
MicroRNA通路中的关键因子Dicer-1和Argonaute-1对豌豆蚜免疫防御的影响
2022-02-08 02:04:08许珍珍蒋新怡石苏可吕志强
昆虫学报 2022年12期
许珍珍,蒋新怡,石苏可,吕志强
(西北农林科技大学植物保护学院,陕西杨凌 712100)
microRNA(miRNA)是一类非编码RNA,长约22 nt,在基因表达调控中发挥重要作用,对真核生物的生长发育至关重要(Bartel,2009;Jaubertetal.,2010;Pasquinelli,2012)。研究表明,miRNA及其通路中的组分与昆虫的先天性免疫有关(Sabinetal.,2009;Asgari,2013;Wangetal.,2021)。黑腹果蝇Drosophilamelanogaster的miR-958靶向Toll通路中的Toll和Dif基因,降低下游抗菌肽基因Drosomycin的表达(Lietal.,2017)。斯氏按蚊Anophelesstephensi体内let-7和miR-100可靶向球孢白僵菌Beauveriabassiana的sec2p和C6TF,降低其表达,抑制真菌毒力(Wangetal.,2021)。棉铃虫Helicoverpaarmigera在干扰Dicer-1和Ago-1后,更容易受到细菌的侵染(Baradaranetal.,2019)。
Dicer-1是miRNA产生过程中一个重要的内切酶,属于RNaseⅢ家族,它可将pre-miRNA剪切并转化为成熟的miRNA(Niuetal.,2017)。沙漠蝗Schistocercagregaria的Dicer-1包括1个DEAD解螺旋域(DEAD-like helicase domain,DEXDc)、1个C端解螺旋域(helicase,HELI)、1个Dicer二聚化结构域、1个PAZ结构域、2个RNase III结构域和1个dsRNA结合基序(Wynantetal.,2015)。Dicer-1对昆虫的许多生物学过程具有重要意义。抑制飞蝗Locustamigratoria的Dicer-1导致若虫到若虫以及若虫到成虫的蜕皮不能正常进行(Wangetal.,2013)。沉默棉铃虫的Dicer-1基因导致miRNA合成受到抑制,miRNA的表达降低,成虫发育出现畸形,死亡率升高(Rahimpouretal.,2019)。
Argonaute-1(Ago-1)蛋白是RNA诱导沉默复合物(RNA-inducing silencing complex,RISC)的核心成分,该复合物包含Ago和PIWI蛋白亚类。橘小实蝇Bactroceradorsalis的Ago-l包含DUF1785结构域、PAZ结构域和PIWI结构域(Yangetal.,2021)。Ago-1和Ago-2是Ago蛋白亚类的两个成员,它们在小RNA导向的RNA切割途径中有不同的作用。Ago-1是miRNA通路中RISC的主要成分,参与miRNA通路,而Ago-2主要与siRNA通路相关(Okamuraetal.,2004)。
Dicer-1和Ago-1是miRNA通路中的两个关键组分,昆虫体内Dicer-1和Ago-1的缺失会导致其miRNA表达异常(Maetal.,2017;Rahimpouretal.,2019;Yangetal.,2021)。与其他被广泛研究的模式昆虫相比,豌豆蚜Acyrthosiphonpisum具有相对有限的免疫系统(Gerardoetal.,2010),但其体内的miRNA合成及作用通路是完整的(International Aphid Genomics Consortium,2010)。而且,研究表明miRNA及其通路中相关因子与昆虫抵御病原物侵染的免疫防御反应有关(Baradaranetal.,2019;Maetal.,2020)。因此,本研究检测了豌豆蚜感染细菌和真菌后Dicer-1和Ago-1基因的转录水平,并分析了沉默Dicer-1和Ago-1对豌豆蚜体内细菌和真菌增殖以及豌豆蚜存活率的影响。本研究明确了miRNA通路中两个关键组分Dicer-1和Ago-1在豌豆蚜抵御病原物侵染的免疫防御反应中的作用。
1 材料与方法
1.1 供试虫源
本研究所用的豌豆蚜采自中国云南省,在实验室培养箱中饲养,实验种群由单头成蚜孤雌生殖产生,饲养条件如下:温度20±1℃、相对湿度70%±5%、光周期16L∶8D。取15头左右健康成蚜接于盆栽的蚕豆苗上,并用透明塑料罩将蚕豆苗罩住,塑料罩顶端用细的尼龙网纱布封口,约24 h后将蚕豆苗上成蚜全部移除,留下所产的若蚜,继续饲养直至其发育为无翅成蚜,饲养期间每天清洁蜜露和蜕掉的皮,并定期给蚕豆苗浇水。
1.2 细菌和真菌侵染
侵染所用的细菌为金黄色葡萄球菌Staphylococcusaureus和大肠杆菌Escherichiacoli,将菌液活化后,重新接于LB培养基中,37℃下240 r/min培养至OD600为0.8~1.0,离心,收集菌体,然后用无菌0.85% NaCl溶液洗涤3次,最后将金黄色葡萄球菌悬浮至终浓度为2×1011CFU/mL,大肠杆菌为1010CFU/mL;侵染所用真菌为球孢白僵菌,将其接于马铃薯葡萄糖培养基(potato dextrose agar,PDA)上,27℃光照培养箱中培养6~8 d,使其充分产孢,洗涤菌丝,并使菌丝与孢子分离,使用血球计数板计算孢子悬浮液浓度,最后加入0.05% Tween-20,使得孢子悬浮液终浓度为2×107孢子/mL,用于感染。
细菌侵染时将毛细玻璃管尖头末端截为2 mm左右的斜面,平头末端用封口膜密封,尖端蘸取菌液刺入豌豆蚜成蚜背部,大约3 s后将毛细玻璃管取出,对照组豌豆蚜成蚜用0.85% NaCl溶液;真菌侵染时吸取1 μL孢子液滴于豌豆蚜背部,对照组同样用0.85% NaCl溶液。侵染后将成蚜继续饲养于新鲜蚕豆苗上。在细菌侵染后6,12和24 h时和真菌侵染后1,2,3和4 d时,从对照组和处理组中分别取5头成蚜至1.5 mL RNase-free 离心管,加入200 μL TRIpure Isolation Reagent (Roche,Basel,瑞士),置于-80℃保存。侵染实验独立重复3次。
1.3 Dicer-1和Ago-1表达的qRT-PCR检测
将1.2节所得样品充分研磨使其完全破碎,利用TRIpure试剂(Roche,Basel,瑞士)提取豌豆蚜成蚜总RNA。按照First Strand cDNA Synthesis反转录试剂盒(Roche,Basel,瑞士)说明合成cDNA。以所得cDNA为模板,根据豌豆蚜Dicer-1(GenBank登录号:XM_001944279.4)和Ago-1(GenBank登录号:XM_003240572.3)的基因序列,在线设计目的基因和内参基因核糖体蛋白L7(ribosomal protein L7,Rpl7)基因(GenBank登录号:NM_001135898.1)的引物(表1)。将cDNA原液稀释至工作液(100 ng/μL),进行qRT-PCR反应,所有反应做3次技术重复。反应体系(20 μL):2×Fast qPCR Master Mix (Biosystems) 10 μL,cDNA工作液2 μL,上下游引物(10 μmol/L)各1 μL,ddH2O 6 μL。反应程序:95℃预变性10 min;95℃变性15 s,58℃退火15 s,72℃延伸20 s,40个循环。熔解曲线温度为65℃-95℃。
1.4 Dicer-1和Ago-1的RNAi
将1.2节所得样品充分研磨使其完全破碎,利用TRIpure试剂(Roche,Basel,瑞士)提取豌豆蚜成蚜总RNA。按照First Strand cDNA Synthesis反转录试剂盒(Roche,Basel,瑞士)说明合成cDNA。以所得cDNA为模板,基于Dicer-1和Ago-1序列,利用在线工具SnapDragon (http:∥www.flyrnai.org/cgi-bin/RNAi_find_primers.pl)设计合成dsRNA的引物(表1),通过PCR扩增干扰片段,然后使用天根公司胶回收试剂盒对得到的PCR产物进行回收,作为dsRNA合成的模板,按T7 Ribo MAXTMExpress RNAi System Kit试剂盒(Promega)说明,合成dsRNA。将dsRNA稀释至终浓度为10 μg/μL,分装后置于-80℃超低温冰箱保存。
参考张永栋等(2017)的方法,用CO2将豌豆蚜成蚜麻醉,在显微注射仪下进行dsDicer-1和dsAgo-1注射,注射浓度均为10 μg/μL,注射量为100 nL,对照组注射小鼠淋巴毒素A(Musmusculuslymphotoxin A,LTA)基因的dsLTA,注射浓度和剂量同处理组,引物序列见表1。待豌豆蚜苏醒后,将其接于新的蚕豆苗上继续饲养。在注射dsRNA后1-3 d,利用qRT-PCR检测(同1.3节)干扰效率。

表1 引物信息Table 1 Primer information
1.5 RNAi后细菌和真菌侵染豌豆蚜
收集1.4节注射dsDicer-1后48 h时和注射dsAgo-1后12 h时的豌豆蚜成蚜,进行金黄色葡萄球菌、大肠杆菌和球孢白僵菌侵染,方法同1.2节,以注射dsLTA的豌豆蚜成蚜为对照。细菌菌落形成单位(colony-forming unit,CFU)和真菌孢子数检测:在大肠杆菌侵染后12,24和36 h时,金黄色葡萄球菌侵染后24,36和48 h时,球孢白僵菌感染后6 d时,从每组取10~20头成蚜,放入无菌EP管中,每管1头,用75%乙醇对豌豆蚜表面进行消毒,用无菌的0.85% NaCl溶液清洗成蚜表面两次后吸出溶液,加入200 μL 0.85% NaCl溶液,手动研磨,使虫体完全被磨碎作为含菌原液,稀释至合适浓度后吸取10 μL均匀涂于LB固体培养基上,37℃培养箱中倒置培养后统计CFU/头;吸取50 μL真菌溶液均匀涂于PDA固体培养基上,27℃培养箱中培养后统计孢子数。
每组各选取20头成蚜于健康蚕豆苗上继续饲养,在侵染后1-8 d每天统计各组成蚜死亡数,计算存活率。实验重复3次。
1.6 数据分析
采用2-ΔΔCt法进行基因表达量的相对定量分析(Schmittgen and Livak,2008);使用Gragpad Prism 5.0软件作图,采用 Student氏t检验进行差异显著性分析,其中死亡率结果采用Log-rank(Mantel-Cox)检验分析。
2 结果
2.1 细菌和真菌侵染后豌豆蚜成蚜Dicer-1和Ago-1基因的表达变化
结果显示,与对照组(0.85% NaCl溶液处理)比较,在感染金黄色葡萄球菌后12 h时豌豆蚜Dicer-1的表达量显著上升(P<0.01)(图1:A),感染大肠杆菌后12和24 h时,Dicer-1的表达量略高(图1:B),在感染这两种细菌后12-24 h内,Dicer-1的表达量逐渐升高;在感染球孢白僵菌后1-3 d,Dicer-1的表达量均显著升高(P<0.05),2 d时的表达量为最高(图1:C)。在感染金黄色葡萄球菌后24 h时Ago-1的表达量显著高于对照组的(P<0.01),且在12-24 h逐渐升高(图1:D);感染大肠杆菌后12 h时,Ago-1的表达量为最高,与对照组存在显著差异(P<0.01)(图1:E);感染球孢白僵菌后1 d时Ago-1的表达量略低于对照组,2和4 d时的略高于对照组,3 d时的与对照组的相当(图1:F)。以上结果表明豌豆蚜Dicer-1和Ago-1的表达受到细菌及真菌侵染的诱导,暗示可能与豌豆蚜抵御细菌及真菌侵染有关。
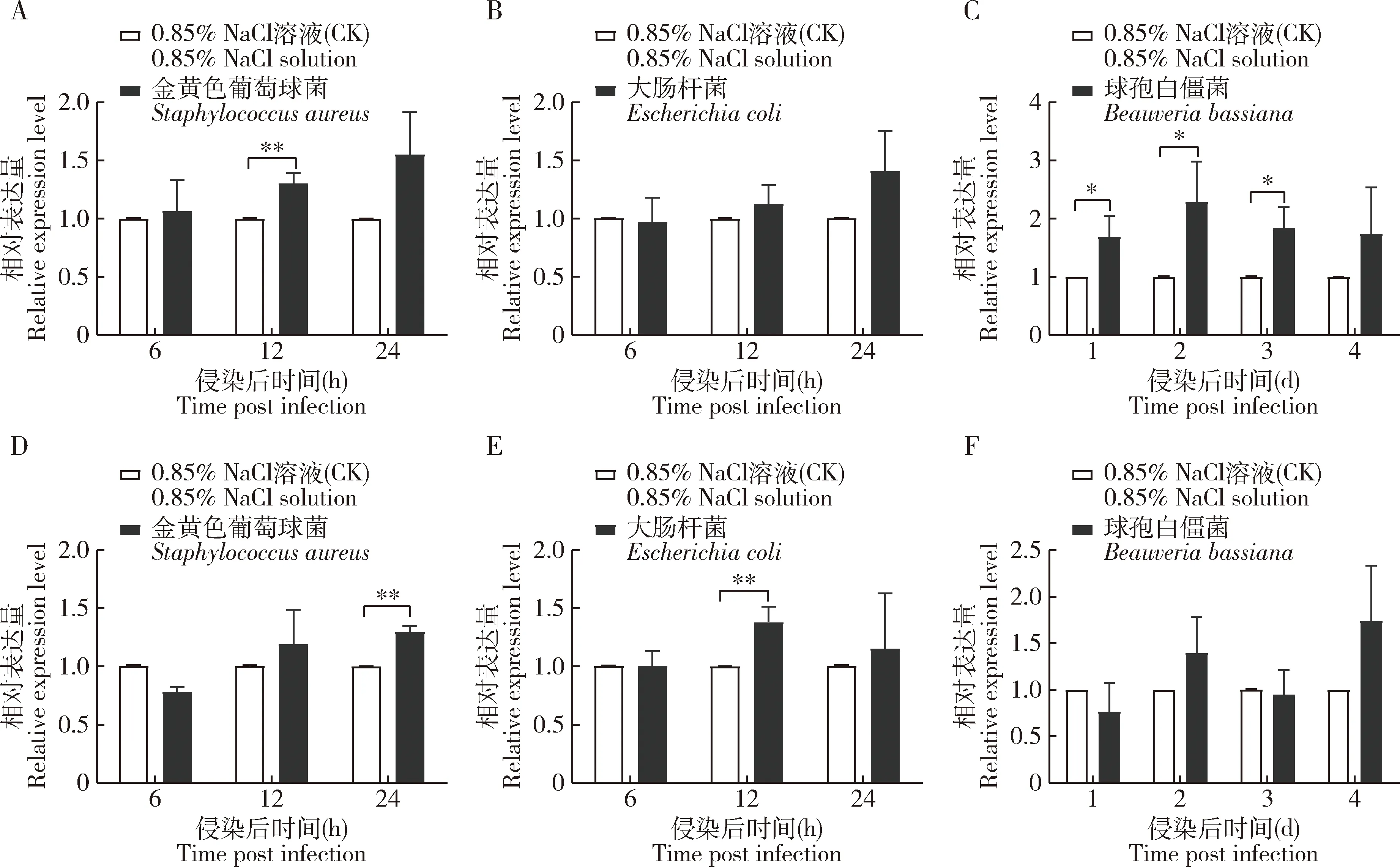
图1 细菌和真菌侵染后豌豆蚜成蚜Dicer-1(A,B,C)和Ago-1(D,E,F)的相对表达量Fig.1 Relative expression levels of Dicer-1 (A,B,C) and Ago-1 (D,E,F) in Acyrthosiphon pisum adults after bacterial and fungal infectionsA,D:金黄色葡萄球菌Staphylococcus aureus (2×1011 CFU/mL);B,E:大肠杆菌Escherichia coli (1010 CFU/mL);C,F:球孢白僵菌Beauveria bassiana (2×107 conidia/mL).CFU:菌落形成单位Colony-forming unit.图中数据为3次独立重复的平均值±标准误;柱上星号表示两组间差异显著(*P<0.05;**P<0.01;***P<0.001)(Student氏t检验)。Data in the figure are mean±SE of three independent experiments.Asterisks above bars indicate significant difference between two groups (*P<0.05;**P<0.01;***P<0.001)(Student’s t-test).下图同The same for the following figures.
2.2 Dicer-1和Ago-1的RNAi效率
结果显示,在注射dsDicer-1后1-3 d,豌豆蚜成蚜体内Dicer-1的表达量下调,且在注射后3 d时干扰效率最高,Dicer-1的表达量约为对照组(注射dsLTA)的50%,二者差异极显著(P<0.01)(图2:A)。在注射dsAgo-1后1 d时,豌豆蚜Ago-1基因表达量约为对照组的50%,两组间差异极显著(P<0.01);且在注射后2 d时仍存在一定的干扰效果,两组间Ago-1基因表达量差异极显著(P<0.01)(图2:B)。表明干扰导致目的基因表达量下降具有一定的持续性,可确保后续实验的进行。

图2 豌豆蚜成蚜中Dicer-1(A)和Ago-1(B)的RNA干扰效率Fig.2 RNA interference efficiency of RNAi-mediated knockdown of Dicer-1 (A) and Ago-1 (B) in Acyrthosiphon pisum adults
2.3 RNAi干扰Dicer-1和Ago-1后感染细菌或真菌豌豆蚜成蚜体内细菌和真菌的增殖
结果显示,干扰Dicer-1后感染金黄色葡萄球菌36 h后豌豆蚜体内的细菌菌落数(CFU/蚜虫)显著高于对照组(注射dsLTA)的(P<0.05)(图3:A);感染大肠杆菌后12和24 h时豌豆蚜体内细菌菌落数显著高于对照组的(P<0.05)(图3:B);感染球孢白僵菌后6 d时豌豆蚜体内的孢子数显著高于对照组的(P<0.05)(图3:C)。在干扰Ago-1后感染金黄色葡萄球菌后36 h时豌豆蚜体内细菌菌落数显著高于对照组的(P<0.05)(图3:D);感染大肠杆菌后24 h时豌豆蚜体内细菌菌落数显著高于对照组的(P<0.05)(图3:E);感染球孢白僵菌后6 d时豌豆蚜体内的孢子数显著高于对照组的(P<0.05)(图3:F)。

图3 RNAi干扰Dicer-1(A,B,C)和Ago-1(D,E,F)后细菌和真菌侵染对豌豆蚜成蚜体内细菌和真菌增殖的影响Fig.3 Effects of RNAi of Dicer-1 (A,B,C) and Ago-1 (D,E,F) followed by bacterial and fungal infections on the bacterial and fungal proliferation in Acyrthosiphon pisum adultsA,D:金黄色葡萄球菌Staphylococcus aureus (2×1011 CFU/mL);B,E:大肠杆菌Escherichia coli (1010 CFU/mL);C,F:球孢白僵菌Beauveria bassiana (2×107 conidia/mL).注射dsDicer-1 48 h和注射dsAgo-1 12 h后分别对豌豆蚜成蚜用1 μL金黄色葡萄球菌、大肠杆菌和球孢白僵菌感染。图4同。图中每个点表示1头蚜虫。A. pisum adults were infected with 1 μL of S. aureus,E. coli and B. bassiana,respectively,at 48 h after dsDicer-1 injection and 12 h after dsAgo-1 injection.The same for Fig.4.Each dot in the figure represents one aphid.
2.4 RNAi干扰Dicer-1和Ago-1后感染细菌或真菌的豌豆蚜的存活率
结果显示,干扰Dicer-1后豌豆蚜成蚜的存活率略低于对照组(注射dsLTA)的,但两者无显著差异(P>0.05);干扰Dicer-1后感染金黄色葡萄球菌,豌豆蚜成蚜的存活率略微低于对照组,二者也无显著差异(P>0.05)(图4:A)。干扰Dicer-1的豌豆蚜成蚜在感染大肠杆菌后2 d时,存活率约为60%,而对照组的约为80%;感染后4 d时,处理组存活率约为40%,而对照组的大于60%;在实验结束(7 d)时,处理组的仅有约5%存活,而对照组约有20%存活。整体生存分析表明,7 d时处理组豌豆蚜成蚜的存活率极显著低于对照组的(P<0.001)(图4:B)。干扰Dicer-1的豌豆蚜成蚜在感染球孢白僵菌后2 d时,存活率约为50%,对照组的约为70%;感染后4 d时,处理组存活率约为20%,而对照组的约为45%;感染后7 d时,处理组的仅有约10%存活,而对照组的约有25%存活。整体分析表明,7 d时处理组豌豆蚜成蚜的存活率显著低于对照组的(P<0.05)(图4:C)。
干扰Ago-1后,未感染的情况下,豌豆蚜成蚜约有42%的存活率,极显著低于对照组的(P<0.01);在受到金黄色葡萄球菌感染后,干扰Ago-1的豌豆蚜成蚜与对照组(注射dsLTA)豌豆蚜成蚜均在感染后5 d时全部死亡,但干扰Ago-1的豌豆蚜成蚜在感染后2 d时仅有约50%的个体存活,而对照组存活率约为75%,生存分析表明处理组与对照组之间存活率有极显著差异(P<0.001)(图4:D)。在干扰Ago-1后感染大肠杆菌,豌豆蚜成蚜的存活率略低于对照组的,但二者无显著差异(P>0.05)(图4:E)。在受到球孢白僵菌感染后7 d时,干扰Ago-1的豌豆蚜成蚜的存活率仅为40%,而对照组存活率高于70%;在7 d时,处理组豌豆蚜成蚜全部死亡,而对照组仍有约20%的个体存活,二者存在极显著差异(P<0.001)(图4:F)。

图4 RNAi干扰Dicer-1和Ago-1后细菌和真菌增侵染对豌豆蚜成蚜存活率的影响Fig.4 Effect of RNAi of Dicer-1 and Ago-1 followed by bacterial and fungal infections on the survival rates of Acyrthosiphon pisum adults通过Log-rank(Mantel-Cox)方法分析感染后7 d内的豌豆蚜成蚜存活率。星号表示侵染后7 d时两组间存活率差异显著(*P<0.05,**P<0.01;***P<0.001)(Student氏t检验)。Survival rates of the infected adults of A. pisum in 7 d were compared using Log-rank (Mantel-Cox) test.Asterisks indicate significant difference in the survival rate at 7 d post infection between two groups (*P<0.05,**P<0.01;***P<0.001)(Student’s t-test).
3 讨论
生产实践中,人们通常利用天敌和病原微生物对豌豆蚜进行生物防治(潘明真等,2022),而天敌和病原微生物会引发豌豆蚜的免疫防御反应。研究表明miRNA及其合成过程中的多种元件参与了昆虫抵御细菌、真菌以及病毒侵染的免疫防御过程。Baradaran等(2019)发现在感染苏云金芽孢杆菌Bacillusthuringiensis和马氏沙雷菌Serratiamarcescens后,棉铃虫体内Dicer-1和Ago-1的转录水平升高。南美白对虾Litopenaeusvannamei与昆虫同属节肢动物门,在感染陶拉综合征病毒(taura syndrome virus,TSV)后,其血细胞和鳃中Dicer-1基因的表达显著上升,淋巴器官中Ago-1基因的转录水平也升高(Yaoetal.,2010)。本研究得到类似结果,在感染细菌金黄色葡萄球菌和大肠杆菌以及真菌球孢白僵菌后,豌豆蚜体内Dicer-1和Ago-1基因的表达量升高(图1),暗示可能与豌豆蚜抵御细菌及真菌侵染有关。
在棉铃虫中,RNAi干扰Dicer-1使其更容易受到苏云金芽孢杆菌和马氏沙雷菌的侵染,细菌在体内增殖速度更快,并且有更高的死亡率(Baradaranetal.,2019)。本研究中,RNAi干扰Dicer-1和Ago-1后感染金黄色葡萄球菌和大肠杆菌,豌豆蚜体内细菌增殖速度显著升高(图3),存活率也下降(图4),尤其是干扰Dicer-1后感染大肠杆菌及干扰Ago-1后感染金黄色葡萄球菌,豌豆蚜的存活率显著低于对照组(图4),表明豌豆蚜Dicer-1和Ago-1参与其对细菌侵染的免疫反应,在一定程度上参与豌豆蚜抵御细菌侵染。
球孢白僵菌是一种广泛应用的生防真菌。本研究中,RNAi干扰Dicer-1和Ago-1后感染球孢白僵菌,豌豆蚜体内的孢子数均显著高于对照组(图3:C和F),存活率显著低于对照组(图4:C和F)。因此,我们认为Dicer-1和Ago-1参与抑制入侵真菌在豌豆蚜体内的增殖,对豌豆蚜抵御真菌侵染的免疫防御反应具有重要作用。昆虫miRNA可影响入侵真菌球孢白僵菌的毒力。在受到球孢白僵菌侵染后,斯氏按蚊Anophelesstephensi体内let-7和miR-100的表达上调,靶向真菌sec2p和C6TF,抑制真菌毒力,增强自身免疫(Wangetal.,2021)。因此,我们推测,豌豆蚜Dicer-1和Ago-1的沉默影响了其体内某些miRNA的生成,或阻止其生成RNA沉默诱导复合物,抑制其功能的发挥,而这些miRNA可能在豌豆蚜抵御真菌侵染中发挥重要作用。具体作用及调控机制还需进一步研究。
我们在干扰了Ago-1之后,发现豌豆蚜的存活率显著低于对照组(图4:D)。橘小实蝇Ago-1基因的下调导致7个表达量相对较高的miRNA差异表达,同时延缓了卵巢的发育,推测这7个差异表达的miRNA与橘小实蝇的卵巢发育有关(Yangetal.,2021)。果蝇miR-305与其寿命长短有关,果蝇体内过表达miR-305可抑制4种抗菌肽基因和类胰岛素肽基因的表达,并可能通过对这些基因的调控影响果蝇的衰老(Uedaetal.,2018)。我们推测,Ago-1蛋白作为昆虫miRNA通路中RISC的重要组成部分,可能通过影响在豌豆蚜的生长发育中发挥重要作用的miRNA的产生,从而影响豌豆蚜的存活。干扰Ago-1对豌豆蚜生存的缓慢影响(图4:D)也暗示了这一点。
RNAi是一种可持续的、环境友好的害虫防治方法,对预防昆虫抗药性具有重要意义。通过RNAi抑制羧酸酯酶(carboxylesterases,CarE)基因的表达可增加抗性棉蚜Aphisgossypii对有机磷杀虫剂氧化乐果的敏感性(Gongetal.,2014)。通过RNAi干扰Dicer-1和Ago-1导致棉蚜对单宁酸和棉酚有更高的敏感性,死亡率显著高于对照组(Maetal.,2017)。在本研究中,我们的结果也表明,通过RNAi干扰Dicer-1和Ago-1后,豌豆蚜在受到细菌和真菌侵染时出现更高的死亡率(图4),表明豌豆蚜体内miRNA通路中的两种重要组分Dicer-1和Ago-1,可以作为未来通过病原微生物防治豌豆蚜的潜在靶标。
总体来说,我们的研究发现Dicer-1和Ago-1参与豌豆蚜抵御细菌和真菌侵染的免疫防御反应,尤其在抵御真菌免疫防御反应中发挥重要作用,可作为病原物防治豌豆蚜的重要靶标。另外,由于Dicer-1和Ago-1在昆虫miRNA通路中的重要作用,我们推测豌豆蚜体内依赖Dicer-1和Ago-1的miRNA在其抵御病原物侵染的免疫反应中具有重要作用。这些发现加深了我们对豌豆蚜免疫防御系统的了解,为豌豆蚜的生物防治提供了理论基础。
猜你喜欢
环境昆虫学报(2023年5期)2023-11-09 12:31:52
中国生物防治学报(2021年3期)2021-08-11 02:15:38
湖北林业科技(2019年2期)2019-05-05 05:36:12
大灰狼(2018年5期)2018-06-20 14:49:32
中成药(2017年5期)2017-06-13 13:01:12
课外生活(小学1-3年级)(2017年5期)2017-06-10 10:38:03
上海农业学报(2016年2期)2016-10-27 00:49:41
故事作文·高年级(2016年9期)2016-10-25 08:50:53
广西林业科学(2016年4期)2016-03-16 05:44:50
故事作文·高年级(2015年1期)2015-09-07 08:21:57
