
外来入侵海洋红藻牛岛薄膜藻Haraldiophyllum udoense的形态观察及rbcL基因序列分析*
2022-01-19 05:58于雅楠王宏伟
海洋与湖沼 2022年1期
于雅楠 卞 瑶, 2 董 浩 王宏伟, 2
外来入侵海洋红藻牛岛薄膜藻的形态观察及L基因序列分析*
于雅楠1卞 瑶1, 2董 浩1王宏伟1, 2①
(1. 辽宁师范大学生命科学学院 辽宁大连 116081; 2. 辽宁省植物生物技术重点实验室 辽宁大连 116081)
为统计大连海域红藻的物种多样性, 在2016年5月至2021年5月进行标本采集时, 发现了红叶藻科入侵海藻牛岛薄膜藻M. S. Kim et J. C. Kang, 对其形态结构进行了详细的观察, 并对L基因序列进行了分析。结果表明: ①藻体直立; 呈暗红色或红褐色; 高7~30 cm, 宽6~25 cm; 叶片状, 薄膜质; 藻体除基部和生殖结构外为单层细胞。②L基因序列分析的结果显示本研究的6个样本间无碱基差异, 与产自韩国的牛岛薄膜藻无碱基差异。牛岛薄膜藻是在我国海域首次发现的新纪录种, 同时薄膜藻属Zinova为我国新纪录属, 认为船舶压舱水及海水流动是该种入侵的主要原因。本研究结果将丰富我国红叶藻科海藻的多样性, 同时为外来海藻的入侵途径及对当地生态的影响提供新资料。
牛岛薄膜藻; 外来入侵; 新纪录; 形态观察; 分子分析
牛岛薄膜藻M. S. Kim et J. C. Kang隶属于红藻门Rhodophyta、仙菜目Ceramiales、红叶藻科Delesseriaceae、薄膜藻属Zinova, 目前该属在全世界共报道了8个种, 模式种为(Kylin) Zinova (Paiano, 2020)。最早Kylin (1925)提出属的三个种Kylin、Kylin、Harvey果胞系发育过程相同, 但与属模种不同, 前者有两组不育细胞, 后者只有一组不育细胞。之后有学者根据这种生殖结构的不同, 建立薄膜藻属Zinova, 将Kylin、Kylin、Harvey移入该属, 产自大西洋的(Kylin) Zinova为该属模式种(Wynne, 1983)。Nam等(1996)最先在韩国济州牛岛发现了牛岛薄膜藻但错认为属模种, 随后Kim等(2011)基于形态观察和L基因序列分析确定该种为一新种, 命名为牛岛薄膜藻M. S. Kim etJ. C. Kang。该种主要分布于韩国西、南部及济州岛。
海洋运输业的高速发展使得外来物种的传播与入侵愈发严峻(刘艳等, 2013), 已成为海洋生态系统的危害之一(杨圣云等, 2001)。近年来一些大型有害藻类在大连海域被发现, 如舌状酸藻死亡后会分泌硫酸, 可导致其他藻类的死亡和水质污染(邱政, 2010); 硬毛藻属新纪录种强壮硬毛藻在大连海参养殖池中大量繁殖, 可将海参缠绕致死, 给海参养殖业造成了巨大损失(迟永雪等, 2009); 在黑石礁海域发现的具孔斯帕林藻, 藻体较大并在该地大量繁殖, 遮盖小型藻类, 影响其光合作用和发育繁殖, 可能会对该海域藻类物种多样性产生影响(宋学文等, 2018)。
2016年5月在辽宁省大连市黑石礁、付家庄、金石滩等海域发现了牛岛薄膜藻, 目前我国暂无关于该种的报道, 同时薄膜藻属也未见报道。在本研究中, 对其进行详细的形态结构观察和分子分析, 明确该种的分类地位, 讨论该种入侵途径。本研究结果将丰富我国红叶藻科海藻的多样性, 同时为外来海藻的入侵途径及对当地生态的影响提供新资料。
1 材料与方法
1.1 样本的采集和处理
海藻样本于2016年5月至2021年5月采自辽宁省大连市黑石礁、付家庄和金石滩, 样本处理参照李芳(2016)、宋学文等(2018)的方法, 对采集的样本进行编号, 新鲜藻体用于制作冰冻切片观察内部结构, 一部分进行硅胶干燥处理用于提取DNA, 藻体形态完好的制成腊叶标本, 保存于辽宁师范大学生命科学学院植物标本馆(LNU)。使用解剖镜和显微镜(Olympus BH2)对藻体形态结构进行观察并用Nikon HFX-Ⅱ A照相机拍照记录。
1.2 DNA提取与PCR扩增
使用植物基因组DNA提取试剂盒(TIAGEN, Valencia, CA, Beijing)对海藻样本的DNA进行提取, 并进行PCR扩增, 具体PCR参数参照李芳(2016)。应用软件Primer5.0设计引物, 引物组合见表1, 由上海生工生物公司合成。扩增后的产物用1%琼脂糖凝胶电泳检测, 检测后产物的纯化和测序由上海生工生物公司完成。

表1 rbcL序列扩增所用引物
1.3 rbcL基因序列分析
从GenBank上选取并下载薄膜藻属3个种、属3个种、属2个种和橡叶藻属4个种, 以及作为外群种的松节藻科鸭毛藻属鸭毛藻、凹顶藻属细枝凹顶藻与仙菜科仙菜属柔质仙菜(表2), 与本研究6个样本的L基因序列进行比对分析。
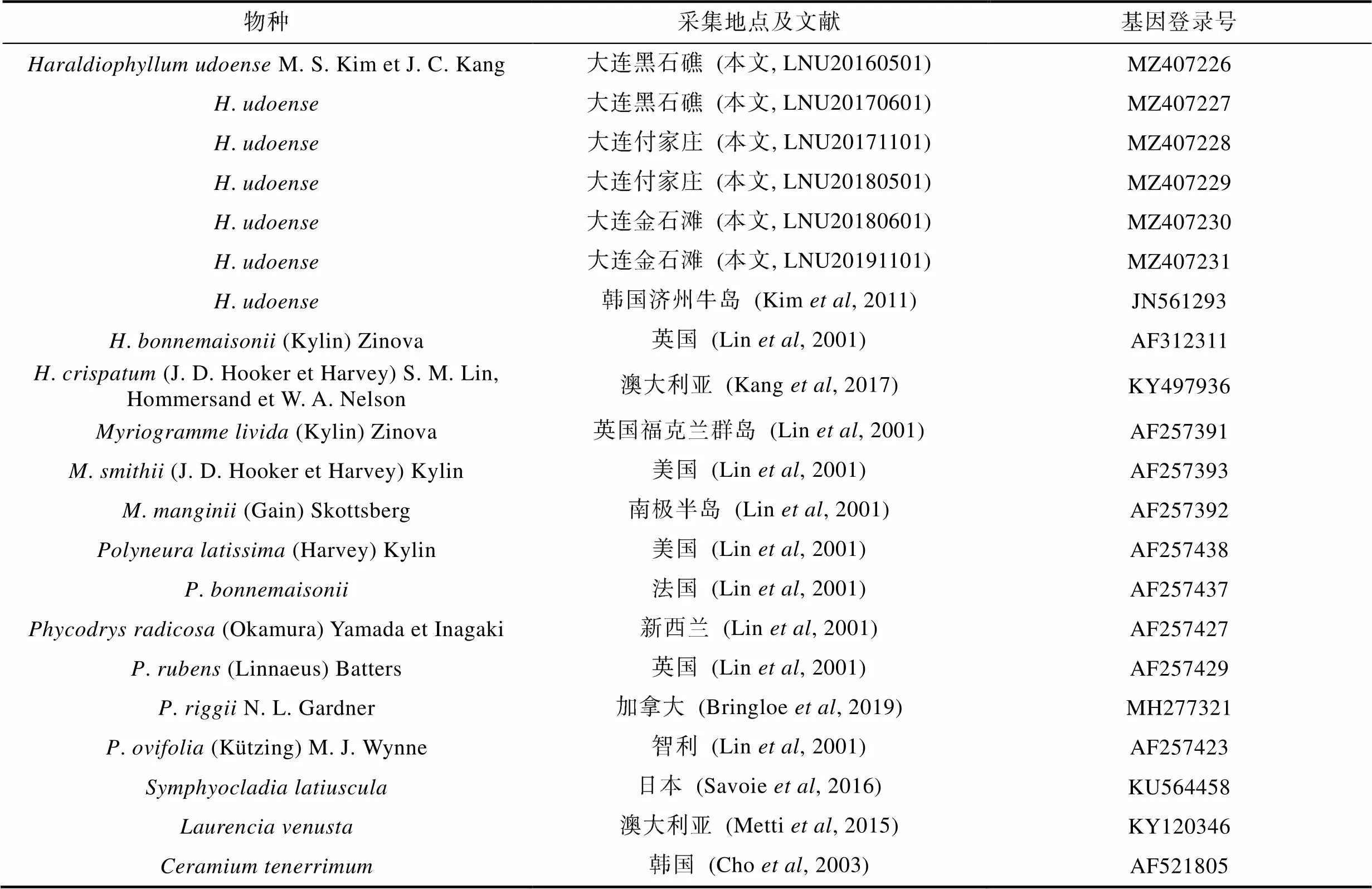
表2 rbcL基因序列分析所用样品信息
应用软件Clustalx (1.83)进行序列比对, 软件MEGAX进行碱基差异度分析以及系统树的构建, 使用最大似然法Maximum likelihood (ML)、邻接法Neighbor joining (NJ)和最大简约法Maximum parsimony (MP)构建系统发育树, 遗传距离计算模型为Number of differences和双参数遗传距离, Bootstrap重复1 000次。
2 结果
2.1 薄膜藻属(新拟)
Zinova, 1981
藻体直立, 薄膜质, 叶片宽线形至倒卵形, 深裂或浅裂, 边缘全缘、波状或具小齿, 无中脉和显微细脉; 固着器圆盘状, 具多个匍匐状假根; 皮层细胞呈多角边形, 藻体除基部和生殖结构外均为单层细胞; 雌雄异体, 配子体和四分孢子体同型; 果胞枝由4个细胞构成, 形成囊果时, 产孢丝与下部细胞发生融合, 在产孢丝顶端发育出单果孢子; 精子囊在藻体表面呈不规则状白斑; 四分孢子囊排列为两层, 呈四面锥形分裂。
模式种:(Kylin) Zinova, 1981
2.2 牛岛薄膜藻(新拟,图1)
M. S. Kim etJ. C. Kang, 2011
模式标本产地: 韩国济州牛岛
外部形态: 藻体直立, 高7~30 cm, 宽6~25 cm, 暗红色或红褐色; 叶片状, 薄膜质, 叶片全缘, 浅裂或深裂, 一至多个宽线性、卵形或倒卵形叶片; 基部具圆柱形短柄, 长2~3 cm, 厚约1 mm; 固着器圆盘状, 无中脉和显微细脉(图1a~1f)。

图1 牛岛薄膜藻的外部形态
注: a, b: 雌配子体; c, d: 四分孢子体; e, f: 雄配子体
营养结构: 藻体由单层多边形细胞构成, 无髓部(图2a, 2b), 厚约25 μm, 近基部细胞层数变多(图2c)。
生殖结构: 雌雄异体。成熟雌配子体上囊果近球形, 突出藻体表面呈红色斑点(图2d, 2e); 果孢子近球形, 由产孢丝发育形成, 大小约30~35 μm (图2f)。成熟雄配子体上精子囊群呈肉眼可见的白色条纹(图2g); 藻体单层细胞发生分裂, 形成精子囊母细胞(图2h, 2i), 随后形成瓶状的精子囊(图2h, 2j), 内有成熟精子, 大小约1~2 μm (图2j, 2k)。四分孢子囊群圆形或椭圆形, 在藻体表面略有隆起(图2l); 四分孢子囊四面锥形分裂, 成熟的四分孢子近球形, 大小约30~ 35 μm (图2m, 2n)。
习性: 生长在中、低潮带或潮下带的礁石上(图3)
产地: 大连市黑石礁(121°33′E, 38°52′N)、大连市付家庄(121°38′E, 38°52′N)、大连市金石滩 (121°59′E, 39°04′N)
国外分布: 韩国南部、西部海岸及济州岛
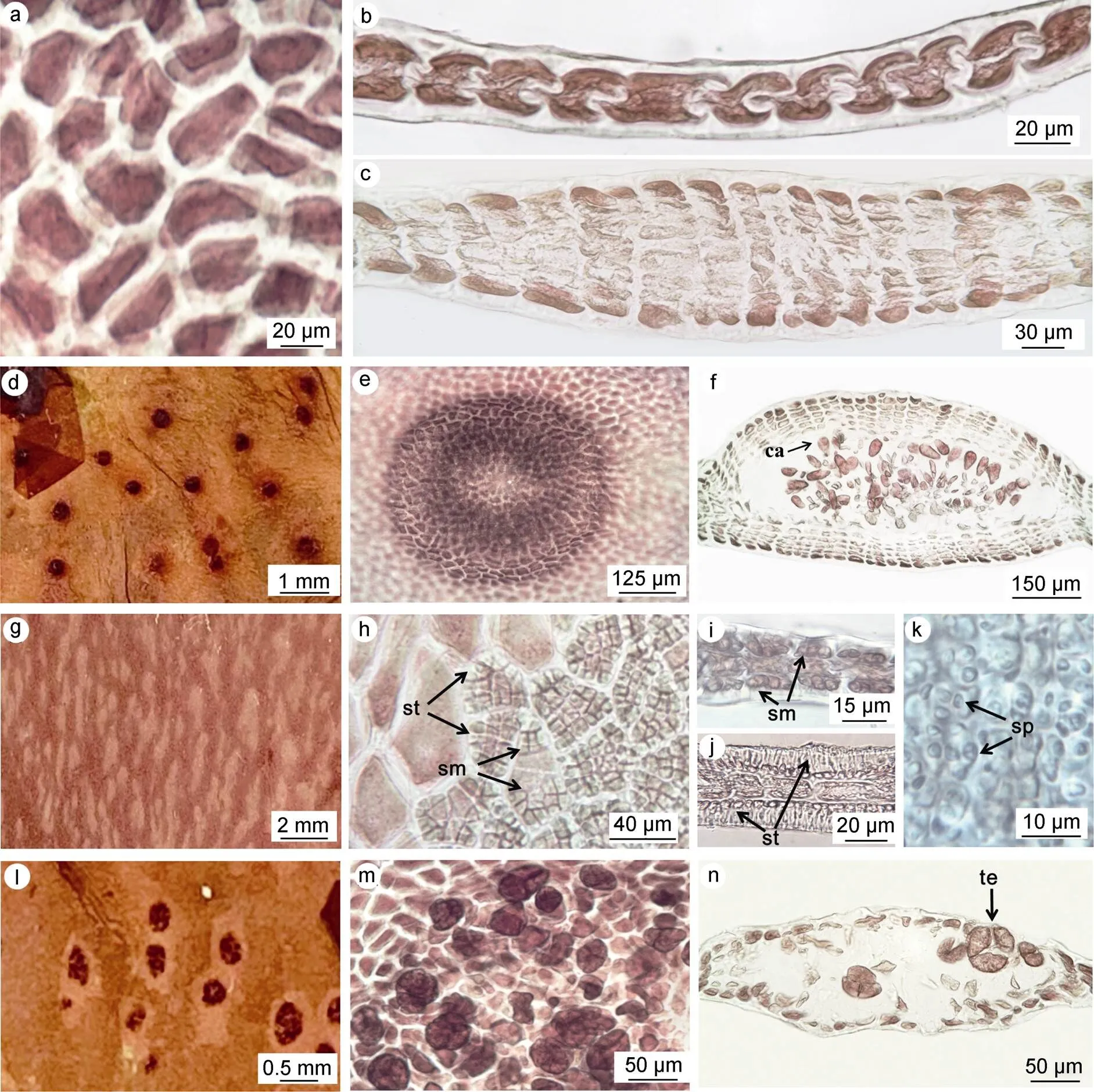
图2 牛岛薄膜藻的内部结构
注: a: 藻体表面观; b, c: 藻体横切面观; d, e: 雌配子表面观示囊果; f: 囊果横切面观(ca: 果孢子); g, h: 雄配子体表面观示精子囊群(sm: 精子囊母细胞; st: 成熟精子囊); i: 精子囊母细胞横切面观; j: 精子囊横切面观; k: 精子(sp); l, m: 四分孢子体表面观示四分孢子囊; n: 四分孢子囊横切面观(te: 成熟的四分孢子囊)

图3 牛岛薄膜藻的野外生境
2.3 rbcL基因序列分析
本研究6个样本的L基因序列长度为1 253 bp, 比对校正后的序列长度为1 253 bp。由ML系统发育树显示:本研究的6个样本与韩国产的牛岛薄膜藻聚在一支, 支持率为100%。与.和.聚集在一个大分支上, 支持率为100% (图4)。
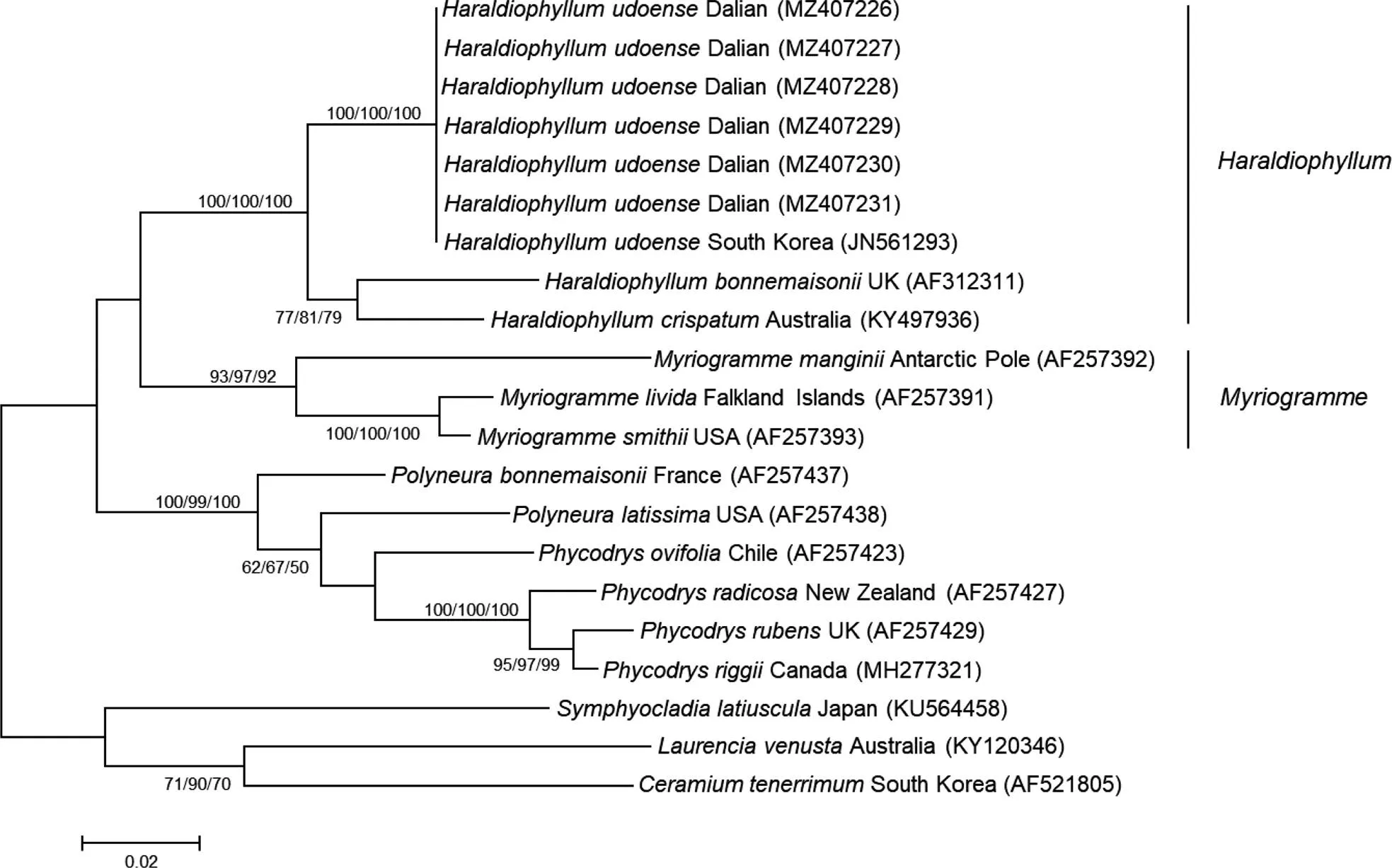
图4 基于rbcL基因序列构建的ML系统发育树
注: 节点数值表示支持率, 只显示大于50%, 从左至右分别为ML、MP、NJ
序列比对结果显示: 6个样本间无碱基差异, 与产自韩国的牛岛薄膜藻无碱基差异, 与.和.的碱基差异为71 bp (5.67%)、63 bp (5.03%), 与属3个种的碱基差异为114 bp (9.10%)~153 bp (12.21%), 与外群种鸭毛藻、细枝凹顶藻及柔质仙菜的碱基差异为192 bp (15.32%)、192 bp (15.32%)、199 bp (15.88%)。
3 讨论
采自大连的标本与韩国的牛岛薄膜藻(Kim, 2011)形态一致, 基于L基因序列构建的系统发育树显示, 本研究的6个样本与韩国产的牛岛薄膜藻聚在一支, 支持率为100%, 与薄膜藻属两个种和聚集在一个大分支上, 区别于其他种, 证明该藻为牛岛薄膜藻。关于薄膜藻属, Kang等(2017)根据牛岛薄膜藻的雌配子体生殖结构与该属其他种不同而将其分离出来, 建立新薄膜藻属, 将牛岛薄膜藻更名为牛岛新薄膜藻, 后有学者认为应将新薄膜藻属移回薄膜藻属 (Paiano, 2020), 我们认为根据生殖结构这一不同点来建立一新属是不严谨的, 不支持新薄膜藻属的建立, 因此本研究的外来入侵海藻为牛岛薄膜藻。该种是在我国首次发现的新纪录种, 同时薄膜藻属为我国新纪录属。
海洋外来物种的入侵一般通过有意和无意两种途径, 有意如物种引进, 无意如船舶压舱水(赵淑江等, 2005)。从日本海域入侵到大连市黑石礁的具孔斯帕林藻, 其入侵途径有三种即海洋养殖品种的引进、船舶压舱水和海水流动(宋学文等, 2018)。牛岛薄膜藻产自韩国, 而近年来大连市未从韩国引进海洋经济物种, 所以该藻可能通过船舶压舱水和海水流动这两种方式入侵到大连海域。据统计, 澳大利亚沿岸发现的外来海洋物种, 绝大多数通过船舶压舱水入侵(朱建庚等, 2006), 约100多种外来物种通过船舶压舱水入侵到美国多个水域(邵柏, 2001), 有害赤潮藻类通过船舶压舱水入侵, 已对当地生态系统造成了严重危害(王朝晖等, 2010)。我国对船舶压舱水的管理较为薄弱, 所以牛岛薄膜藻的孢子可能随与韩国往来的船只携带的压舱水进入大连海域。此外, 海水的流动也可能是该种入侵的原因之一。近些年全球变暖引发海平面上升(Chust2010), 可改变海水潮汐, 促进海水流动(Nicholls, 2010; 宋学文等, 2018), 地理位置上大连市和韩国毗邻黄海, 牛岛薄膜藻的孢子可能随海水从韩国济州岛流动到大连海域, 且大连市的气候类型和海洋环境与韩国相似, 适合该藻的生长繁殖。
外来入侵物种具备适应力强、繁殖力强、传播力强等特征, 可能会对当地生物种群、生态系统造成灾难性后果(邱政, 2010)。外来海藻由于没有栖息压力, 会大量占据资源而处于优势地位(陆琴燕等, 2013), 自在大连海域发现牛岛薄膜藻后, 每年都能在黑石礁、付家庄、金石滩等地发现, 且生物量逐年增多, 可见该藻在大连海域大量繁殖, 已经有了一定的规模。牛岛薄膜藻藻体较大, 可能会和当地小型藻类抢占生存空间, 通过遮盖影响其光合作用, 使其生存和繁殖受到限制(Paiano, 2020)。藻体腐烂分解需要消耗海水中的溶解氧, 可能出现局部缺氧的情况而导致其他海洋生物死亡。该种的出现可能对当地的物种多样性产生了一定影响。
4 结论
基于形态学观察和L基因序列分析, 发现牛岛薄膜藻M. S. Kim et J. C. Kang为我国新纪录种, 同时薄膜藻属Zinova为我国新纪录属。认为船舶压舱水及海水流动是该种入侵的主要原因, 其入侵已对当地的藻类物种多样性产生了一定影响。
王朝晖, 陈菊芳, 杨宇峰, 2010. 船舶压舱水引起的有害赤潮藻类生态入侵及其控制管理[J]. 海洋环境科学, 29(6): 920-922, 934.
朱建庚, 张露藜, 2006. 海洋外来物种入侵与我国压载水管理[J].生态经济, (10): 307-310.
刘艳, 吴惠仙, 薛俊增, 2013. 海洋外来物种入侵生态学研究[J].生物安全学报, 22(1): 8-16.
李芳, 2016. 四种红藻的形态学观察和分子系统学研究[D]. 大连: 辽宁师范大学: 50-51.
杨圣云, 吴荔生, 陈明茹, 等, 2001. 海洋动植物引种与海洋生态保护[J]. 台湾海峡, 20(2): 259-265.
邱政, 2010. 外来海洋生物入侵对我国水产养殖业的危害与预防[J]. 中国水产, (4): 21-22.
宋学文, 娄宇, 依朋, 等, 2018. 外来入侵海洋红藻具孔斯帕林藻的形态观察及分子系统分析[J]. 海洋与湖沼, 49(1): 78-86.
迟永雪, 王丽梅, 栾日孝, 等, 2009. 中国硬毛藻属新记录种—强壮硬毛藻[J]. 水产科学, 28(3): 162-163.
陆琴燕, 刘永, 李纯厚, 等, 2013. 海洋外来物种入侵对南海生态系统的影响及防控对策[J]. 生态学杂志, 32(8): 2186-2193.
邵柏, 2001. 加强入境船舶压舱水管理的新观念[J]. 中国国境卫生检疫杂志, 24(1): 39-42.
赵淑江, 朱爱意, 张晓举, 2005. 我国的海洋外来物种及其管理[J]. 海洋开发与管理, 22(3): 58-66.
BRINGLOE T T, SAUNDERS G W, 2019. Trans-Arctic speciation of Florideophyceae (Rhodophyta) since the opening of the Bering Strait, with consideration of the “species pump” hypothesis [J]. Journal of Biogeography, 46(4): 694-705.
CHO T O, FREDERICQ S, BOO S M, 2003.sp. nov. (Ceramiaceae, Rhodophyta) from Korea: a new species based on morphological and molecular evidence [J]. Journal of Phycology, 39(1): 236-247.
CHUST G, CABALLERO A, MARCOS M,, 2010. Regional scenarios of sea level rise and impacts on Basque (Bay of Biscay) coastal habitats, throughout the 21st century [J]. Estuarine, Coastal and Shelf Science, 87(1): 113-124.
KANG J C, YANG M Y, KIM M S, 2017., a new genus of Delesseriaceae (Rhodophyta) based on carposporophyte development and molecular data [J]. Botanica Marina, 60(5): 515-532.
KIM M S, KANG J C, 2011. A new Korean red algal species,sp. nov. (Delesseriaceae, Rhodophyta) [J]. Algae, 26(3): 211-219.
KYLIN H, 1925. The marine red algae in the vicinity of the Biological Station at Friday Harbor, Wash [J]. Lunds Universitets Årsskrift, Ny Följd, Andra Afdelningen, 21(9): 1-87.
LIN S M, FREDERICQ S, HOMMERSAND M H, 2001. Systematics of the Delesseriaceae (Ceramiales, Rhodophyta) based on large subunit rDNA andL sequences, including the Phycodryoideae, subfam nov. [J]. Journal of Phycology, 37(5): 881-899.
METTI Y, MILLAR A J K, STEINBERG P, 2015. A new molecular phylogeny of thecomplex (Rhodophyta, Rhodomelaceae) and a review of key morphological characters result in a new genus,, and a description of.[J]. Journal of Phycology, 51(5): 929-942.
NAM K W, KIM Y S, 1996.(Rhodophyta): a Delesseriaceous genus newly recorded in Korea [J]. Algae, 11(1): 101-106.
NICHOLLS R J, CAZENAVE A, 2010. Sea-level rise and its impact on coastal zones [J]. Science, 328(5985): 1517-1520.
PAIANO M O, HUISMAN J M, CABRERA F P,, 2020.sp. nov. (Delesseriaceae, Rhodophyta): a new mesophotic genus record for the Hawaiian Islands [J]. Algae, 35(4): 337-347.
SAVOIE A M, SAUNDERS G W, 2016. A molecular phylogenetic and DNA barcode assessment of the tribe Pterosiphonieae (Ceramiales, Rhodophyta) emphasizing the Northeast Pacific [J]. Botany, 94(10): 917-939.
Wynne M J, 1983. The current status of genera in the Delesseriaceae (Rhodophyta) [J]. Botanica Marina, 26(9): 437-450.
MORPHOLOGICAL ANDL GENE SEQUENCE ANALYSES ON INVASIVE MARINE RED ALGAE,
YU Ya-Nan1, BIAN Yao1, 2, DONG Hao1, WANG Hong-Wei1, 2
(1. College of Life Sciences, Liaoning Normal University, Dalian 116081, China; 2. The Key Laboratory of Plant Biotechnology of Liaoning Province, Dalian 116081, China)
In order to count the species diversity of red algae in Dalian sea areas,M. S. Kim et J. C. Kangin the family Delesseriaceae was discovered as an invasive species during May 2016 to May 2021 when we collected specimens. Detailed morphological structure was observed and theL gene sequence was analyzed. Results show that the thallus is erect, dark red or reddish brown, 7~30 cm high and 6~25 cm wide, leaf-like, membrane quality. The algae is monostromatic except for the lower portion and reproductive structures. TheL gene sequence analysis showed no divergence between our six samples ofand sequence from Korea. Both the species ofand the genusZinova are the new record in China. We infer that the ship ballast water and seawater flow are the main reasons for this species invasion.
; foreign species; new record; morphological observations; biomolecular analysis
Q943.2
10.11693/hyhz20210700155
*国家自然科学基金项目, 31270251号。于雅楠, 硕士研究生, E-mail: 1643136760@qq.com
王宏伟, 教授, 博士, 博士生导师, E-mail: kitamiwang@163.com
2021-07-06,
2021-08-27
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
广西科学(2022年1期)2022-04-12
中国化肥信息(2020年6期)2020-11-20
今日农业(2020年15期)2020-09-25
水产科学(2019年4期)2019-07-23
中南民族大学学报(自然科学版)(2018年4期)2018-12-29
科技创新与应用(2017年20期)2017-07-15
中国船检(2017年3期)2017-05-18
中国塑料(2015年3期)2015-11-27
中国塑料(2015年9期)2015-10-14
