
多次交配对小菜蛾精包形成与生殖力的影响
2022-02-08 08:18邹明民刘莉莉董诗杰黄梦琪曹敏慧尤民生
昆虫学报 2022年12期
邹明民,刘莉莉,董诗杰,黄梦琪,曹敏慧,尤民生,*,彭 露,*
(1.福建农林大学应用生态研究所,闽台作物有害生物生态防控国家重点实验室,福州 350002;2.广东省农业科学院作物研究所,广州 510640;3.广东省农作物遗传改良重点实验室,广州 510640;4.福建农林大学海峡两岸特色作物安全生产省部共建协同创新中心,福州 350002;5.福建农林大学,教育部害虫生态防控国际合作联合实验室,福州 350002;6.福建农林大学,福建省昆虫生态重点实验室,福州 350002)
在进化过程中,营两性生殖的不同昆虫种类发展出不同的交配策略以获得对亲代或后代最大的生殖利益。昆虫采取的交配策略通常包括3种形式(Price PWetal.,2011;张诗语等,2015):(1)单配制(monogamy),雌雄两性一生只交配一次。这种交配策略多见于雌雄两性均具有亲代抚育行为的昆虫,如葬甲科(Silphidae)甲虫。(2)多配制(polygamy):可细分为一雌多雄(polyandry)和一雄多雌(polygyny)两种形式。其中,一雌多雄制即雌性能与多个配偶交配,而雄性通常只交配一次。这种交配策略在动物界中是最稀少的,多见于具有性食同类(sexual cannibalism)行为的物种中,例如亚利桑那螳螂Stagmomantislimbata(Maxwelletal.,2010);与之相反的,一雄多雌交配策略是动物界中最常见的形式,这类雌虫可能具有守卫(defending)和求偶行为(Alcocketal.,1976)。(3)混交制(promiscuity),雌雄两性均能与多个配偶交配。越来越多的研究表明,雌雄的多次交配行为广泛存在于昆虫类群中,主要有鞘翅目、膜翅目、鳞翅目、直翅目、双翅目、半翅目、长翅目、广翅目等昆虫种类(Arnqvistetal.,2000;刘兴平等,2008)。
交配策略具有可塑性,即同一个物种因地理种群(Välimäkietal.,2006;Välimäki and Kaitala,2006)、生态环境(McFarlaneetal.,2011)、个体生理状态(Syriatowicz and Brooks,2004;Frickeetal.,2010)和遗传基础(Harano and Miyatake,2005;Välimäki and Kaitala,2006;Price TARetal.,2011)等因素的差异,其种群或个体间往往会表现出不一样的交配策略。例如,研究人员曾分别对来自英国(Smith,1956)、西班牙(Holmanetal.,2008)和加拿大(Lizéetal.,2012)的Drosophilasubobscura交配行为进行了研究,结果均表明D.subobscura雌虫是严格的一夫制昆虫;但Loukas等(1981)在对希腊种群进行研究时却发现有20%的雌虫可以再次交配,并推测D.subobscura雌虫为多夫制昆虫。随后,Fisher等(2013)则观测比较了英国和希腊种群D.subobscura雌虫在不同实验条件下的再交配率,结果发现平均14.4%的雌虫具有多次交配行为,而不同地理种群雌虫再交配率没有显著差异,但两个种群雌虫的再交配行为对温度变化的响应却有很大差别。Kaňuch等(2013)比较了来自北欧16个地点的灌丛蟋蟀Metriopteraroeselii雌虫的再交配率,结果发现来自高纬度种群雌虫的再交配次数普遍多于低纬度地区的;同时雌虫的体型也影响其交配次数。考虑到交配策略的可塑性,目前通常根据一个种群雌虫的交配频率或多次交配的比例来界定昆虫的交配策略类型:若一个种群中雌虫的平均交配次数大于1.5次或雌虫多次交配的比例大于40%,则认为雌虫是实行多夫制交配策略(Arnqvistetal.,2000;Torres-Vilaetal.,2004)。对于小菜蛾的交配策略,不同学者仍持有不同观点,且多次交配实验中仅观察了雌虫与同一头雄虫的重复交配情况(Wangetal.,2005;谭永安等,2011;Songetal.,2014),同时多次交配是否带来生殖效益仍没有定论。
昆虫的多次交配对雌雄两性而言都是一个生殖利益和代价共存的行为,雄虫的生殖成功与其交配次数密切相关,但是精液的输出对雄虫来说是一个巨大的能量损耗(Duplouyetal.,2018);交配也可引起雌虫多种生理和行为改变,如:抑制雌虫再交配、刺激雌虫排卵、提高雌虫产卵量和孵化率以及促进卵巢发育和卵子发生等(Avilaetal.,2011;Guédotetal.,2013;Shahidetal.,2016)。交配过程的实质是精液的输出与接收,精液中除精子外,还包含蛋白、盐分、糖类、防御物质、脂质、水和微生物等组分(Poiani,2006)。在多数昆虫中,精液会以精包的形式暂时储存在交配囊中,精包的完全消耗和降解是雌虫可再次交配的信号之一,如灶蟋Gryllodessigillatus(Batemanetal.,2001)、烟芽夜蛾Heliothisvirescens(LaMunyon and Huffman,2001)、斜眼褐蝶Bicyclusanynana(Kehletal.,2013);但也有一些昆虫交配后只会形成交配塞,如黑腹果蝇Drosophilamelanogaster(Lung and Wolfner,1999);还有一些昆虫在交配过程中,雄虫生殖器会刺穿雌虫的生殖道或表皮,精液则直接通过伤口进入雌虫体内,如Drosophilaeugracilis(Kamimura,2010)。然而,在交配过程中,小菜蛾雄虫精液是通过什么形式进入到雌虫体内目前尚未定论。
小菜蛾是为害十字花科蔬菜最为严重的世界性害虫,每年造成的损失以及防治费用总计可高达40亿~50亿美元(Furlongetal.,2013)。小菜蛾强大的生殖潜力以及抗药性的迅速发展使其成为田间最难防治的害虫之一(Sunithaetal.,2020)。对于营两性繁殖的昆虫而言,交配是其繁育后代实现种群增长的一个必要生理过程。本研究旨在明确小菜蛾G88敏感品系成虫的交配行为及交配后的生理响应,如:是否形成精包结构,雄虫多次交配下其精液含量变化及其对雌虫生殖力的影响,雌虫是否具有多次交配行为,交配对雌虫再交配的抑制时长,以及多次交配是否会给雌虫带来生殖效应等。研究结果可为雌虫交配后的生殖行为和生理变化以及雄虫在生殖调控中的分子机制作用奠定基础,对于小菜蛾种群监测和综合防治具有重要意义。
1 材料与方法
1.1 供试昆虫
本研究使用实验室长期饲养的小菜蛾G88敏感品系,该品系采用人工饲料饲养。幼虫于7.3 cm(杯底直径)×10.4 cm(杯口直径)×8.5 cm(高)的纸杯中取食人工饲料直至化蛹,饲养条件为:温度25±1℃,RH 70%±5%,光周期16L∶8D。种群延续:将50~80个蛹放入纸杯中,羽化后的成虫可自由交配,杯盖开口并塞入用10%的蜂蜜水浸湿的棉花,为小菜蛾成虫提供营养补充。实验用虫:将蛹分装到1.5 mL离心管中(1头/管)单性饲养直至羽化,羽化后第1天记为1日龄,第2天为2日龄,以此类推。
1.2 雌成虫生殖系统结构观察
1.2.1雌性生殖腺形态观察:将2日龄未交配的小菜蛾成虫放入3.0 cm(杯底直径)×4.0 cm(杯口直径)×3.0 cm(高)的透明塑料杯中进行单对配对,待其交配完成时立即用CO2麻醉,将雌虫腹部用解剖剪剪下并置于1×PBS溶液中。在显微镜下完成解剖并分离出雌成虫生殖腺,使用显微镜自带软件进行拍照。使用同样的方式解剖、拍照和观察未交配雌成虫生殖腺。
1.2.2精包结构冷冻切片观察:解剖和分离交配后雌成虫交配囊,随后立即将样本包埋入OTC包埋剂(SAKURA)中。于-80℃中放置12 h后,使用冷冻切片机(Shandon Cryotome FSE)进行5 μm厚度半薄切片。将切片移入干净载玻片上,使用激光共聚焦显微镜(Leica SP5)进行拍摄。
1.3 交配行为特性和生殖力观测
1.3.1交配能力测定:将2日龄未交配的小菜蛾成虫分别放入塑料杯中进行单对配对,每20 min观察并记录一次交配情况,持续12 h。待交配结束后,立即将雌雄分开并置于新的塑料杯中。而后在杯中放入2日龄未交配的另一性别成虫,每20 min中观察并记录交配情况,持续12 h。该试验设3次重复,每个重复处理20对小菜蛾成虫。
1.3.2雌成虫精包的消耗:将45对2日龄未交配的小菜蛾成虫分别放入塑料杯中进行单对配对,每20 min观察一次交配情况。待交配结束后,立即取出雌成虫,解剖交配后2 h 和12 h的雌成虫个体(交配后不同时间各15头),分离出交配囊,拍照观察交配囊内精包的消耗情况。
1.3.3多次交配对雄成虫射精量的影响:将30对2日龄未交配的小菜蛾成虫分别放入塑料杯中进行单对配对,观察并记录每对试虫的交配时长。待交配结束后,取出雄成虫做两种处理:(1)将15头雄成虫置于新的塑料杯中,放入2日龄未交配的雌成虫进行单对配对,观察并记录每对试虫的交配时长;(2)其余15头雄成虫单性饲养于新的塑料杯中,24 h后放入2日龄未交配的雌成虫进行单对配对,观察并记录每对试虫的交配时长。在交配结束后,立即解剖并分离出所有供试雌成虫的交配囊,拍照并计算精包大小(面积)。
1.3.4多次交配对雌成虫生殖力的影响:将单对2日龄未交配的小菜蛾成虫分别放入塑料杯中进行配对,每20 min观察一次交配情况。交配结束后,取出雌虫做两种处理:(1)放入新的2日龄未交配雄成虫,每20 min观察一次交配情况,持续12 h。期间观测到再交配行为的雌成虫个体记为有效的多次交配雌成虫。记录15头有效多配雌成虫交配后3 d内的产卵量和卵孵化率,观察期间不移走雄成虫;(2)另15头雌虫单性饲养于新的塑料杯中,记录每头雌成虫交配后3 d内的产卵量和卵孵化率,3 d后解剖并分离出所有供试雌成虫的交配囊,拍照并计算交配囊内精包的个数。
1.4 数据分析
使用SPSS 19.0软件进行数据统计分析。多组数据比较采用单因素方差分析(one-way ANOVA)和Tukey HSD检验;两组数据比较采用独立样本t检验。
2 结果
2.1 雌成虫生殖系统结构
未交配小菜蛾雌成虫交配囊大约呈圆形扁平状,半透明状,交配囊内未发现Signa结构(Sánchezetal.,2011);受精囊呈管状,细长型,受精囊腺位于顶端,呈椭圆型。交配后,雌虫交配囊明显膨胀变大,呈圆球状。交配囊腔内存在精包结构,呈白色,不透明球状。受精囊内已有精液存在,说明交配短时间内精液即可从精包内转移到受精囊中(图1)。为了进一步验证小菜蛾精包结构,对交配后雌成虫交配囊进行冷冻切片和观察,发现小菜蛾交配囊膜厚度不均,表面粗糙状;紧贴交配囊膜内侧还存在一层厚度均匀的包膜结构,此结构即为精包的包膜外层(envelope)(图2)。
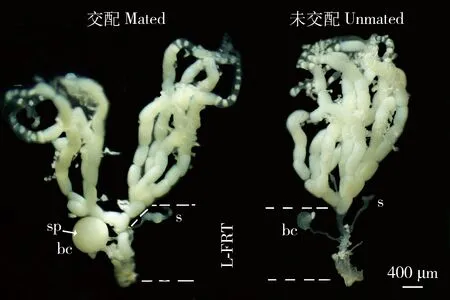
图1 小菜蛾雌成虫生殖腺结构Fig.1 Structure of reproductive gonads of female adults of Plutella xylostellabc:交配囊Bursa copulatrix;sp:精包Spermatophore;s:受精囊Spermatheca;L-FRT:下生殖道Lower reproductive tract.
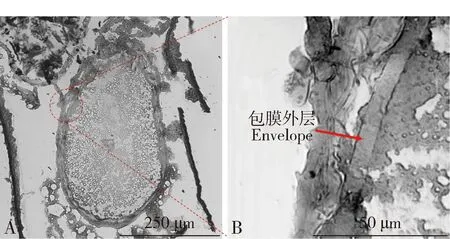
图2 小菜蛾雌成虫交配囊冷冻切片Fig.2 Frozen section of bursa copulatrix of female adults of Plutella xylostellaA:交配囊的冷冻切片Frozen section of bursa copulatrix;B:包膜外层的放大图Enlarged view of the envelope.红色箭头所示为精包的包膜外层。Red arrow indicates the envelope of spermatophore.
2.2 成虫交配能力及首次交配后精包的消化情况
小菜蛾2日龄成虫首次交配的响应时间短、成功率高,在20 min,2 h和12 h观测时间内,其交配成功率分别为86.7%,97.8%和97.8%(图3)。首次交配结束后,小菜蛾雄成虫表现出短暂的再交配延迟,20 min内其交配成功率为54.6%,显著低于首次交配时的(t=4.11,df=4,P=0.015)。而在首次交配后2 h和12 h内,雄成虫再交配成功率分别为86.9%和100%,与其首次交配时无显著差异(2 h:t=1.000,df=4,P=0.091;12h:t=1.000,df=4,P=0.374)。小菜蛾雌成虫则表现出较弱的连续交配能力,在首次交配后 20 min,2 h和12 h内雌虫的交配率分别为26.6%,64.6%和83.4%,均显著低于雌虫首次交配时的(20 min:t=5.177,df=4,P=0.007;2 h:t=7.460,df=4,P=0.002;12 h:t=4.325,df=4,P=0.012)(图3)。结果说明,小菜蛾雌雄成虫均具有多次交配行为特性,而交配对雌成虫再交配的抑制作用更加明显。
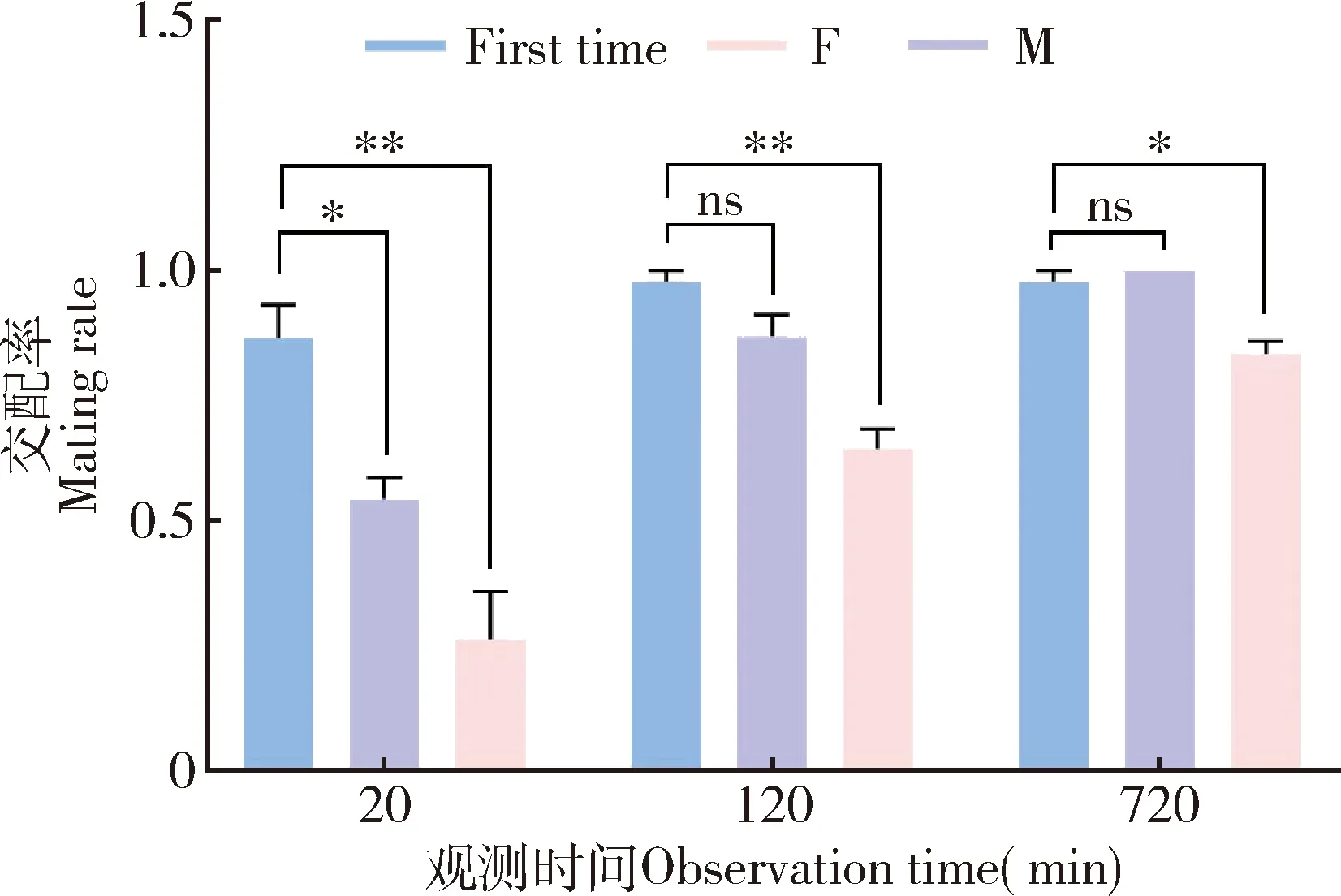
图3 小菜蛾成虫交配能力Fig.3 Mating capability of Plutella xylostella adultsFirst time:成虫首次交配First mating of the adults;M:雄虫第2次交配Second mating of the male;F:雌虫第2次交配Second mating of the female.图中数据为平均值±标准误;星号表示两组间差异显著(*P<0.05;**P<0.01),ns表示两组间差异不显著(P>0.05)(t检验)。Data in the figure are mean±SE.Asterisks indicate significant differences between two groups (*P<0.05;**P<0.01),and ns indicate no significant differences between two groups (P>0.05)(t-test).
昆虫精包的充分消化和吸收是雌虫再交配的前提之一(Oberhauser,1989)。为了探究小菜蛾雌成虫再次交配的生理基础,对其首次交配后精包的消化情况进行了观察。交配后2 h,雌成虫交配囊内的精包已有明显的消耗,其中有40%(6/15)雌虫的精包已缩小成白色小团块,交配囊恢复成半透明状;其余60%(9/15)雌虫的精液仍充满整个交配囊,但交配囊有明显皱缩。在交配后12 h,所有雌成虫交配囊内的精包已缩小成白色小团块,交配囊已恢复成半透明状(图4)。结果说明,雌成虫再交配可能取决于首次交配后精包的消化和吸收速率。
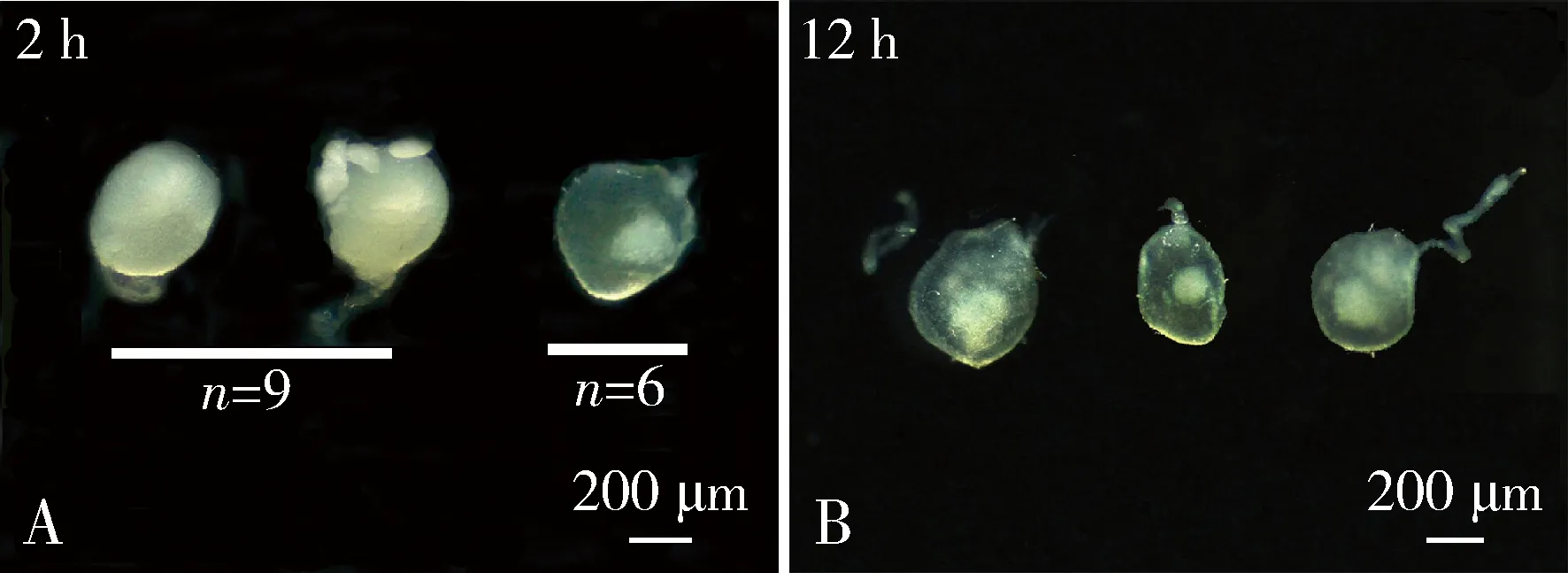
图4 小菜蛾成虫交配后2 h(A)和12 h(B)的精包形态Fig.4 Morphology of spermatophore of Plutella xylostella adults at 2 h (A) and 12 h (B) after mating
2.3 雄成虫不同交配次数对精包形成和雌成虫生殖力的影响
雄成虫不同交配次数并不影响交配时长,3组试虫的交配时长(M1:72.07±2.56;M2:78.60±3.29;M2-24:72.93±3.22)间没有显著差异(F2,42=1.35,P=0.268)(图5:A)。雄成虫连续交配显著影响交配后所形成的精包大小(F3,56=152.49,P<0.001)(图5:B,C)。雄成虫首次交配(M1)所形成的精包显著大于雄成虫第2次交配(M2:首次交配后立即进行;M2-24:首次交配后恢复24 h)(P<0.05)。M2-24雄虫所形成的精包虽然相应增大,但与M2相比精包大小没有显著差异(P=0.847)。结果说明,雄成虫第2次交配时的射精量(精包大小)明显减少,并且很难得到恢复补充。
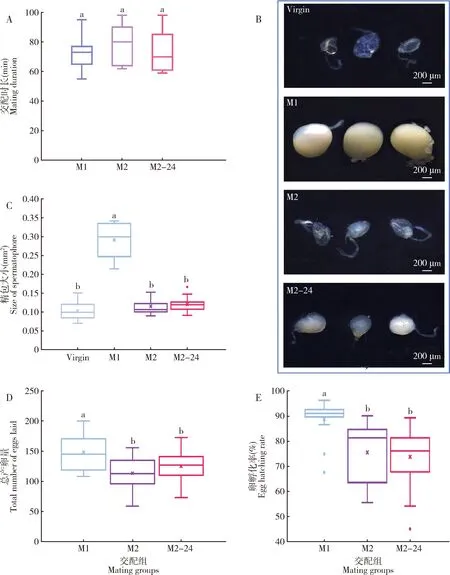
图5 小菜蛾雄成虫交配次数对精包形成和雌虫生殖力的影响Fig.5 Effects of mating frequency of male adults on the spermatophore formation and female fecundity of Plutella xylostellaA:交配时长Mating duration;B:精包形态Morphology of spermatophore;C:精包大小Size of spermatophore;D:交配后3 d内的总产卵量Total number of eggs laid within 3 d after mating;E:卵孵化率Egg hatching rate.Virgin:未交配Unmated;M1:第1次交配First mating;M2:第2次交配(首次交配后立即进行)Second mating (immediately after the first mating);M2-24:第2次交配(首次交配后恢复 24 h)Second mating (recovery for 24 h after the first mating).图中数据为平均值±标准误;不同字母表示经Tukey HSD检验分析在0.05水平差异显著。Data in the figure are mean±SE.Different letters mean significant difference at the 0.05 level by Tukey HSD test.
雄成虫不同交配次数下雌成虫的产卵量存在显著差异,雄成虫首次交配所诱导的雌成虫产卵量(M1:148.53±6.99)显著高于雄成虫第2次交配时的(M2:113.53±6.87;M2-24:125.06±6.33)(F2,42=7.00,P=0.002)(图5:D)。相比于连续交配,经过24 h恢复的雄成虫交配后所诱导的雌成虫产卵量没有显著差异(P=0.454)。雄成虫不同交配次数下雌虫的卵孵化率同样存在显著差异。雄成虫首次交配所诱导的雌成虫卵孵化率(M1:89%)显著高于雄成虫第2次交配时的(M2:76%;M2-24:74%)(F2,42=8.43,P=0.001)(图5:E)。结果说明,小菜蛾雄成虫交配史对雌成虫的生殖力有明显的影响。
2.4 雌成虫多次交配后的精包形成和生殖力
为了验证雌成虫多次交配后的生殖力变化,我们进行了雌成虫与不同雄成虫的多次交配试验。小菜蛾的交配次数可以通过交配后交配囊囊内精包的数量来证实(图6:A)。交配后3 d时间内,多配组小菜蛾雌成虫的平均交配次数是4.27次,但多次交配并没有显著影响雌成虫的产卵量(t=0.424,df=28,P=0.675)(图6:B)和卵孵化率(t=0.021,df=28,P=0.983)(图6:C)。结果表明,小菜蛾雌成虫多次交配并没有获得产卵量和孵化率的收益,雌成虫只交配1次,就可达到75%以上的产卵量。
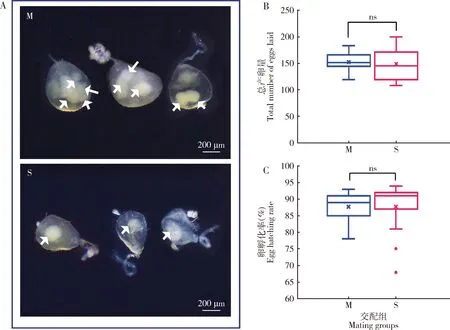
图6 小菜蛾雌成虫多次交配对精包形成和雌虫生殖力的影响Fig.6 Effect of multiple mating of female adults of Plutella xylostella on the spermatophore formation and female fecundityA:交配囊Bursa copulatrix (示精包数量不同Showing the number of spermatophores different);B:交配后3 d内的总产卵量Total number of eggs laid within 3 d after mating;C:卵孵化率Egg hatching rate.M:多次交配Multiple mating;S:单次交配Single mating.白色箭头所示为精包。图中数据为平均值±标准误;ns表示两组间差异不显著(P>0.05,t检验)。Spermatophore is indicated by white arrow.Data in the figure are mean±SE,and ns indicates no significant differences between two group (P>0.05,t-test).
3 讨论
在许多昆虫交配过程中,雄成虫精液进入雌成虫交配囊后会形成一个具有坚硬包膜外层的精包结构。在G88敏感品系小菜蛾中,同样发现精包结构的存在,其形态与Yang和Chow(1978)描述的一致。但Justus和Mitchell(1999)在实验中却并未发现类似结构,他们认为小菜蛾精液不形成精包,而是在雌虫体外囊管末端开口处硬化形成交配塞。这种外部交配塞多见于鳞翅目蝴蝶类昆虫中,它通常是一个临时性的结构,起到延迟雌虫再交配行为的作用。而在一些物种中,外部交配塞也可以永久性地存在于雌虫腹部,因此也将其称为封瓣(sphragis)(Carvalhoetal.,2017,2019)。是否不同种群小菜蛾精液组分的差异导致了不同结构的形成,有待进一步的研究。同时,结合本研究精包消化和吸收的观察结果,推测这种差异也可能是由于观测时间不同导致的。
小菜蛾雄成虫具有多次交配的行为,但其生殖力随着交配次数的增加显著降低,其原因是雄虫提供精液的能力逐次降低,并且其射精量很难得到恢复,该结果与Wang等(2005)结果一致。张诗语等(2016)对7科27种蛾类的实验数据进行Meta分析结果也表明,从总体而言,雄蛾的交配史对其自身的精包大小以及雌虫的生殖力均具有明显的负作用。雄性昆虫还可根据雌虫的交配状态调整自身射精含量或组分的变化,例如:在烟芽夜蛾中,雄虫与已交配过的雌虫交配时会提供更大的精包和相同数量的精子,非精子组分的增加可提高雄虫的精子竞争力(LaMunyon and Huffman,2001)。但小菜蛾雄成虫与已交配过的雌成虫交配时,却只提供较小的精包。在黑腹果蝇中,雌成虫再交配时其产卵速率并不会得到再次提高,因此当雄成虫与已交配过的雌成虫交配时,雄成虫策略性地减少精液中ovulin蛋白的含量,但是保持性肽的含量不变以抑制雌成虫的再交配行为(Sirotetal.,2011)。因此,推测小菜蛾雄成虫精液蛋白在刺激雌虫排卵、提高产卵量和卵孵化率以及参与精子竞争等方面具有相对独立的调控机制,所以当雄成虫与不同交配状态雌成虫交配时可策略性地调整精液蛋白含量或组分的变化,以最小的代价获得最大的生殖利益。
交配能力测定结果显示,小菜蛾雌成虫在首次交配后12 h内的再交配率为83.7%,根据Torres-Vila等(2004)文章中对雌虫交配策略的界定标准,小菜蛾雌虫属于多夫制交配策略。此结果与Wang等(2005)的结果截然相反,作者在实验中只观测到19.3%的雌虫发生再次交配,因此认为小菜蛾雌虫采取一夫制交配策略。影响雌虫交配次数的因素有很多,如地理种群(Välimäkietal.,2006)、生态环境(McFarlaneetal.,2011)、雌虫个体生理状态(Syriatowicz and Brooks,2004;Frickeetal.,2010)和遗传基础(Harano and Miyatake,2005;Välimäki and Kaitala,2006;Price TARetal.,2011)等因素。比较小菜蛾G88品系与Wang等(2005)所用品系,发现除了饲养环境(温度、湿度和光照时间)相似外,幼虫食物、种群来源和实验室驯化时间均有很大差异,这些差异因素是否会导致小菜蛾雌虫交配能力的差异还有待进一步验证。小菜蛾雌成虫通过多次交配并没有获得产卵量和卵孵化率收益。相似的结果也见于其他多夫制昆虫,如苹果巢蛾Yponomeutapadella(Parkeretal.,2013)、梨小食心虫Grapholitamolesta(de Moraisetal.,2012)和蔬菜夜蛾Copitarsiadecolora(Callado-Galindoetal.,2013)。许多研究也表明雌虫多次交配不仅可获得产卵量和卵孵化率收益,通过接受不同雄虫的精子还可增加后代基因的多样性和减少基因不相容性的负作用(Arnqvist and Nilsson,2000;刘兴平等,2008)。小菜蛾雌虫多次交配策略是否可以获得基因利益,还有待进一步验证。
猜你喜欢
广西林业科学(2022年1期)2022-03-03
植物保护(2022年1期)2022-02-10
农业技术与装备(2022年11期)2022-01-01
甘肃农业科技(2021年6期)2021-07-25
环境昆虫学报(2020年5期)2020-11-11
上海农业科技(2020年5期)2020-10-24
科学导报(2019年35期)2019-09-03
蔬菜(2018年9期)2018-09-21
江苏农业科学(2014年3期)2014-07-16
