
单瓣与重瓣垂丝海棠花器官形态发育比较观察
2022-01-26 03:04杨立梅
西北植物学报 2021年12期
魏 景,彭 冶,杨立梅
(南京林业大学 生物与环境学院,南京 210037)
垂丝海棠(MalushallianaKoehne)为蔷薇科(Rosaeeae)苹果亚科(Maloideae)苹果属 (Malus)植物,是中国蔷薇科特有的木本观赏植物之一[1]。据《中国植物志》记载,垂丝海棠产于江苏、浙江、安徽、陕西、四川、云南,多生于海拔50~1 200 m的山坡丛林中或山溪边。因花梗细长下垂,花色鲜艳,小花簇拥于伞房花序,垂丝海棠具有较高的观赏价值,是重要的园艺品种及种质资源,有重瓣、白花等变种[2]。1974年,《中国植物志》将其重瓣变种名称作为垂丝海棠的异名;1985年,《中国树木志》将其重瓣变种记载为重瓣垂丝海棠(Malushallianavar.parkmaniiRehd.);至1996年,《中国农业百科全书》将重瓣垂丝海棠作为栽培品种[3]。重瓣垂丝海棠花为半重瓣(8~14瓣),其余特点与垂丝海棠相近[4]。
花瓣作为观赏植物中最具观赏价值的结构, 其数量和形状的变化对花型的发展与进化有着重要意义, 是决定观花植物观赏价值的重要因素之一[5]。相比单瓣品种,重瓣垂丝海棠因其重瓣的花朵具有更高的观赏价值,且花重瓣属于观赏树种的创新性状[6]。迄今为止,国内外对垂丝海棠的形态学,胁迫生理学[7]、生殖生物学研究逐渐丰富[8-9],但对重瓣垂丝海棠的研究较少,对其重瓣性状的研究更为匮乏。深入研究其花器官分化过程与花形态变异是进一步探索重瓣性状的基础。
本研究主要观察了重瓣垂丝海棠花形态及花器官分化过程,以积累有关重瓣垂丝海棠花器官研究的资料,并为进一步研究垂丝海棠重瓣花形成原因和分子机制奠定形态学基础。
1 材料和方法
1.1 实验材料
实验材料采自南京林业大学校园内树龄大于40年且生长健康的单、重瓣垂丝海棠。
1.2 实验方法
1.2.1 花器官分化观察于7月底垂丝海棠花芽分化期,每隔3日分别从单瓣垂丝海棠和重瓣垂丝海棠取20个花芽固定保存于50% FAA固定液中,于Nikon SMZ18体式镜下解剖观察,直至花芽内顶花发育到萼片原基出现开始拍照记录直至花芽内顶花雌蕊原基分化结束。
将各时期解剖后的样品进行脱水干燥并置于EMITECH K850 临界点干燥仪中用 CO2临界法干燥,将干燥后的样品粘于样品台上放置在HITACHI E-1010离子溅射仪中喷金镀膜,最后用 OXFORD Inca X-act 能谱仪观察拍照单瓣与重瓣垂丝海棠萼片原基分化期,花瓣原基分化期,雄蕊原基分化期以及雌蕊原基分化期4个时期。
1.2.2 花器官形态观察于垂丝海棠3月份花期,对重瓣垂丝海棠盛开花朵的花器官进行观察并记录异常的花器官形态。另外,随机采取重瓣垂丝海棠不同花序中的50朵大花蕾(气球状)于体视显微镜下进行解剖观察,并对其萼片、花瓣、雄蕊、花柱、瓣化中的雄蕊以及异常花柱数目进行统计(苹果属植物的雌蕊多为3~5心皮复雌蕊,下位子房,花柱分离,因此,在统计心皮数时,以花柱数目代替心皮数目)。
1.2.3 数据处理原始数据经过Excel(2016版)初步分析,再应用R语言(4.0.3版)进行相关性分析。
2 结果与分析
2.1 花器官分化
单瓣垂丝海棠与重瓣垂丝海棠的花芽分化时期大致可分为花芽分化前期,花序原基分化期和花器官分化期。其中花器官分化期又可分为萼片原基分化期,花瓣原基分化期,雄蕊原基分化期以及雌蕊原基分化期4个时期[4],各轮花器官按照向心的顺序依次分化发育。其中,单、重瓣垂丝海棠在花序原基分化过程中均为顶花最早分化,周围3~6朵小花分化阶段一致,均较晚。
从六月上旬开始,单、重瓣垂丝海棠均进入花芽分化前期,直至十月中旬雌蕊分化完成,整个花芽分化期持续4个月。于八月上旬,单、重瓣垂丝海棠进入萼片原基分化期,小花原基上着生出5个突起,三大两小,为萼片原基(图1,A;图2,A)。九月中上旬垂丝海棠进入花瓣原基分化期,在单瓣垂丝海棠花芽内的顶花中,5枚圆形凸起的原基均起生于外轮萼片交汇处的内部,为花瓣原基(图1,B);而重瓣垂丝海棠每个花芽中的顶花内第一轮起生的5至7枚花瓣原基在萼片内部不均匀分布,在第一轮花瓣原基内部同时或相错极短时间分化出7至10个突起为第二轮花瓣原基(图2,B)。与叶原基发育模式相同,萼片原基与花瓣原基是由小花原基表面的几层细胞分裂形成最初的圆形凸起,接着向长、宽、厚三个方向生长,但各方向生长速率不同且厚度生长开始与停止均较早,使其进行扁平生长[10]。九月下旬,花瓣原基内部分化出2至3轮等大的圆形突起为雄蕊原基,单、重瓣垂丝海棠进入雄蕊原基分化期(图1,C;图2,C)。直至十月末,雌蕊原基在杯型花托最底部分化发育(图1,D;图2,D)。其中,雄蕊原基与雌蕊原基起初发育模式相同,其宽度生长与厚度生长停止较早,雌雄蕊均进行伸长生长。
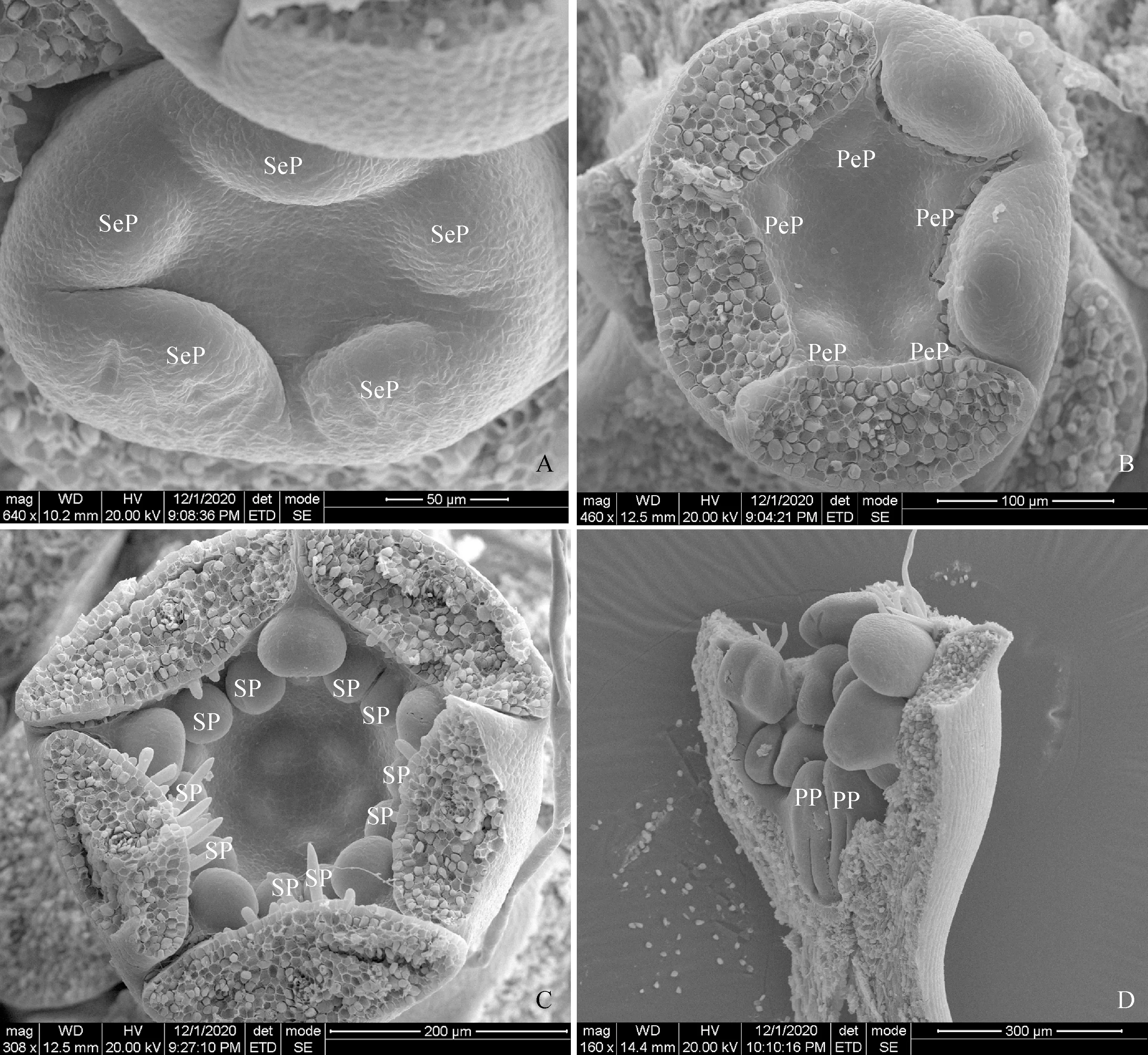
A.萼片原基(SeP)分化期;B.花瓣原基(PeP)分化期;C.雄蕊原基(SP)分化期;D.雌蕊原基(PP)分化期图1 单瓣垂丝海棠花器官分化A. Sepal primordium (SeP) differentiation stage; B. Petal primordium (PeP) differentiation stage; C. Stamen primordium (SP) differentiation stage; D. Pistil primordium (PP) differentiation stageFig.1 Differentiation of floral organs in Malus halliana
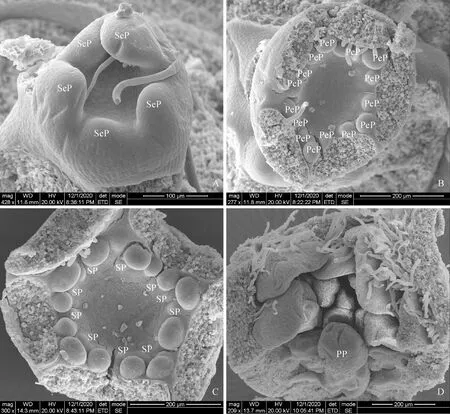
A.萼片原基(SeP)分化期;B.花瓣原基(PeP)分化期;C.雄蕊原基(SP)分化期;D.雌蕊原基(PP)分化期图2 重瓣垂丝海棠花器官分化A. Sepal primordium (SeP) differentiation stage; B. Petal primordium (PeP) differentiation stage; C. Stamen primordium (SP) differentiation stage; D. Pistil primordium (PP) differentiation stageFig.2 Differentiation of floral organs in M. halliana var. parkmanii
总体而言,萼片原基出现到花瓣原基出现间隔30日左右;雄蕊原基出现到雌蕊原基出现同样间隔30日左右;而花瓣原基分化期与雄蕊原基分化期的间隔较短,仅相隔8日左右。
2.2 花器官形态多样性
重瓣垂丝海棠的花包含单轮排列的萼片(5枚), 2至多轮花瓣(多为11~13枚),25~35 枚雄蕊,花柱0~5枚不等[11]。通过对南京林业大学重瓣垂丝海棠盛开花的系统观察发现,相比单瓣垂丝海棠,重瓣垂丝海棠不仅在其花瓣数目上存在巨大差异,在少数萼片中也存在数目与形态的异常(图3,B~D),雄蕊瓣化较为常见(图3,F~H),花药出现畸形发育(图3,J~L),花柱数目0~5枚不等且存在形态异常(图3,N~P)。其中,图3A、E、I、M为单、重瓣垂丝海棠正常形态的各轮花器官。
而对50朵重瓣垂丝海棠大蕾期花蕾的解剖发现,所解剖的重瓣垂丝海棠大蕾期的萼片数目均为5,其中3枚较大的萼片后期生长速度缓慢,2枚较小的萼片生长速度增加,5枚萼片随时间推移生长至等大。在大蕾期的花蕾与盛开的花中,雄蕊瓣化的情况较为常见,不同之处在于大蕾期中的雄蕊瓣化多数处于初期(仍可见未瓣化的花药部分),而盛开的花中的雄蕊瓣化多数处于末期(花药已完全瓣化为花瓣);在大蕾期的雄蕊群中也发现较多形态异常,如部分雄蕊的花粉囊呈现萎缩状态。除此之外,在重瓣垂丝海棠中还观察到萼片瓣化存在两种形式,一种是从萼片的一侧横向逐渐演变为花瓣(图3,C),另一种则是从顶端开始纵向演变为花瓣(图3,D);同样,雄蕊瓣化在重瓣垂丝海棠中也出现两种形式,一种是从花药的一侧出现淡红色(图3,F),逐渐变化为薄片状花瓣,另一种是从花药顶端开始变化(图3,H)。在大蕾期花柱还未完全伸长生长时,观察到部分花柱分出细枝,在花柱的中部或中下部斜生出另一支短小的花柱。正常雌蕊花柱为红色,雄蕊花丝为白色,而在重瓣垂丝海棠的雄蕊群与雌蕊群间的过渡区出现红色花丝的花药,其形态与空间位置均处于雄蕊群与雌蕊群之间(图3,J)。总体而言,重瓣垂丝海棠的花部结构从外至内由一轮萼片、2轮分布不均匀的花瓣、1轮直立伸长的雄蕊、1轮弯曲且未完全伸长的雄蕊、1轮花柱组成。
2.3 花器官数目相关性
统计重瓣垂丝海棠的萼片、花瓣、雄蕊、花柱、瓣化中的雄蕊以及异常形态的花柱数目,并利用R语言对以上6个指标进行相关性分析。图4为重瓣垂丝海棠各轮花器官数目间的相关系数矩阵图,其主对角线为各个指标的直方图;主对角线下方为各指标间的相关性散点图;主对角线上方为各指标间的 Spearman相关系数,其相关性系数越大,则相关性越强。该图结果显示,在重瓣垂丝海棠大蕾期的各轮花器官中,萼片数目与其余各轮花器官数目之间并无相关性,始终保持在5的水平上;而花瓣数目与雄蕊以及瓣化中的雄蕊数目相关性系数分别为0.54、0.43,具明显的正相关性,即雄蕊与瓣化中雄蕊数目越多,其花瓣的数目也相应较多。花柱数目与异常的花柱数目之间的相关性系数为0.48,也具有一定的正相关性,其余花器官之间的相关性并不显著。
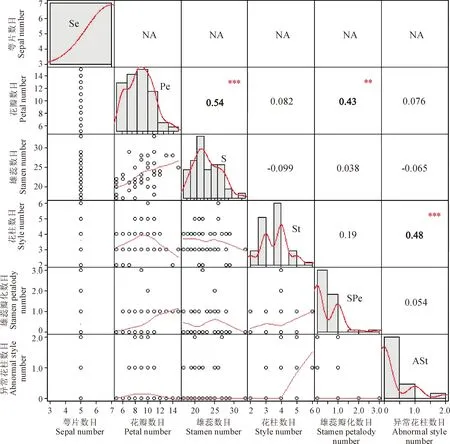
Se. 萼片数目;Pe. 花瓣数目;S. 雄蕊数目;St. 花柱数目;SPe. 雄蕊瓣化数目;ASt. 异常花柱数目;主对角线为各指标的直方图;主对角线左下方为各指标间的相关性散点图;主对角线右上方为各指标间的 Spearman相关系数其相关性系数越大,则相关性越强。**代表有显著差异(P<0.01), ***代表有极显著差异(P<0.001)图4 重瓣垂丝海棠各轮花器官数目间相关性分析Se. Sepal number; Pe. Petal number; S. Stamen number; St. Style number; SPe. Stamen Petalody number; ASt. Abnormal Style number; The main diagonal is the histogram of each indicator; the lower left of the main diagonal is the scatter plot of the correlation between the indicators; the upper right of the main diagonal is the Spearman coefficient between the indicators. The larger the correlation coefficient, the stronger the correlation. ** indicate significant difference (P<0.001); *** indicate extremely significant difference (P<0.001)Fig.4 Correlation analysis of the number of floral organs in each round of M. halliana var. parkmanii
3 讨 论
3.1 单、重瓣垂丝海棠花器官发育比较
观赏海棠不仅是苹果的优良砧木,也是景观园林中重要的风景线[12],前人对垂丝海棠及其变种重瓣垂丝海棠的形态结构学与生殖生物学研究已较为透彻[4,11],但对重瓣垂丝海棠的花器官特点未有深入研究。花起生于花分生组织,由同心轮生的花器官原基分化发育为外围的花被以及中心的生殖器官,花被分为第一轮的萼片与第二轮的花瓣,雄蕊在花瓣内部的第三轮发育,心皮在第四轮发育[13],单瓣垂丝海棠与重瓣垂丝海棠均遵循这个规律。
据文献记载,垂丝海棠为伞房花序[14],开花顺序应由外至内。本研究通过对单、重瓣垂丝海棠的花芽分化期、大蕾期花朵与盛开的花朵系统观察,发现其每个花序中大多包含4~7枚小花且顶花最先分化并绽放,这与传统的描述不同。就花芽分化过程而言,除在花瓣原基分化期时单、重瓣垂丝海棠的花瓣数量与位置存在差异,其余阶段特点大致相同。其中单瓣垂丝海棠在花瓣原基分化时,两片萼片之间分化出5个均匀分布的花瓣原基;而重瓣垂丝海棠在花瓣原基分化期中,萼片原基内部分化出两轮花瓣原基,第一轮的5~6枚花瓣原基大小相同,但位置分布不均匀,第二轮花瓣原基相比于第一轮花瓣原基体积相近或较小,排列在第一轮花瓣原基的内下方。与蔷薇属植物相似,单瓣垂丝海棠萼片、花瓣等大对称,且数目稳定,而重瓣垂丝海棠的花瓣位置分布不规则,数目也无规律可循[15]。
花瓣原基分化期后数天,在花瓣原基的内侧陆续分化出雄蕊原基,雄蕊原基与花瓣原基不仅在时间距离上较短,在空间上的距离也更为相近。在重瓣垂丝海棠的花蕾中,雄蕊与花瓣的关系则更为密切,可观察到由雄蕊到花瓣变异的不同阶段。与罗敏蓉对天葵花花器官发生的研究结果相似,垂丝海棠的花瓣原基与萼片原基形状差异较大,且存在较长的发生时间间隔;但花瓣原基与雄蕊原基形状相似,且相隔较短时间发生,该现象也暗示了重瓣垂丝海棠的花瓣雄蕊起源[16]。
3.2 垂丝海棠重瓣花成花原因推测
观赏植物重瓣的起源大致包括积累起源、苞片起源、雌雄蕊起源、台阁起源、重复起源和花序起源6种方式[17]。在同一种植物中也可能同时存在多种起源方式,如朱高浦在山茶花的重瓣化研究中发现山茶花同时存在积累起源,苞片起源,雌雄蕊起源[18]。其中,积累起源学说认为原种的单瓣花花瓣数量一般是固定的, 起初花瓣的变化数目较小,后来在自然选择或人工选择的作用下, 经过若干代后使花瓣数目逐渐增多。苞片起源与雌雄蕊起源均由于萼片与雌雄蕊在发育过程中逐渐演变为花瓣。
在重瓣垂丝海棠盛开的花的观察中,萼片与雄蕊在数量以及形态上出现差异,即萼片数目增多或逐渐演变出花瓣特征,雄蕊的瓣化在大蕾期中也存在。其中,雄蕊分为两轮相错排列在花瓣的内侧,雄蕊瓣化的情况通常仅出现在外轮雄蕊中,属于雄蕊的向心式瓣化。因此,结合重瓣垂丝海棠花芽分化阶段的形态学特征,可推断重瓣垂丝海棠的重瓣起源包含苞片起源与雄蕊起源。
雌雄蕊起源是由雄蕊或雌蕊变瓣,通过雌雄蕊数目减少以增加花瓣的数目[19],但从50朵大花蕾解剖的数据分析角度看,其花瓣数目与雄蕊数目和瓣化中的雄蕊数目存在正相关性,即花瓣的数目增多不完全依赖于雄蕊数目的减少,相反,花瓣数目较多的花朵中,其雄蕊数目也较多。在熊阳阳等对大花铁线莲花芽分化研究中发现大花铁线莲的重瓣起源仅涉及雌雄蕊起源,其花瓣的增多伴随着雌雄蕊由外向内的减少与退化[20];而孙明伟对百合的重瓣化研究中表明,百合的花器官变异包含花瓣褶皱面积变大、花器官的二次发育、雄蕊瓣化、花瓣与雄蕊的累积重复出现以及苞片瓣化等现象,其重瓣起源涉及积累起源与雌雄蕊瓣化起源,其中‘Siberia’品种的花被片数目与雄蕊数目同时增加,而‘Sornonne’品种的花被片数目与雄蕊数目减少[21-22]。结合重瓣垂丝海棠花瓣原基分化期时在第一轮花瓣原基内侧相继分化第二轮花瓣原基的情况可推测,重瓣垂丝海棠的重瓣化原因不仅涉及苞片与雌雄蕊起源,亦同时存在积累起源。
研究植物重瓣花成花的原因,必然会涉及到花发育与花器官发育的相关研究,其中研究最多的为ABCDE花发育模型相关基因和MADS-box基因家族[23]。虽然,现阶段的形态学证据可证明重瓣垂丝海棠的重瓣化起源为积累起源,苞片起源以及雌雄蕊起源,但其重瓣化起源存在多元化与复杂化的特点,需要展开更为深入的形态学研究,并结合重瓣垂丝海棠花器官分化过程的基因调控及表达特点等分子生物学方面的证据才可能获得科学的解释。
猜你喜欢
食品与生物技术学报(2021年8期)2021-01-17
广西植物(2020年13期)2020-09-12
广西植物(2020年13期)2020-09-12
山西农业科学(2018年9期)2018-09-12
现代园艺(2017年1期)2017-07-14
生物学教学(2017年6期)2017-02-18
农村百事通(2016年7期)2016-05-14
红蜻蜓·低年级(2016年9期)2016-05-14
花卉(2015年11期)2015-09-14
华南师范大学学报(自然科学版)(2014年6期)2014-08-28
