
养分元素在荔枝叶片—枝条—果实间的转移能力
2022-01-28 07:41苏雪霞白翠华周昌敏朱陆伟姚丽贤
中国南方果树 2022年1期
苏雪霞,白翠华,周昌敏,朱陆伟,姚丽贤
(华南农业大学资源环境学院,广州,510642)
果树不同部位的养分储备对新组织器官的生长有非常重要的作用[1]。Ela等[2]研究了N在核桃树中的迁移规律,发现在新芽生长过程中N的吸收和迁移同时发生,其中上一年储存在树体中迁移而提供的N约占总N量的54%。而大多数树种树体生长和果实发育所需的矿质营养直接来源于叶片养分的转移[3]。枇杷叶片N、P、K、Mn、Zn等元素与果实矿质元素含量显著相关[4]。冬枣结果枝叶片累积的矿质元素N、P、K、Ca、Mg、Na等在果实发育期作为果实的养分“源”提供了更多的养分[5]。荔枝叶片N、P、K养分含量在收获期内逐渐降至年生长周期内最低水平,在施肥后迅速恢复,在秋梢老熟期到花芽分化期达最大值,而后叶片营养逐渐由“库”转变为“源”,随着生殖器官和果实生长发育逐渐下降,至新的收获期又降低至年生长周期的最低水平[6-7]。
不少研究表明,不同元素在植物体内的移动能力不同。一般认为N、P、K、Mg等在植物中容易被再利用,在植株中移动性高,而Ca、Fe、Zn、Mn、Mo等在植物体内再利用率较低,相对不易移动。苹果树中N、B具有较高的移动性,叶片、枝条积累的N、B可被运输到新生器官利用[8-9],但施于叶片中的Ca则极少向果实转移[10]。在大多数作物中,N、P、K、Mg无论是在缺乏还是供应充足的情况下都容易从衰老组织转移到幼嫩组织,而Cu、Fe、Zn、Mn的移动能力则取决于叶片的供应水平,只有供应充足时植物才能从衰老组织中重新调动这些营养[11]。而且,即使同一营养元素,在不同植物体中的移动能力也有所差别。对B在6个树种中的移动性进行比较,发现B在杏仁、苹果、油桃等富含山梨醇的树种中移动性很强,叶面施用的B以山梨醇的形式向库(果实)运输,且输出量很高;而在山梨醇含量低的阿月浑子、开心果、胡桃等3个树种,叶面施B只能增加叶片中B的含量,对果实B含量影响不大[12]。对红富士苹果叶面喷施Mg肥,可显著提高了叶片和果实中的Mg含量[13];但葡萄叶面喷施Mg肥,虽然极显著提高了葡萄叶片Mg含量,但对果实Mg含量影响却不大[14]。
我国荔枝虽然栽培面积世界最大,总产量最高,但存在荔枝单产偏低,且年际波动幅度较大的问题[15-16]。曾有研究指出,荔枝N、K、B、Cu、Zn等养分元素的缺乏可能会限制荔枝着果和果实的发育,导致单产低和果实品质差[17]。国内外对荔枝树体养分的吸收、转移和累积已有报道。10年生妃子笑荔枝在末次梢老熟至开花初期,树体除继续累积Ca、Zn、B外基本不吸收其他养分,果实累积的N、K、Ca、Zn、S养分基本全部来自果实膨大期树体的吸收;而P、Mg、B、Mo则部分来自于第一次梢和第二次梢的养分转移[18]。对6年生荔枝树体不同部位N、P、K、S、Cl、Ca、Mg、Cu、Zn、Mn、B等养分含量进行分析,发现约75%的养分存在于叶片、嫩枝(直径<1 cm)和小树枝(直径1~3 cm)中,为树体开花结果做储备,而这些储备的养分在开花和树体生长过程中的使用将取决于它们的相对移动性[19]。目前,关于营养元素从荔枝叶片经枝条到果实的移动能力研究较少。因此,本文拟探讨比较不同营养元素在荔枝体内的运输移动能力,可进一步丰富养分元素在植物体内的运输利用机制,为荔枝不同养分元素的差异化管理和施用提供参考。
1 材料与方法
1.1 荔枝品种
2019年6月下旬至7月初分别在广东省广州市从化区、增城区,汕尾市及广西灵山县采集了井岗红糯、仙进奉、凤山红灯笼、红绣球、糯桂、岭丰糯、美园糯、庙种糯、观音绿、红脆糯和糯米糍等11个品种的14份带叶片果穗样本。其中,井岗红糯、红灯笼、仙进奉各有2份样本,分别采自2个果园,每个样本3个果穗,重复3次,所有样本用于测试各种养分含量备用。
1.2 样本处理与测试
每份带叶片果穗分别采集叶片、枝条(指叶片与果实之间的枝条)、果柄、果蒂、果皮、种柄、果肉和种子8个部位的样本。除果肉外,所有样本用自来水清洗,再用去离子水冲洗,然后所有样本杀青、烘干和制样,用于测定N、P、K、Ca、Mg、Fe、Mn、Cu、Zn、B和Mo含量。N含量用H2SO4-H2O2消解-凯氏定氮仪测定,P、K、Ca、Mg、Fe、Mn、Cu、Zn含量用HNO3-HClO4消解-ICP-OES测定,B、Mo含量用HNO3-HClO4消解-ICP-MS测定。样本所有测定项目均用标准物质GBW07603控制测试质量。
1.3 数据分析
数据用Excel整理,用SPSS 20.0进行方差分析及Tukey多重比较。养分元素从叶片到不同部位的转移系数=不同部位养分含量/叶片养分含量,养分元素从叶片—枝条—果实的逐级转移系数=后一部位的养分含量/前一部位的养分含量。
2 结果与分析
2.1 不同部位养分含量
本研究中,由于采样包括新审定品种,种植面积较小,部分样本难以作为当地代表性果穗样本,因此目前不能对不同产地不同品种间各种养分的累积和转移进行比较。从表1可以看出,将全部品种荔枝叶片、枝条和果实不同部位养分含量进行比较,则N含量在叶片、种柄中较高,在果柄中次之,在果皮、种子、果蒂、枝条和果肉中较低。P在果柄中含量最高,其他部位含量从高到低依次是种柄、枝条、果蒂、果肉、种子和果皮,叶片中最低。K在果柄、果皮、果肉和果蒂中含量较高,枝条和种柄中次之,叶片和种子中较低。叶片和果柄中Ca含量较高,枝条、果蒂、果皮和种柄中次之,种子和果肉中Ca含量显著低于其他部位。Mg在叶片中含量显著高于其他部位,果柄、果皮、种子、种柄和果蒂中次之,在枝条和果肉中含量较低。Fe在叶片、种柄中含量远高于其他部位,果柄、果蒂和枝条中含量次之,在种子、果皮和果肉中含量较低。Mn在叶片中含量最高,其他部位含量从高到低依次是果蒂、果皮、果柄、种柄、枝条、种子和果肉,且在果肉中含量远低于其他部位。Cu在各个部位中含量较为均衡,其中在种子中含量最高,枝条中含量最低。对于Zn,在种柄、果柄、果蒂和种子中含量较高,叶片、果皮和枝条中含量次之,果肉中含量最低。B在种柄、果皮、叶片和果柄中含量较高,种子、果蒂、枝条中含量次之,在果肉中含量最低。Mo是含量最低的养分元素,除了果蒂外,它在种柄中含量显著高于其他部位,而在其他不同部位中含量差异不显著。由此可见,多数养分元素如N、Ca、Mg、Fe和Mn在叶片中含量最高;而P和K相反,在叶片中含量较低,在果柄中最高;部分元素如Zn、B和Mo在种柄中含量最高,在果肉中含量最低。
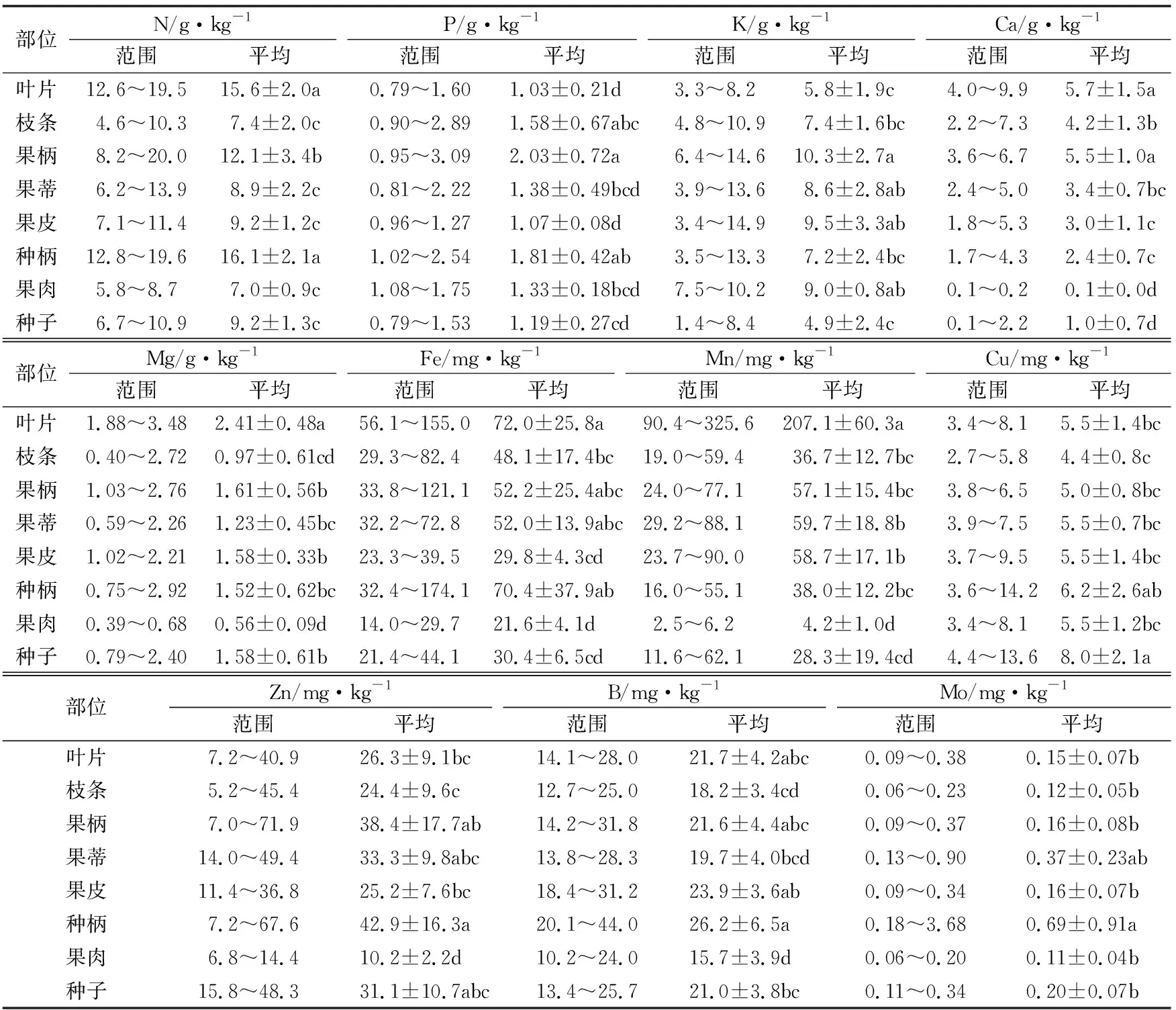
表1 供试11个荔枝品种叶片、枝条和果实不同部位的养分含量
2.2 养分从叶片向枝条和果实不同部位的转移
养分从叶片经枝条到果实的运输,体现了养分从源到库累积能力的强弱。从图1可以看出,将不同部位中养分含量与叶片中含量进行比较,则Mo从源(叶片)到库(果实)的转移能力最强,从叶片至各个部位的平均转移系数高达1.97;主要体现在向种柄移动的能力最强,转移系数达5.60,果蒂(2.79)、种子(1.48)、果柄(1.21)、果皮(1.13)次之,枝条(0.86)和果肉(0.74)较弱;说明Mo具有优先向繁殖器官转移累积的特点。K、P、Zn、Cu和B从叶片运输至各个部位的能力也较强,不同部位平均转移系数分别为1.54、1.47、1.22、1.13和0.98。其中K和P均表现为从叶片至果柄的运输能力最强,转移系数分别为1.95和1.98;Zn和B均是从叶片运输至种柄的移动能力最强,转移系数分别为1.74和1.24;Cu则表现为叶片运输至种子的移动能力最强,转移系数为1.60。Ca、Mg、Fe和N从叶片到枝条及果实不同部位的整体移动能力较低,平均转移系数分别为0.52、0.56、0.64、0.64;尤其从叶片至果肉的移动性最差,Ca、Mg、Fe和N移动系数分别为0.02、0.24、0.32和0.46。与其他养分元素相比,Mn的移动性最弱,平均转移系数仅为0.20,从叶片向各个部位的移动性都较差;尤其向果肉的移动能力最差,转移系数低至0.02。
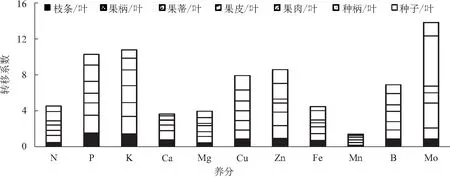
图1 不同养分元素从荔枝叶片转移至枝条和果实不同部位的转移系数比较
从图2可以看出,养分从叶片沿枝条逐级运输至果实不同部位的转移情况,K在不同部位运输的平均转移系数最高,达到1.17;而Cu、Mo和P的平均转移系数次之,平均转移系数分别为1.07、1.05、1.02;随后为Mg、B、Zn、N、Fe和Mn,平均转移系数分别为0.93、0.90、0.89、0.82、0.75、0.69;Ca的平均转移系数最低,仅为0.59。从养分在不同部位间转移系数的变异系数来看,则Mn的变异系数最大,达到86.0%;Ca次之,为79.8%,Mo、Mg、N、Zn、Fe、P、B、K的变异系数分别为75.1%、62.3%、52.3%、48.7%、38.8%、33.9%、26.8%和24.4%;Cu的变异系数最低,仅为15.5%。说明K从叶片至果肉的逐级运输中,平均移动能力相对最强,而且在不同部位间的移动能力较为接近,即K在不同部位间的转运均顺利流畅。Cu、P、B与K类似,不但移动能力强,而且在不同部位运输的能力相当。虽然Mo的平均移动系数较高,但其变异系数大,从种柄到果肉的移动成为Mo从叶片至果实运输的限制部位。Mg、Zn的移动能力整体也较高,但Mg和Zn从种柄及果皮到果肉的移动性较差。Fe和Mn的平均移动能力中等,但Fe从种柄至果肉的移动能力较差;Mn从果皮到果肉的移动性极差,转移系数仅为0.07,从种柄到果肉和从叶片到枝条的移动能力也极弱,移动系数分别为0.12和0.18。Ca在不同部位间的移动能力均较差;尤以从果皮到果肉的移动性最差,转移系数仅为0.05;从种柄到果肉的移动性也极差,转移系数为0.06。由此可见,Ca、Mn、Mg、Mo、Zn营养从果皮或种柄到果肉的移动性低,尤其是Ca和Mn极难从果皮或种柄移动到果肉,可能成为荔枝果实发育阶段的营养限制因素。
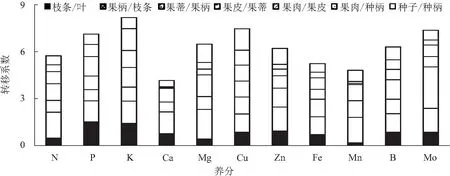
图2 不同养分元素从荔枝叶片—枝条—果实的逐级转移系数比较
3 结论与讨论
3.1 养分元素移动运输障碍导致的植物生理性病害
本研究显示,K、Cu、P、B在不同部位的逐级移动能力很强,Mo、Mg、Zn整体移动性也较强,Fe和Mn移动能力中等,而Ca的逐级移动能力均较差。Ca对维持细胞壁和细胞膜的结构和功能具有重要作用,由于运输障碍造成的缺Ca会导致膜结构破碎,引发一系列果实异常现象[20-21]。如苹果的苦痘病[22]、鸭梨的黑心病[23]、杧果的软鼻病[24]等。而且,通常认为果实缺Ca更多情况下是由于Ca在植物体内没有被运输分配到果实,而与土壤Ca供应水平关系不大[25]。Casero等[26]也发现苹果叶片与果实中的Ca含量没有相关性。荔枝缺Ca则会加剧荔枝裂果[27]。本课题组近年来调查发现华南多起荔枝龙眼果实膨大期果实发育异常现象,如小果期和中果期外果皮出现黑褐色斑点,内果皮黑褐色斑块更为明显,严重者果皮开裂,果核坏死;大果期外果皮皱缩凸起,果实畸形变小,内果皮有黑褐色或淡绿色向果肉侵入的痂状物等,初步判断主要是不良气候导致的养分运输障碍造成Ca或Ca和B的不足引发的生理性病害[28]。
同时,本研究表明,Mn在叶片—枝条—果实间的运输能力中等,但从果皮或种柄向果肉的运输能力极低。Mn是叶绿体的组成部分和植物体内多种酶的活化剂,直接参与植物的光合作用,并参与调节植物体内的氧化还原状况[29]。目前国内外缺乏荔枝缺Mn的报道,但在其他作物的研究表明,缺Mn会影响植物的生长、果实产量和品质。缺Mn导致的柑桔紫血焦病[30],甜瓜缺Mn会抑制生长发育,降低产量和质量[31],枇杷缺Mn显著影响果实品质[4],甜柿缺Mn会降低果实硬度和维生素C含量[32]。荔枝树体Mn的移动障碍可能对果实生长发育的影响有待进一步的研究。
3.2 提高荔枝叶片养分累积及向枝条—果实转移运输的措施
提高养分元素在荔枝叶片的累积,并适时促进它们向果实转移累积,是荔枝果实正常生长发育的重要物质保障。K、Cu、P、B、Mo、Mg、Zn等在荔枝树体移动能力较强,可根据荔枝不同生长发育期的养分需求,在土壤施入足量K、P、Mg肥;对于微量元素Cu、B、Mo、Zn,由于用量低且土壤吸附能力强[33-34],可按照不同生育期需要叶面喷施补充。
由于Ca从荔枝叶片至果实有较大的运输障碍,常用的施Ca措施如叶面喷施、树干注射、土壤施肥等难以有效补充果实Ca含量,可以在果实生长期直接施于果实上。植物成熟果实的角质层对果面Ca进入果肉组织具有阻碍作用,在幼果期对果实喷施Ca的效果较好[35]。将Ca与吸收促进剂共同喷施,则补钙效果更好。对于苹果,果面施Ca适宜时期为幼果形成1个月内,且吲哚乙酸(IAA)、赤霉素 (GA)、萘乙酸(NAA)均可促进果面Ca运往果实内[10]。另外,Ca在植物体内的运输主要通过蒸腾拉力实现。杏果实累积的Ca有一半以上是由于果实蒸腾作用驱动。通过夏季修剪或灌溉等促进果实蒸腾作用,可改善果实Ca营养状况[36]。因此,在荔枝果实发育阶段,适当修剪弱枝和荫蔽枝,可能会提高果实蒸腾作用而促进Ca向果实运输,但具体效果有待进一步研究。
虽然华南荔枝园土壤有效锰含量较丰富[37],但Mn在荔枝树体内的移动能力很低。目前缺乏荔枝果实Mn营养的深入研究,但其他作物果实中Mn营养及施肥措施已有不少报道。叶面喷施适宜浓度的Mn能提高番茄果实番茄红素、可溶性糖、维生素C含量并且可以改善糖酸比[38]。猕猴桃盛花期和盛花后3周各喷洒1次硫酸锰可提高果实Mn含量[39];采前喷施Ca、Mn以及Ca加Mn均有助于促进桃果实着色,并提高其耐贮性[40]。结合本研究结果,可考虑在荔枝果实发育期同时补充Ca和Mn,从而提高喷施效率。整体而言,荔枝果实发育阶段如何叶面补充Ca、Cu、B、Mo、Zn和Mn,有待进一步研究。
根据养分从叶片向不同部位的移动性,Mo从源(叶片)到库(果实)的转移能力最强,表现出优先向繁殖器官转移累积的特点。K、P、Zn、Cu和B的移动能力也较强,Ca、Mg、Fe和N则较差,而Mn的移动性最差。对养分从叶片至种子的逐级转移能力进行比较,则K、Cu、P、B在不同部位的逐级移动能力强;Mo、Mg、Zn和N整体移动性也较强,Fe和Mn逐级移动能力中等,Ca的逐级移动能力较差,尤其难以从果皮及种柄运输至果肉。综合而言,Ca、Mn、Mg、Mo、Zn营养从果皮或种柄到果肉的移动性低,尤其是Ca和Mn极难从果皮或种柄移动到果肉,可能成为荔枝果实发育的营养限制因子。
猜你喜欢
阅读(快乐英语中年级)(2019年4期)2019-09-10
语文周报·教研版(2019年9期)2019-03-20
移动通信(2017年19期)2017-11-14
电子技术与软件工程(2017年1期)2017-03-06
红领巾·探索(2016年12期)2017-01-04
中华手工(2016年4期)2016-04-20
老区建设(2015年18期)2015-10-26
伴侣(2015年9期)2015-09-09
软件导刊(2015年6期)2015-06-24
中国民族民间医药·下半月(2014年4期)2014-09-26
