控植物分枝(Catterou et al.,2002;Zubko et al.,2002)。对拟南芥基部施加CTK 可减弱顶端优势,促进侧芽生长(Chatfield et al.,2000)。CTK 还可以促进AM 的起始和侧芽的形成,拟南芥()多分枝突变体中由于CTK 水平增加导致莲座叶和茎生叶的叶腋形成多个AM,增加分枝的数量,同时也可以促进侧枝的伸长(Tantikanjana et al.,2001)。CTK 合成酶关键基因()高表达可促进CTK 对侧芽的活化作用,进而促进拟南芥的分枝(Guo &Gan,2011)。
2.3 独脚金内酯
SLs 的出现为研究植物腋芽的生长发育机制提供了一种新的思路(Rameau,2010)。有研究表明,SLs 可以作为一种长距离信号物质抑制侧芽的生长发育,并且作为一种嫁接转移物质调控TCP 家族的分枝抑制转录因子BRC1、TEOSINTE BRANCHED 1(TB1)、FINE CULM 1(FC1)的表达,从而抑制侧芽伸长(Antonio et al.,2007;Shinohara et al.,2017)。SLs 可通过下调豌豆腋芽内的CTK 水平来抑制侧芽生长(Victoria et al.,2008)。研究发现,SLs 可抑制打顶豌豆的侧枝伸长从而抑制植物分枝(Dun et al.,2009)。在拟南芥中SLs 可通过减少IAA 输出载体蛋白PIN 的数量来降低IAA 极性运输的能力,从而抑制侧枝生长(Ongaro &Leyser,2007)。随着对SLs 的研究越来越深入,从生物合成到激素运输再到信号转导,SLs 调控植物分枝的机理也越来越清晰。SLs受体复合物D14-MAX2 的底物D53-like-SMXLs和BES1 相互作用,抑制的表达,从而促进拟南芥分枝(Hu et al.,2020)。SLs 的信号通路中有3 个抑制因子,分别是、、,研究发现在没有SLs 的情况下SMXL6 +TPL 蛋白可直接与、、的启动子结合并抑制其表达来增加SLs 的含量,从而抑制拟南芥分枝。同时,SMXL6 还可以与未知的转录因子形成复合物,这些转录因子有望识别并结合、或的启动子,从而抑制它们的转录来促进拟南芥分枝(Wang et al.,2020)。
2.4 其他激素
研究发现,赤霉素(gibberellin,GA)、脱落酸(abscisic acid,ABA)和油菜素内酯(brassinosteroids,BRs)都可以调节植物分枝。水稻()基因可抑制分蘖,高水平的GA 可以激活APC/C(E3 泛素连接酶复合物),促进AM 的MONOCULM 1(MOC1)降解,从而抑制水稻分蘖(Lin et al.,2020)。高浓度ABA 会抑制拟南芥侧芽的萌发从而抑制分枝(Pei et al.,1998)。BRs 信号通路调控基因会抑制拟南芥AM 起始关键基因-()的表达,从而抑制AM 启始,抑制分枝(Gendron et al.,2012)。为获得影响植物分枝的激素类型,对相关研究文献进行了总结(表1)。
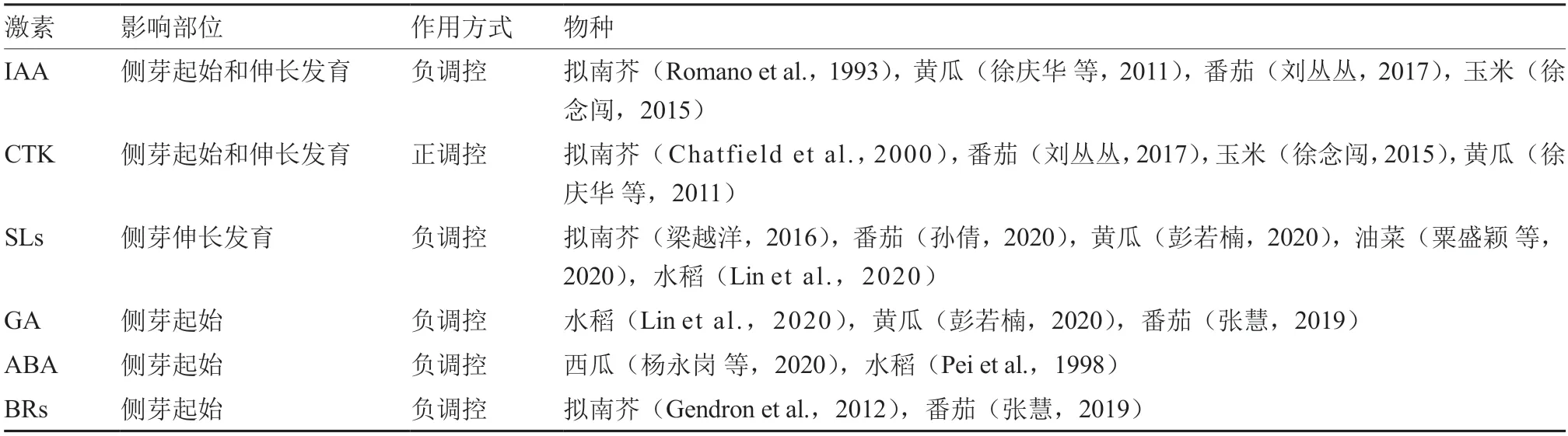
表1 影响植物分枝的激素类型
3 植物分枝的基因调控
影响植物分枝除了环境因素和植物激素外,还有遗传因子。植物的分枝主要是由AM 起始,发育成腋芽,再由腋芽发育成侧枝。所以其内在调控基因分为3 种:一是侧芽起始发育调控基因;二是侧芽伸长发育调控基因;三是侧芽起始和伸长发育协同调控基因。
3.1 侧芽起始发育调控基因
植物侧芽的起始主要受到基因的调控。()属于WRKY 转录因子家族,可促进AM 的启动,产生较多的侧芽来促进拟南芥分枝(于延冲,2011;Guo et al.,2015)。-(-)属于MYB 转录因子家族,正调控拟南芥分枝(Muller et al.,2006)。在的上游,正调控的转录促进AM 起始,从而促进拟南芥分枝(于延冲,2011;Guo et al.,2015)。番茄中基因是的同源基因,参与AM 起始(Schmitz et al.,2002)。NAC 转录因子CUC 参与拟南芥AM 启动(Stirnberg et al.,1999;Raman et al.,2008;宋玉光 等,2016),在拟南芥AM 形成中发挥重要作用(Hibara et al.,2006)。研究发现,CUC2 和CUC3 直接与DA1(其肽酶底物UBP15 可抑制AM 起始)启动子结合并激活其表达,形成CUC2/CUC3-DA1-UBP15 调控模块来控制拟南芥AM 的启动(Li et al.,2020)。()属于GRAS 转录因子家族,促进AM 形成,正调控拟南芥分枝(Greb et al.,2003)。和分别是在水稻和番茄中的同源基因,正调控AM 起始(Groot et al.,1994;Li et al.,2003)。研究表明,在AM 的启动和建立过程中,和在拟南芥()的上游起作用(Matsuoka et al.,1993;Long et al.,1996;Li et al.,2003;Liang et al.,2014)。()属于HDZIP 转录因子家族,作用于拟南芥AM 起始(Talbert et al.,1995)。可上调基因表达来启动拟南芥AM(Otsuga et al.,2001)。属于KNOX转录因子家族,通过调控AM 细胞的分化来促进拟南芥AM 形成(Williams &Fletcher,2005;Scofield &Murray,2006)。()基因通过维持干细胞的活性和调控下游基因促进腋芽原基形成;-(-)基因主要功能是促进拟南芥AM 细胞分化,STM 蛋白结合在启动子上,上调其表达,同时受到的调控从而影响拟南芥腋芽原基形成(李亚栋 等,2009;周 超,2015)。()和()均 属 于bHLH 转 录 因子家族,调控AM 形成,在水稻和突变体中都会产生较少的AM 和分蘖(张彩霞,2014)。玉米中的()基因和拟南芥中()基因为的同源基因,通过编码蛋白调节分蘖或分枝(Gallavotti et al.,2004;Yang et al.,2012)。为获得影响侧芽起始发育调控基因的类型,对相关研究文献进行了总结(表2)。
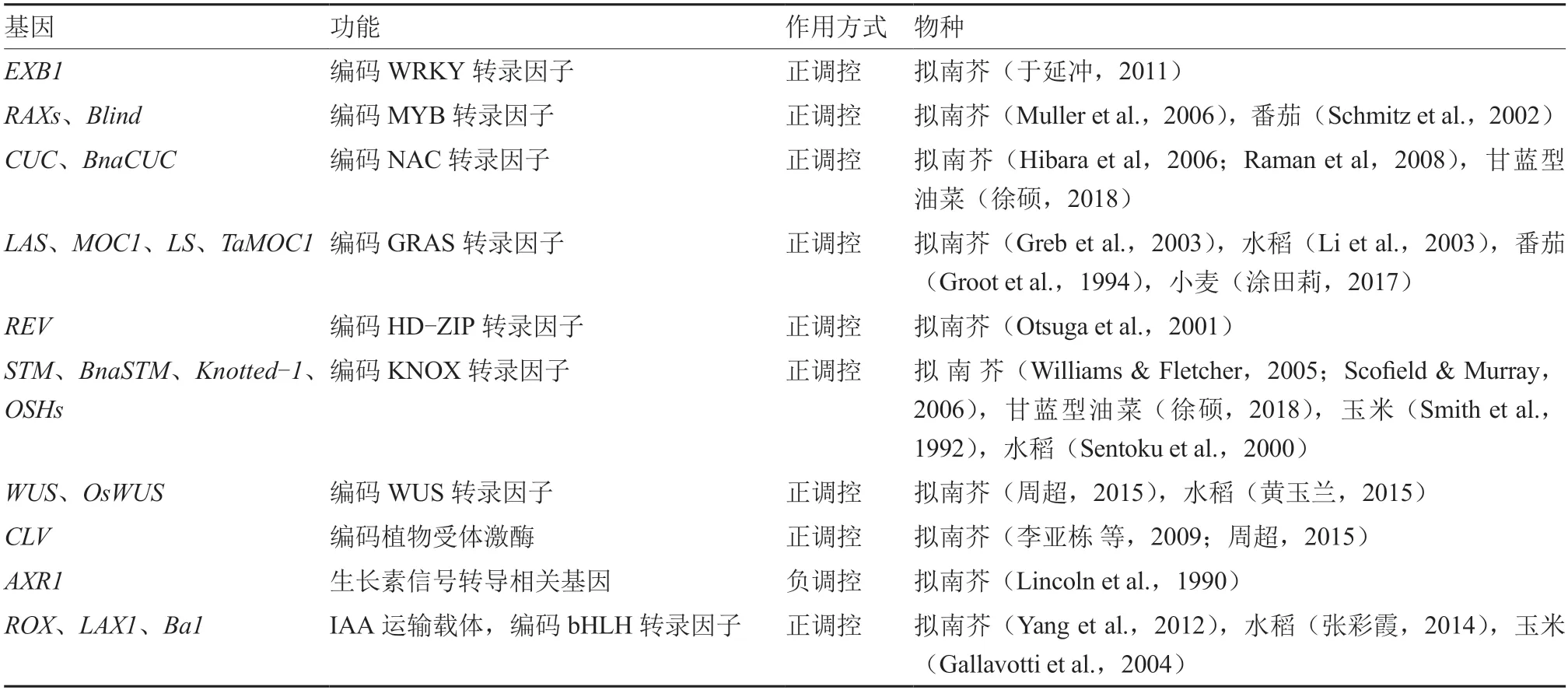
表2 侧芽起始发育调控基因
3.2 侧芽伸长发育调控基因
、、分别是拟南芥、玉米、水稻中的同源基因,都是TCP 转录因子家族的成员,抑制分枝、分蘖的生长。通过QTL 定位发现,玉米的基因造成了玉米和多分蘖Teosinte 玉米之间的结构差异(Wang et al.,1999;Hubbard et al.,2002)。主要在AM 中表达,以协调玉米芽的 生 长(Finlayson,2007)。在AM 和SAM基部均有表达,通过促进芽休眠来抑制水稻分蘖(Takeda et al.,2003)。拟南芥的基因在腋芽中表达,抑制腋芽伸长发育,是多个调控途径的下游基因(Antonio et al.,2007)。黄瓜分枝抑制基因可以和生长素极性运输基因协同调控黄瓜分枝(Shen et al.,2019)。基因属于细胞色素P450 家族,通过减少IAA 含量,抑制顶端优势促进拟南芥分枝(Bak et al.,2001)。在拟南芥中,基因是IAA 反应基因,可抑制腋芽的生长(Lincoln et al.,1990;Stirnberg et al.,1999)。()基因负调控IAA 信号转导,可以促进拟南芥分枝(Michniewicz et al.,2007)。基因负调控CTK 含量,从而抑制拟南芥分枝(Guo &Gan,2011)。基因参与多胺的形成,突变体会导致拟南芥分枝增加(Cui et al.,2010)。基因参与水稻中SLs合成来抑制分枝(Lin et al.,2009)。基因编码细胞色素P450,作用于、的下游,是SLs 信号转导的关键基因,抑制拟南芥分枝(Booker et al.,2005)。水稻中基因与拟南芥中基因同源,是SLs 调控基因中的重要基因,在、、下游起作用,可抑制分 枝(Zou et al.,2005,2006;Hu et al.,2020)。水稻中基因与拟南芥中基因同源,与为等位基因,是编码SLs 合成酶的关键基因,抑制分枝(Zou et al.,2005,2006);水稻中基因与基因同源,参与SLs 的生物合成,抑制分枝(Zou et al.,2006;Wang &Li,2011)。水稻中3 个等位基因、、在SLs 信号转导中发挥作用,是水稻分蘖的负调控因子(Arite et al.,2007,2009;Brady et al.2007;Zhou et al.,2013)。而是SLs 信号转导中的抑制子,正调控水稻分枝(Jiang et al.,2014)。玉米可负调控分蘖抑制基因和/-的表达,并与转录辅阻遏蛋白TOPLESS 互作,从而促进分蘖(Studer et al.,2011;Zhang et al.,2019)。为获得影响侧芽伸长发育调控基因的类型,对相关研究文献进行了总结(表3)。
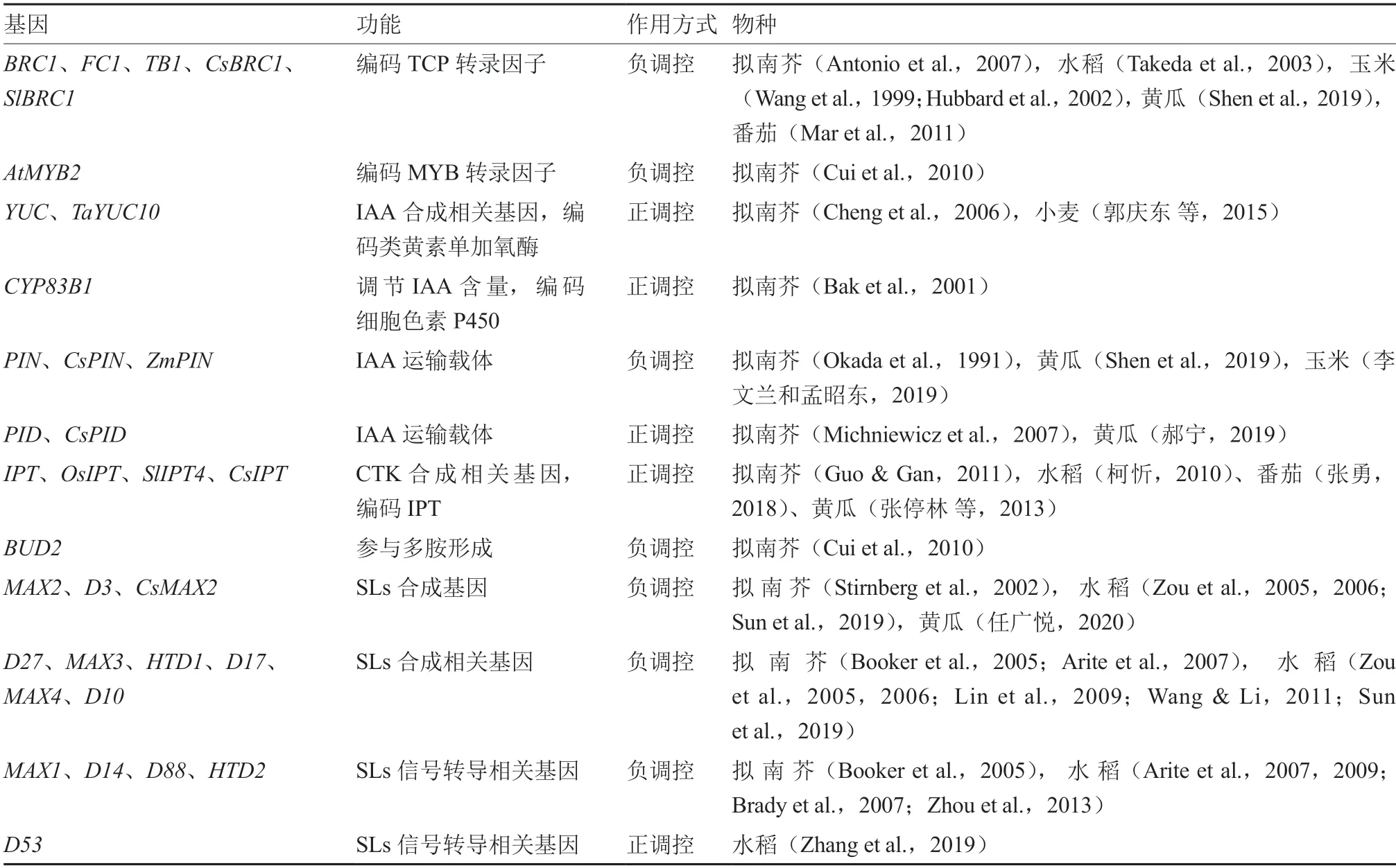
表3 侧芽伸长发育调控基因
3.3 侧芽起始和伸长发育协同调控基因
在AM 起始和侧芽发育都起作用的基因是和(),它们是同源基因,基因主要在叶腋处强烈表达,控制IAA 的合成来调控拟南芥分枝(Reintanz et al.,2001)。拟南芥突变体特征是在叶腋中形成的分生组织数量增加以及侧芽的停滞释放,导致侧芽大量繁殖,从而形成了可重复的分枝(Tantikanjana et al.,2001)。也有研究表明,基因通过加速CTK 的氧化降解来降低叶腋处的CTK 的浓度,使得AM 的起始和腋芽的发育都受到抑制,导致拟南芥分枝减少(Grbic &Bleecker,2000;Tantikanjana et al.,2001)。
4 植物分枝性在蔬菜育种中的应用及展望
我国蔬菜产值约2.2 万亿元,已经超过粮食作物成为种植业第一大产业。番茄和黄瓜等茄果类和瓜类蔬菜是我国设施栽培的主要蔬菜作物,且以搭架栽培为主。由于分枝较多,在茄果类和瓜类蔬菜生产中需要频繁地进行整枝、打杈、吊蔓等耗费人工的操作,增加了人工成本;其次,分枝较多还容易造成植株养分分配不均,易发生病害,从而降低了蔬菜作物的产量和品质。随着人工成本的逐渐增加,培育具备理想株型(侧枝少或弱)、适宜轻简化栽培的茄果类和瓜类蔬菜品种是我国蔬菜产业亟待解决的问题之一。在生产过程中,除了选择理想株型品种之外,也可根据上述研究结果对茄果类和瓜类蔬菜的分枝性进行调控。首先,可通过改变生长环境条件来调节植株分枝。例如,可在栽培环境中适当增加远红光、降低温度或增施氮肥来减少分枝,也可通过减少钾肥和糖类物质来减少分枝。第二,可利用植物激素调节分枝。通过喷施IAA、SLs、GA、ABA、BRs 等减少分枝,同时CTK 抑制剂处理亦可减少分枝。第三,利用已鉴定的植物分枝调控基因开发分子标记,通过分子标记辅助育种培育理想株型新品种。闫立英等(2015)通过分子标记辅助选择技术等育种手段选育出了适合棚室省力化栽培且分枝性强的优质丰产旱黄瓜品种“绿岛7 号”。第四,利用已克隆的植物分枝调控基因,可以通过基因编辑技术获得不同分枝类型新种质。Heidi(2017)对番茄基因进行编辑,获得了有分枝且花果大小合适的番茄种质材料。在黄瓜方面,中国农业大学张小兰教授团队克隆了调控黄瓜分枝发育的关键基因,由于中国的黄瓜种植多倾向于选择侧枝数目少、长度短、萌发时间晚的品种,因此可以利用基因信息,开发分子标记,或者通过基因编辑技术鉴定、创制不同分枝类型黄瓜种质资源(Shen et al.,2019)。随着人们对于植物分枝调控理解的不断深入,将会有效解决茄果类和瓜类蔬菜生产中分枝较多的问题,创新种质资源,并培育出符合育种目标的优质、高产蔬菜新品种,有效解决蔬菜生产过程中用工难、用工贵等难题,降低蔬菜生产成本,提高蔬菜生产轻简化栽培程度。
猜你喜欢
语文周报·教研版(2021年28期)2021-08-19
农民致富之友(2020年33期)2020-12-03
安徽农业科学(2020年18期)2020-10-09
农村农业农民·B版(2018年2期)2018-03-22
中国绿色画报(2017年4期)2017-05-22
恋爱婚姻家庭·养生版(2016年5期)2016-05-06
湖北林业科技(2016年1期)2016-03-25
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
