
植物油体消化特性研究的进展
2022-02-08 03:18杨旭凤侯文淇郭本虎丁秀臻张大健赵路苹
中国粮油学报 2022年11期
杨旭凤,侯文淇,郭本虎,丁秀臻,张大健,赵路苹
(山东农业大学食品科学与工程学院1,泰安 271018) (山东农业大学作物生物学国家重点实验室2,泰安 271018)
油体是植物种子储存甘油三酯的天然细胞器。油体具有特殊结构,内部主要为甘油三酯(triglycerides、TAGs)、外部由单层磷脂和结构蛋白组成的生物膜所覆盖[1-3];结构蛋白包含油体蛋白(oleosins),油体钙蛋白(caleosins)和油体固醇蛋白(steroleosins),结构蛋白对维持油体完整性、抵抗环境压力有重要作用[4-7]。油体均匀分散到水相体系,形成较稳定的水包油(O/W)乳液[8],不需要添加乳化剂或均质,满足食品加工中绿色安全要求。此外,油体还含有一些维生素E[9]、植物甾醇[10]等功能性小分子。油体的特殊结构和这些功能性小分子使其具有良好的分散性和氧化稳定性,故油体在食品中有巨大的应用潜力。
油体作为膳食脂质形式之一,可以应用于冰激凌、人造奶油、饮料、半固体酱料等食品领域[10]。同时,油体是天然乳化剂(结构蛋白、磷脂)包裹的油滴,可以为运载功能性营养素(如姜黄素[11]、β-胡萝卜素[12])同时提供油相和天然乳化剂,故油体是功能性营养素的新型载体。为了更好的指导油体在食品中的应用,系统了解油体在胃肠道中的消化特性尤为重要。影响油体消化的因素有消化环境、油体粒径、甘油三酯类型、界面层等,其中界面层是关键因素之一;界面层可以通过种子来源、提取方式及添加多糖[13]、酚类物质[14]等调控。本文简要综述油体的组成、来源和提取方式,根据界面层组成将其分为天然界面层和复合界面层,重点介绍天然界面层和复合界面层对油体消化特性的影响规律,为油体作为膳食脂质和功能营养素载体的输送提供参考。
1 油体的组成和来源
油体广泛存在大豆、花生、油茶籽等油料作物种子中[15],直径为0.5~2.5 μm[17, 18]。Tzen等[19, 20]研究发现,油体的结构示意图如图1所示:内部为中性脂质、外部由结构蛋白及磷脂构成的单层膜覆盖,中性脂质约占油体质量的94.0%~98.0%,结构蛋白占0.6%~3.0%,磷脂占0.6%~2.0%。中性脂质主要是甘油三酯,还有少量的甘油二酯和游离脂肪酸;结构蛋白主要是分子质量为15~26 ku的油体蛋白[21-24];磷脂主要是磷脂酰胆碱(PC)和磷脂酰丝氨酸(PS),还有少量的磷脂酰乙醇胺(PE)、磷脂酰肌醇(PI)。因此,植物细胞内的油体界面层由磷脂和结构蛋白组成,也就是油体天然界面层。研究发现,油体来源对其天然界面层的组成有重要影响,不同植物种子油体天然界面层的组成见表1,如花生油体中磷脂不含有磷脂酸(PA),花生油体蛋白主要是14.5~40 ku oleosins[25];芝麻油体蛋白12.5、14 ku oleosins[26];而豆薯油体蛋白是16、22 ku oleosins[1]。这说明不同植物油体的天然界面层是不同的,油体来源是影响油体天然界面层的重要因素。
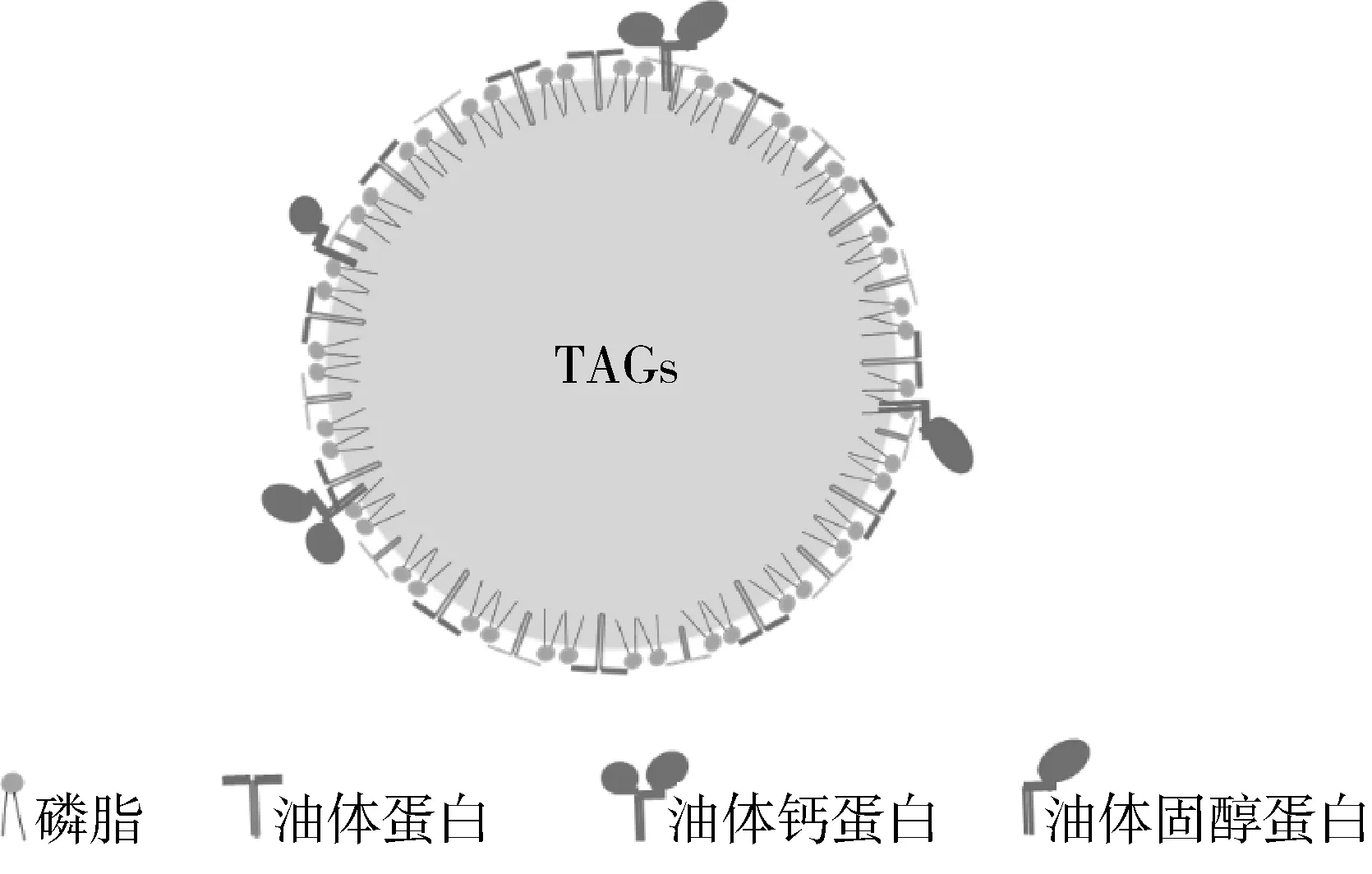
图1 油体的结构示意图

表1 不同植物种子油体天然界面层的组成
2 油体的提取
油体提取的主要过程:研磨、离心、收集。其中,研磨分为湿法研磨和干法研磨:湿法研磨是将种子浸泡在水中使其充分泡发后磨浆;干法是种子在研磨后加入水相。目前大多采用湿法研磨,通过研磨破坏植物细胞壁,蛋白质储存液泡释放出外源性蛋白质(如活性蛋白酶和贮存蛋白等),部分外源性蛋白质吸附到油体表面,离心后得到粗油体,其界面层为外源性蛋白质、磷脂和结构蛋白组成的复合界面层[35]。利用高碱(pH 11)或高浓度的尿素(8~9mol/L)洗涤粗油体,可以去除表面蛋白等杂质,得到具有天然界面层的纯油体[7]。以大豆油体为例,列举不同提取条件对大豆油体界面蛋白的影响,不同提取条件对大豆油体外源性蛋白质的影响见表2。Ishii等[36, 37]在不同pH条件下提取大豆油体,得到2种油体,纯油体(24、18 ku oleosins)和粗油体,粗油体中除了含有油体蛋白外,油体表面还吸附β-伴球蛋白(7S)和大豆球蛋白(11S)等外源性蛋白质。闫尊浩[38]和赵路苹等[39]在不同温度下提取大豆油体,研究温度对油体表面蛋白的影响,温度升高至70℃时外源性蛋白质开始变性并从油体表面解离,温度越高,外源性蛋白质含量越少。这说明提取条件主要改变油体界面吸附的外源性蛋白质组成。

表2 不同提取条件对大豆油体外源性蛋白质的影响
3 油体的消化特性
油体在人体中消化的大致过程:液滴经过口腔后,经胃液初步乳化[43],胃蛋白酶水解界面蛋白质;传递到小肠,胆盐取代界面磷脂分子和肽,脂肪酶和辅酶结合到液滴表面,不断水解油脂,并释放有益小分子。界面层是油体消化的首要屏障,然而油体来源、提取方式以及油体与食品组分的相互作用会改变界面层组成。本文将界面层分为天然界面层和复合界面层,讨论对油体消化特性的影响。天然界面层是由磷脂和结构蛋白组成,复合界面层是天然界面层与外源性蛋白质、多糖、酚类物质等组合构成。
3.1 天然界面层的油体消化特性
提取具有天然界面层的纯油体主要是在pH 11.0或高浓度尿素条件下多次洗涤、离心得到。White等[27]用9 mol/L尿素提取了含脂质99.2%、蛋白质1.5%的葵花籽纯油体(油体蛋白为18~21 ku),并采用胃-肠二阶段模型研究其消化特性。在胃消化阶段,胃蛋白酶水解18~21 ku油体蛋白产生6.5~14 ku肽段,液滴之间发生融合,粒径由2.3 μm增大到22.5 μm,粒度分布也由单峰变为三峰;由于液滴粒径增大,比表面积减小,胃蛋白酶作用位点减少,导致蛋白水解速率降低。在肠消化阶段,一方面液滴比表面积减小导致脂肪酶的作用位点减少;另一方面油体的磷脂-结构蛋白层会降低脂肪酶的活性[44]。最终肠消化后,维生素E和总脂肪酸的生物可及性仅为0.6%和8.3%。目前对于天然油体消化特性的研究报道的仅有葵花籽纯油体,对于其他天然油体尚未见报道。
3.2 复合界面层的油体消化特性
3.2.1 外源性蛋白质对油体消化特性的影响
在较温和的水相提取条件(如中性pH、60~80 ℃加热等)下得到的多数油体是含有外源性蛋白质的粗油体。Sophie等[28]湿法磨浆过滤(150 μm)得到含有核桃油体和蛋白的核桃乳,并考察了核桃油体胃肠道消化特性。核桃油体粒径呈双峰分布,分别在0.5~1.4 μm和1.4~30 μm。在胃消化阶段,液滴粒度分布向较大尺寸(10~275 μm)移动,这是由于环境的pH为1.2,以及胃蛋白酶不断水解油体表面蛋白质而导致液滴无法被肽段完全覆盖,致使油体界面不稳定,形成聚集体;在消化60 min时,部分谷蛋白(21、33、37 ku)仍存在并产生了一些胃蛋白酶抗性肽(<6.5 ku)。在肠消化阶段时(pH7.5),抗性多肽被肠中胰蛋白酶水解;带负电荷的胆盐吸附到液滴界面,带电荷量增加,聚集体逐渐分散;胆盐置换界面上的磷脂并与磷脂形成混合胶束,胰脂肪酶水解甘油三酯的产物在胶束中运输,并形成一种新型水包油包水乳液。肠中脂肪水解的速度在最初10 min最快,脂肪水解产物在界面上的积累阻碍了胰脂肪酶接近内部的甘油三酯,后期消化速率减慢,核桃油体总游离脂肪酸的释放率是110 μmol/mL。因此,该研究中,核桃油体在胃消化阶段出现抗性肽;在肠消化阶段,抗性肽被胰蛋白酶水解,脂肪水解过程中形成胶束和新型多重乳液,推测可能是由于界面蛋白质水解的不完全促进了这种结构的产生。
Sophie等[29, 45]湿法磨浆过滤(150 μm)得到含有杏仁油体和蛋白的杏仁乳,探究该杏仁油体的胃肠道消化特性。在胃消化阶段,液滴粒径由3 μm增加到30 μm,呈单峰分布;胃蛋白酶将杏仁蛋白水解成低分子量的肽,并出现胃蛋白酶抗性肽(8 ku)。肠消化阶段(pH 7.5),胰蛋白酶水解胃阶段产生的抗性肽;加入胆盐后,液滴表面静电斥力增加,出现较小颗粒(0.6~1.2 μm),同时胆盐置换液滴表面磷脂并与磷脂形成混合胶束和囊泡;加入胰酶后,胰脂肪酶水解甘油三酯的产物由胶束从油相输到水相。由于液滴中的蛋白质和油脂被水解,液滴表面不能被完全覆盖,出现较少的聚集体,液滴粒径变为三峰分布(0.6~1.2 μm,6~45 μm,50~500 μm)。肠消化初期,胰脂肪酶缓慢渗透到油-水界面,这延迟了胰脂肪酶水解甘油三脂,可以通过胃消化期间加入胃脂肪酶,脂肪水解产物有助于胰脂肪酶的结合和活化,从而实现消除延迟现象[46]。25 min时,胰脂肪酶到达油-水界面迅速水解甘油三酯,游离脂肪酸的释放速率增加,随后游离脂肪酸在胶束运输时,界面浓度饱和使其释放速率减慢,肠消化结束时游离脂肪酸的释放率为133 μmol/mL。综上,杏仁油体在胃消化过程中同样有胃蛋白酶抗性肽的形成;进入肠消化后,这些抗性肽在肠中被胰蛋白酶水解,胆盐通过胶束去除积聚在液滴表面的脂肪水解产物来促进脂肪水解。
He等[47]等利用蔗糖和Tris-HCl缓冲溶液(pH 7.5)提取大豆油体,模拟体外胃肠道消化。在胃消化阶段,胃蛋白酶不断水解油体蛋白(24、18 ku)和外源性蛋白质(7S和11S),24 ku油体蛋白在30 min内被彻底水解,18 ku油体蛋白随着消化时间的延长最终被水解;外源性蛋白质部分被水解。蛋白水解使油体表面结构被破坏,且环境的pH为1.5,所以此阶段液滴粒径增大。进入肠消化阶段,所有蛋白被迅速水解;甘油三酯在胰脂肪酶的作用下被水解成游离脂肪酸和单甘油酯,液滴被分解成小液滴,粒径减小。游离脂肪酸的释放速率呈现先增加后减小的趋势,游离脂肪酸的释放率为55.3 μmol/mL。此外,Liu等[12]利用大豆油体包埋姜黄素,并考察了其输送特性。用4%的大豆油体包埋姜黄素,液滴的平均直径为2.7 μm。胃消化阶段,粒径先增大(45.8 μm)后减小(21.87 μm),≤25 ku的蛋白质在60 min时被胃蛋白酶完全水解,而45~66.2 ku的蛋白仅发生部分水解,所以包埋姜黄素后的大豆油体中分子量较大的蛋白质水解程度不高。进入肠消化阶段,液滴逐渐分散,消化180 min后,游离脂肪酸的释放率为59.6 μmol/mL,姜黄素的释放率达到54%。
He等[48]还研究了油菜籽油体(水分36.86%,脂质88.98%,蛋白质7.90%)的消化特性。油菜籽油体中外源性蛋白分布在30~116 ku[49]。在胃消化阶段,随着消化时间的延长,部分蛋白质被水解,油体界面结构被破坏,液滴粒径逐渐增大,粒径分布范围扩大。进入肠消化阶段,蛋白质被全部水解,分散性增强,粒径减小。游离脂肪酸的释放率为31.75 μmol/mL。
油体天然界面层由磷脂和结构蛋白组成,外源性蛋白质吸附到油体界面的示意图如图2a所示,不同植物油体的天然界面层的组成不同;同时,不同提取方式也会影响油体的界面组成,进而影响油体的消化特性。图2b为外源性蛋白质吸附到天然油体界面,构成外源性蛋白质、磷脂、结构蛋白复合界面层,在胃消化中某些外源性蛋白质水解产生抗性多肽,这些抗性多肽被肠中胰蛋白酶水解,且肠消化阶段会产生胶束和新型多重乳液,影响甘油三酯的消化。
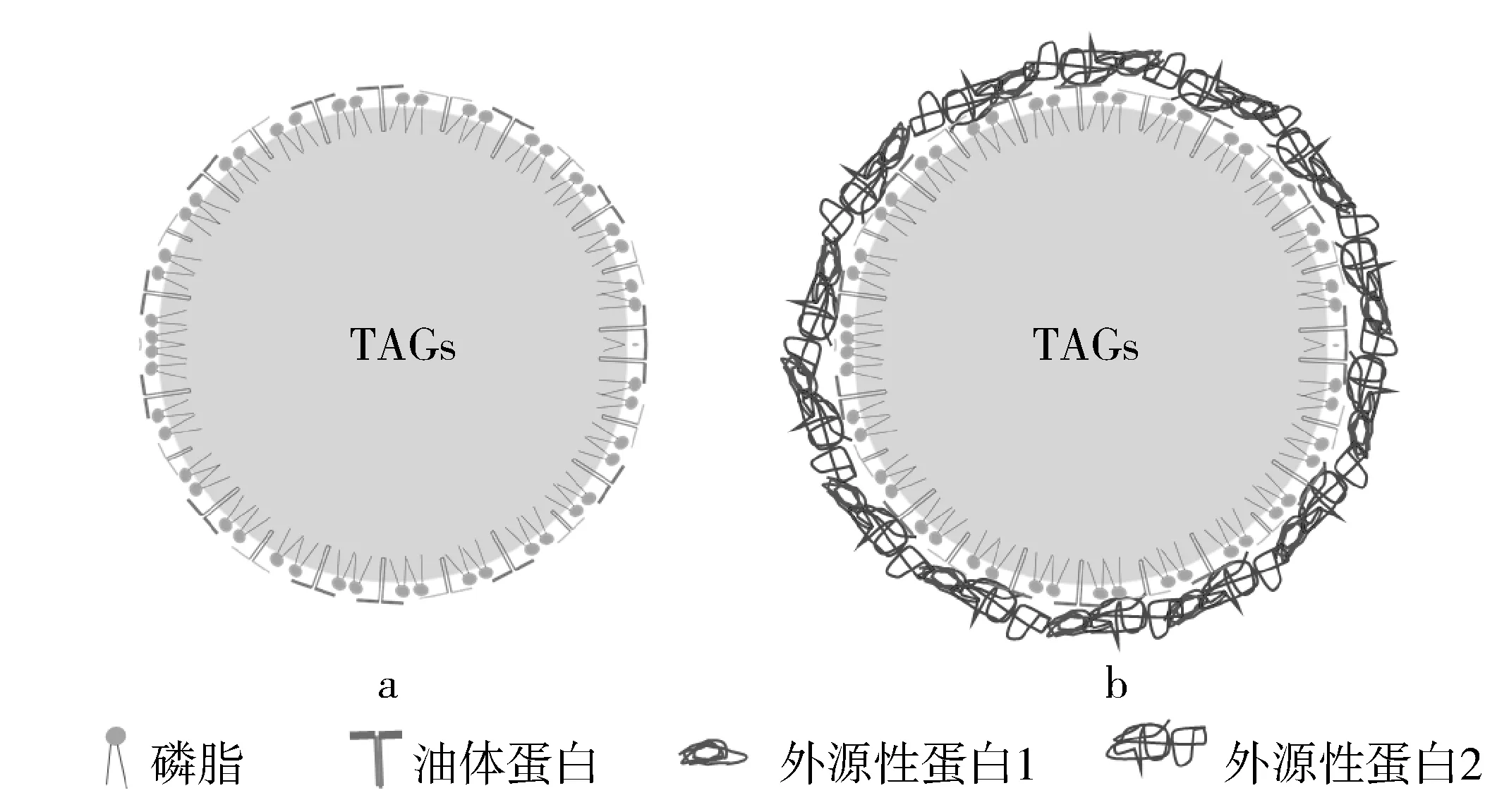
图2 外源性蛋白质吸附到油体界面的示意图
3.2.2 多糖对油体消化特性的影响
多糖具有良好的物理和化学稳定性,添加到蛋白质稳定的乳液中会形成多糖-蛋白质界面层,进而影响液滴的带电荷量、黏度等界面性质;此外,多糖可以在水相中形成网络结构,限制油滴流动,影响蛋白质稳定的乳液性质[50-52]。油体与蛋白质稳定的乳液液滴类似,利用多糖也可影响油体稳定性。
Wu等[13]研究了不同浓度卡拉胶-大豆油体的消化特性,利用NaCl和蔗糖的Tris-HCl缓冲溶液(pH 7.5)提取大豆油体,不同质量分数的ι-卡拉胶(0.05%、0.1%、0.2%、0.4%)包裹的大豆油体,多糖吸附到油体界面的示意图如图3a所示,液滴粒径分布范围在6~108 μm。胃消化60 min后,ι-卡拉胶质量分数为0.05%、0.1%时,液滴直径分别增加了54、25 μm,这是由于在低ι-卡拉胶浓度下,大豆油体蛋白仍可以被彻底水解,出现小于14.4 ku蛋白条带,蛋白水解导致油体液滴不稳定,发生聚集;当质量分数为0.2%、0.4% ι-卡拉胶-大豆油体经过胃消化后,仅有部分蛋白质被水解,液滴直径没有明显变化,这说明高浓度的ι-卡拉胶阻碍了胃蛋白酶接触液滴表面的作用位点,抑制了酶的水解作用;肠消化阶段,高浓度的ι-卡拉胶阻碍了脂肪酶移动到反应位点;此外,ι-卡拉胶可能会与钙离子络合,从而影响脂肪的水解速率,随着ι-卡拉胶质量分数的升高,脂肪酸的生物利用率从62.7%下降到8.3%。
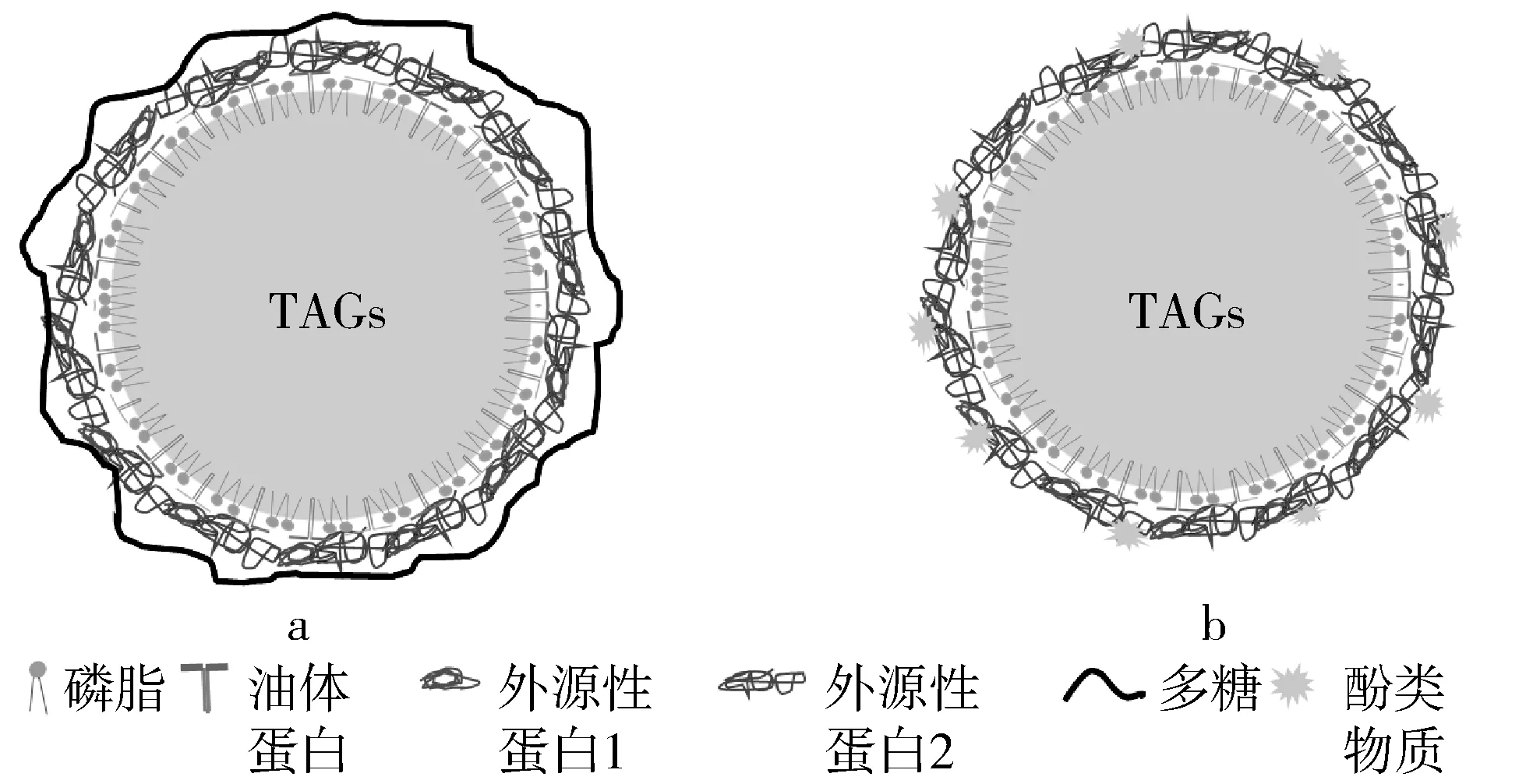
图3 多糖和酚类物质吸附到油体界面的示意图
Ding等[53]利用麦芽糊精(MD)、壳聚糖(CS)和表没食子儿茶素没食子酸酯(EGCG)偶联物包裹的大豆油体,研究其消化特性。CS通过静电作用吸附在油体表面,EGCG可以提高壳聚糖的乳化活性和抗氧化活性,二者偶联(CSEG)与MD混合形成MD-CSEG,利用喷雾干燥法封装大豆油体,油体微粒尺寸减小,界面层的稳定性增强。胃消化阶段,环境的pH为2,CS更加亲水,被紧密包裹的大豆油体界面蛋白不能被快速水解。肠消化中,环境的pH上升至7,CS失去正电荷,变得疏水,液滴不稳定发生聚集,阻碍了胰脂肪酶的扩散和吸附,抑制了脂肪的水解,最终测得油的释放率从85%下降到了40%。
郭安民[54]研究了羟丙基甲基纤维素(HPMC)-大豆油体的消化特性。HPMC浓度小于0.4%时,胃消化阶段油体界面蛋白被水解,液滴直径明显增大;浓度大于0.4%的HPMC-大豆油体在胃消化期间未发现明显的聚集,原因是较高浓度的HPMC作为一种非离子型的纤维素醚,能够形成致密的网状结构,阻碍了消化酶接触油体内部,抑制了油体的消化。肠中游离脂肪酸的释放率随着HPMC浓度的升高而下降,0.8% HPMC-大豆油体游离脂肪酸的释放率下降到30%。
3.2.3 酚类物质对油体消化特性的影响
酚类物质可以与油体界面蛋白结合,提高油体的稳定性,影响消化特性。官梦姝等[14]研究了3种天然酚类物质(儿茶素、白藜芦醇及没食子酸)对大豆油体消化特性的影响。由于酚类物质与油体界面蛋白络合,酚类物质吸附到油体界面的示意图如图3b所示,形成的络合物之间又相互交联,促进油体的聚集,这减缓了胃蛋白酶对油体界面蛋白的水解。随着酚类物质浓度的增加,未被水解的液滴越来越多。胃中较大液滴经过肠消化后被分解为小液滴,但酚类物质-油体复合体系在肠消化仍然保持了较低的消化程度,这是由于酚类物质通过疏水作用与胰蛋白酶结合,破坏其二级结构,抑制酶的活性,减缓了油脂的水解。分别添加500 μmol/L的儿茶素、白藜芦醇和没食子酸,游离脂肪酸的释放下降约22.9%、18.1%和10.7%。
油体的来源、提取方式及外源物质的添加都会改变油体的界面层组成和性质,进而影响油体的消化规律,不同界面层对油体消化特性的影响见表3。天然界面层由磷脂和结构蛋白组成,天然的界面结构会降低甘油三酯的消化率;复合界面层是由外源性蛋白质、多糖或酚类物质与油体的天然界面组成,外源性蛋白质和其他物质的加入会影响油体的消化。油体消化特性可以为油体作为功能营养素载体及相关产品开发提供一定的指导。

表3 不同界面层对油体消化特性的影响
4 展望
油体是一种天然的乳化油滴粒子,作为膳食脂质或功能性营养素的载体在食品、化妆品等领域引起了很多学者的关注。不同油体的界面层组成不同,在不同提取方式及其他添加物质的作用下也会改变油体界面层的组成和性质,影响油体的稳定性及消化特性。然而,油体界面层组分间的相互作用方式还需更深入的研究。此外,关于油体体外消化研究最多的是以胃-肠二阶段静态模型,尽管口腔中没有能够水解脂肪的酶,口腔中的pH、离子强度及温度有助于乳液的初步分散,可能影响油体消化。通过调控油体界面层系统研究油体的消化特性,实现功能营养素的有效释放也是未来主要研究目标。
猜你喜欢
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2019年12期)2020-01-04
中成药(2018年7期)2018-08-04
中成药(2017年12期)2018-01-19
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中国粮油学报(2016年5期)2016-01-23
中国卫生标准管理(2015年18期)2016-01-20
中国粮油学报(2014年6期)2014-12-27
中国酿造(2014年9期)2014-03-11
