
普通烟草生长素抑制因子NtIAA17的克隆与功能分析
2022-01-17 06:44:48王驰尹卓然轩栋栋连文力罗勇张萍符云鹏贾宏昉
中国烟草学报 2021年6期
王驰,尹卓然,轩栋栋,连文力,罗勇,张萍,符云鹏,贾宏昉*
1 河南农业大学烟草学院,郑州 450002;2 厦门大学生命科学学院,厦门 361000
全球土壤盐渍化面积约为9.5×108hm2,我国土壤盐渍化面积约占三分之一,大面积土壤资源难以利用[1]。化肥的过量使用使土壤盐渍化程度越来越严重,从而影响植物生长和作物产量品质[2],盐胁迫是限制全球农作物产量的非生物胁迫之一[3]。土壤中盐分过多,会抑制植物进行光合作用、内源激素的合成和运输、离子吸收等,影响植物的生长发育过程[4]。
生长素作为一种植物激素,能够与植物体内的多种其他激素共同作用,参与从胚胎发生到衰老,调节植物的生长发育,以及响应盐胁迫、干旱等非生物胁迫[5]。生长素/吲哚-3-乙酸(Aux/IAA)蛋白作为响应生长素信号的关键因子[6],在生长素信号转导过程中起重要的调控作用[7]。当Aux/IAA在生长素浓度较低时,与ARF形成ARF-Aux/IAA异质二聚体从而抑制ARF的转录活性;当生长素浓度较高时,生长素受体(TIR1/AFBs)与生长素结合后导致Aux/IAAs发生降解,ARF从Aux/IAA中释放,从而调控下游基因表达[8]。
Aux/IAA家族基因已在许多植物中得到鉴定分析[9],在拟南芥中有29个[10],在水稻中有31个[11],在马铃薯中有26个[12]等。拟南芥AtIAA19过量表达,会显著促进植物根、茎、叶的生长[13]。StIAA2基因的下调表达使植株高度增加、叶片下垂、顶叶原基弯曲生长[14]。葡萄VvIAA18基因的过表达提高葡萄的抗氧化能力进而增强了对盐胁迫的耐受性[15]。盐胁迫和干旱胁迫均使高粱SbIAA1基因在根和叶中上调表达,该基因可能提高高粱的抗盐抗旱能力[16]。以上结果表明,Aux/IAA家族对植物的生长发育起着重要的调控作用,并且可能提高植株对非生物胁迫的耐受性。
目前,对烟草基因组数据的生物信息学分析表明,烟草中至少有77个Aux/IAA成员[17],但这些成员的功能仍不清楚。前期,从高盐胁迫的转录组数据库发现NtIAA17受盐胁迫增强表达。因此,本研究以K326为试验材料,克隆得到NtIAA17基因,对其生物学功能和表达模式进行系统分析,探究NtIAA17基因在抵御盐胁迫中的功能,为培育烟草耐盐新品种提供理论基础。
1 材料和方法
1.1 植物材料与生长条件
使用野生型(Nicotiana tabacumcv,K326)烟草和NtIAA17基因过表达的K326转基因烟草用于本研究。
首先挑选均匀饱满的烟草种子在75%(v/v)乙醇溶液消毒30 s,然后在10%(v/v)次氯酸钠溶液中消毒7 min,用高温灭菌处理后的蒸馏水洗涤6次。将种子播种在海绵育苗盘中,用蒸馏水暗培养7 d催芽[19],然后放置在人工智能培养箱中培养21 d(白天28℃,14 h,晚上22℃,10 h,每24 h循环;湿度设置为60%),1~2 d使用蒸馏水培养,3~5 d(约长至两片子叶)使用1/4 Hoagland营养液培养,5~7 d使用1/2 Hoagland营养液培养,然后7~14 d使用Hoagland营养液培养。
1.2 NtIAA17 基因全长cDNA的克隆
使用Eastep®Super Total RNA Extraction Kit(上海普洛麦格生物产品有限公司)提取烟草根、茎和叶组织部位的总RNA,测定所提总RNA的OD260/280值,并用1.2%琼脂糖凝胶电泳检测其完整性,使用HiScript®III 1st Strand cDNA Synthesis Kit(+gDNA wiper)试剂盒(南京诺唯赞生物技术有限公司)进行反转录,得到cDNA。采用Primer Premier 5.0设计烟草NtIAA17全长基因序列扩增引物(F:5’-ATGTCATCGGAGAAATTCAAAG-3’;R:5’-TTAGCTT GTACTCGAGCATTTC-3’),以反转录的根、茎、叶混合cDNA为模板进行PCR扩增,PCR产物末端加polyA后经电泳回收后克隆到pMD19-T载体(TaKaRa公司),转化大肠杆菌DH5α感受态细胞,挑选阳性单克隆,然后送武汉伯远生物科技有限公司进行测序,获得全长 cDNA 序列信息[20]。

表1 qRT-PCR引物序列Tab. 1 qRT-PCR primer sequence
1.3 NtIAA17基因生物信息学分析
对测序结果进行生物信息学分析,使用ProtParam (http://web.expasy.org/protparam/) 预 测基因编码数量、蛋白质相对分子量和理论等电点;用NCBI数据库查找相关基因及其氨基酸序列,使用DNAMAN6.0(Lynnom Biosoft,美国)对氨基酸序列进行比对;使用PlantCare(http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/)检测启动子区域中含有的顺式作用元件;通过MEGA6.0软件中的Neighbor-Joinning(NJ)算法构建了系统发育树[21]。
1.4 NtIAA17-GFP融合蛋白的亚细胞定位
在烟草原生质体中分析NtIAA17的亚细胞定位,使用聚乙二醇介导的转化将重组35S:NtIAA17-GFP和35S:GFP质粒导入从叶片组织中获得的烟草原生质体中。用共聚焦激光扫描显微镜在转化后16~20 h捕获荧光信号[22]。核定位标记蛋白(NLS)如Shcherbo et al,2007所述[23]。
1.5 NtIAA17基因表达模式分析
1.5.1NtIAA17基因的组织特异性表达分析
培养10周的烟株用NaCl (150 mmol/L) 胁迫处理7 d,分别提取样品根、茎、新叶和老叶(新叶为完全展开的叶片从上往下数第二片,老叶为完全展开的叶片从上往下数第七片)的总RNA,反转录成cDNA,并分析NtIAA17基因在烟草各个组织中的相对表达量。
1.5.2 盐胁迫时间长度下烟草NtIAA17基因表达分析
培养约21 d的烟苗用NaCl (150 mmol/L) 胁迫处理,分别于0 h、1 h、2 h、4 h、6 h、12 h后,提取烟草的总RNA,反转录成cDNA,并分析NtIAA17基因在NaCl(150 mmol/L)处理后的相对表达量变化情况。
1.5.3 实时荧光定量PCR
提取烟草根、茎和叶组织部位的总RNA,反转录后作为qRT-PCR反应的模板。烟草IAA17基因扩增引物qRT-NtIAA17的设计采用Primer Premier 5.0 软件。qRT-PCR反应用Taq Pro Universal SYBR qPCR Master Mix 荧光定量试剂盒(南京诺唯赞生物技术有限公司)说明书进行,3次重复,内参基因为烟草组成型表达基因NtL25(NCBI登录号:L18908.1)。数据用2-△△Ct法处理。
1.6 pCAMBIA1305-NtIAA17过表达载体的构建及遗传转化
根据基因克隆测序正确的NtIAA17基因序列设计构建过量表达载体的引物(NtIAA17-OX),以测序正确的NtIAA17cDNA克隆为模板扩增NtIAA17的全长序列(NCBI ID:XM_016635452.1),将PCR产物连接到pCAMBIA1305载体中,该载体由花椰菜花叶病毒(CaMV)35S启动子和胭脂碱合成酶终止子驱动,转化DH5α感受态细胞,经过菌液PCR检测对比后,得到过表达载体pCAMBIA1305-NtIAA17。利用电转化将其转入农杆菌GV3101感受态细胞中,筛选得到阳性克隆,用阳性菌液侵染野生型烟草(Nicotiana tabacumcv,K326)中[20],室内繁种,收取转基因植株T0代种子,而后加代获得T2代纯合植株用于本研究。
1.7 转基因烟草株系的耐盐性鉴定
1.7.1 观察烟草表型并记录整株生物量
将野生型和过表达株系OX1、OX2用上述水培法培养约21 d,用NaCl(150 mmol/L)胁迫处理7 d后,观察60株烟苗的整体表型并记录单株生物量。
1.7.2 MDA,脯氨酸的测定
通过MadhavaRao和Sresty(2000)的方法测定MDA含量[24]。使用茚三酮比色法测定游离脯氨酸含量[25]。
1.7.3 抗氧化酶相关基因的测定
qRT-PCR反应用Taq Pro Universal SYBR qPCR Master Mix 荧光定量试剂盒(南京诺唯赞生物技术有限公司),测定NtSOD、NtPOD基因的相对表达量。
1.8 统计分析
使用t检验比较两个样本组,结果用平均值和相应的标准误差表示。使用IBM SPSS Statistics软件版本21(IBM)进行统计分析。图片使用Origin2018(Origin Labinv,Northampton,MA,USA)软件绘制。
2 结果与分析
2.1 NtIAA17的克隆和序列分析
以烟草cDNA为模板,利用特异性引物NtIAA17-OX进行PCR扩增,获得NtIAA17基因片段, 目标条带与琼脂糖凝胶电泳显示的结果大小一致(Marker条带为2000 bp)(图1),对测序结果进行分析表明,NtIAA17基因片段为 624 bp,编码207个氨基酸(图2),NtIAA17蛋白的相对分子质量为23.07 kD,理论等电点(pl)为8.24。

图1 NtIAA17基因的PCR扩增Fig. 1 PCR amplification of NtIAA17 gene

图2 烟草NtIAA17的全长cDNA序列及其编码的氨基酸Fig. 2 cDNA full length sequence and deduced amino acid sequence of tobacco NtIAA17
2.2 NtIAA17蛋白生物信息学分析
2.2.1NtIAA17基因同源序列比对及进化分析
利用DNAMAN软件将NtIAA17与其他物种氨基酸序列进行同源比对,结果显示,NtIAA17的蛋白序列具有Aux/IAA家族的四个典型结构域(I-IV),表明NtIAA17是烟草Aux/IAA蛋白(图3)。利用MEGA 6.0软件进行系统进化树分析表明NtIAA17基因与菠菜MoIAA13基因(ID:XM_021983619.1)的亲缘关系最近(图4)。

图3 NtIAA17与其同源氨基酸序列比对Fig. 3 Sequence alignment of NtIAA17 and its homologous amino acid

图4 烟草NtIAA17及其同源氨基酸序列的进化树分析Fig. 4 Phylogenetic tree analysis of sequence of tobacco NtIAA17 and its homologous amino acid
2.2.2NtIAA17基因启动子序列分析
由图5可知,NtIAA17启动子区含有多种顺式作用元件,例如防卫和胁迫响应顺式作用元件,如W-box、MYB;盐胁迫诱导型启动子,如G-box、Box-III;植物激素响应元件,如TGA-element、Auxr r-core、ABRE。
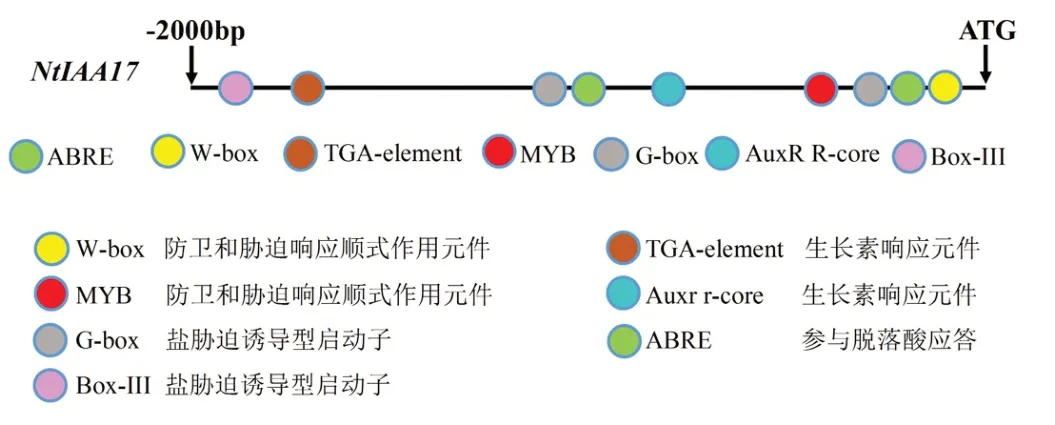
图5 NtIAA17的启动子序列分析Fig. 5 Promoter sequence analysis of NtIAA17
2.2.3 NtIAA17亚细胞定位分析
通过在烟草原生质体中验证NtIAA17的亚细胞定位发现,如图6所示,NtIAA17-GFP的GFP荧光位置与NLS核定位标记蛋白的位置完全匹配,结果表明NtIAA17定位在细胞核内。

图6 NtIAA17的亚细胞定位Fig. 6 Subcellular localization of NtIAA17
2.3 NtIAA17基因的表达模式分析
对检测不同烟草组织中NtIAA17基因的组织特异性表达的结果表明,NtIAA17在茎中表达量最高, 老叶中其次, 新叶中最低(图7A)。培养21 d的烟苗用高盐NaCl(150 mmol/L)胁迫处理,分别于0 h,1 h,2 h,4 h,6 h,12 h取样测定烟草中NtIAA17的相对表达水平,结果显示高盐胁迫上调烟草中NtIAA17基因的表达,在2 h时,NtIAA17的表达量最高(图7B)。这些发现表明,NtIAA17基因参与了植物生长发育,盐胁迫可以调控NtIAA17基因表达。
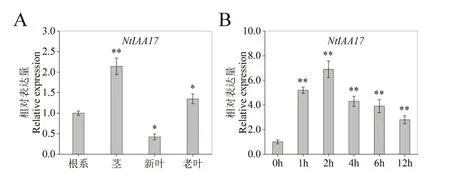
图7 NtIAA17在烟草不同器官和盐胁迫时间长度的表达模式。Fig. 7 Expression patterns of NtIAA17 in different tobacco organs under different salt stress duration
2.4 盐胁迫对NtIAA17转基因烟草生物量表型及抗氧化活性的影响
为了研究NtIAA17的功能,通过转基因技术获得了10个过量表达烟草株系(T0代转基因烟草株系与野生型K326植株在大田生育期无显著差异),进一步通过qRT-PCR分析鉴定了两个过量表达效果较好的烟草转基因株系(命名为OX1和OX2)(图8A)。为了研究盐胁迫下NtIAA17的功能,检测了烟草的耐盐表型和单株生物量。在对照条件下,过表达NtIAA17-OX和野生型(WT)烟草之间的表型和生物量没有显著差异。在高盐胁迫NaCl(150 mmol/L)条件下,NtIAA17-OX烟株的长势较好,生物量更高(图8B,C,D),这表明NtIAA17的过表达可以增加烟草的生物量,缓解盐胁迫带来的伤害。通过测定丙二醛(MDA)和脯氨酸含量可知,与高盐胁迫下的野生型相比,过表达NtIAA17烟草植物显著减少了MDA的积累,增加了游离脯氨酸的含量(图8E,F),进一步对抗氧化酶相关基因进行qRT-PCR分析发现,在盐胁迫条件下,NtIAA17-OX烟草的NtSOD、NtPOD的表达量明显高于野生型(图9)。这表明NtIAA17的过表达可以增强烟草的抗氧化能力。

图8 盐胁迫对NtIAA17转基因烟草表型和抗氧化活性的影响Fig. 8 Effect of salt stress on the phenotype and antioxidant activity of NtIAA17 transgenic tobacco

图9 盐胁迫对NtIAA17转基因烟草抗氧化酶基因的影响Fig. 9 Effect of salt stress on antioxidant enzyme genes of NtIAA17 transgenic tobacco
3 讨论
Aux/IAA蛋白是生长素反应因子,通过与生长素响应因子ARF蛋白相互作用来介导生长素信号通路[26],可调节下游多种生长素反应基因的表达,该基因在调控植物的生长发育中发挥重要作用[27]。先前的研究表明,Aux/IAA家族基因的功能在许多作物中均有研究,但关于Aux/IAA家族在烟草中的功能研究还不够深入。在本研究中,从烟草中分离了NtIAA17基因,并首次在烟草中进行了功能研究。NCBI网站上的Blastp分析显示,NtIAA17与其他植物Aux/IAA基因具有高度相似性,并且与菠菜MoIAA13(ID:XM_021983619.1)的关系最为密切。Aux/IAA 基因家族是一个生长素诱导表达的多基因家族[28],包含四个保守的结构域I、II、III和IV。结构域I具有转录抑制功能[29]。结构域II是生长素信号转导的必需元件,与F-box蛋白TIR1相互作用,导致Aux/IAA的不稳定性和快速降解[30]。结构域III和IV能与其他Aux/IAA或ARF相互作用,形成同源二聚体和异源二聚体,抑制生长素反应基因转录,影响信号转导[31]。NtIAA17具有四个高度保守的结构域(I-IV),是典型的Aux/IAA蛋白,属于Aux/IAA家族成员,此外,亚细胞定位结果显示NtIAA17蛋白位于细胞核中。
启动子作为调控基因表达的重要元件, 通过分析启动子区所含有的顺式作用元件可以推断下游基因编码蛋白可能具有的功能[32],其中的顺式作用元件可以与转录因子相结合,应答相关基因转录[33]。启动子分析结果表明,烟草NtIAA17基因的启动子区域存在有多种激素响应和胁迫响应的顺式作用元件,其中发现NtIAA17启动子存在盐胁迫诱导型元件和生长素响应元件,推断该基因可能参与调控下游生长素相关基因表达,并可能在调节烟草的生长发育和耐盐性中起关键作用。
Aux/IAA家族基因广泛参与植物生长发育的调控,还可以参与响应不同环境胁迫[34]。前人研究发现,香蕉MaIAA20、MaIAA25均可响应干旱、高盐、高温等逆境胁迫,在盐胁迫下,MaIAA15也显著上调表达,增强香蕉的耐盐性[35]。水稻OsIAA6基因由干旱胁迫高度诱导,其在转基因水稻中过表达,可能通过调控生长素蛋白生物合成基因提高对干旱胁迫的耐受性[36]。本实验中,NtIAA17基因的表达模式分析表明,NtIAA17基因在烟草的根、茎、新叶和老叶中均有表达,但表达量存在明显差异,在茎和老叶中高表达,结果表明NtIAA17基因具有组织特异性并参与烟草生长发育过程。NtIAA17的表达水平是在盐胁迫条件下诱导的,并且在盐胁迫处理2 h的表达量最高,这表明NtIAA17可能在调节烟草的耐盐性中起重要作用。
为了验证NtIAA17的功能,通过转基因技术,研究了过量表达NtIAA17对烟草生长发育的影响。通过分析在高盐胁迫(NaCl 150 mmol/L)条件下的表型和生物量发现,与野生型烟草相比,NtIAA17过表达显著增加了转基因植物的生物量,这表明NtIAA17基因参与调控在盐胁迫下的生长发育。
前人的研究表明,渗透胁迫通常会导致更多的脯氨酸积累,而脯氨酸的积累水平与耐盐性有关[37-38]。在这项研究中,盐胁迫条件下,NtIAA17-OX转基因烟草植物的脯氨酸含量明显高于野生型,以保护细胞膜免受盐诱导的伤害,并参与细胞质渗透压平衡的调节,提高植物对盐胁迫的耐受性。而MDA含量可反映膜脂过氧化损伤程度[39],在本研究中,与野生型相比,盐胁迫下,NtIAA17-OX转基因烟草植物中MDA含量积累显著降低。NtIAA17过表达可调节盐胁迫下抗氧化酶相关基因如NtSOD、NtPOD的表达,因此,以上结果表明,NtIAA17-OX转基因植物提高烟草在高盐胁迫下的生物量可能是由于提高了烟草的抗氧化能力。
4 结论
Aux/IAA 家族基因参与调控植物生长发育过程,本研究首次从普通烟草K326中克隆并鉴定了一个Aux/IAA家族基因NtIAA17,亚细胞定位结果表明该蛋白是一个核蛋白,调控烟草的生长发育;启动子和表达模式分析表明该基因受盐胁迫诱导表达,参与应答盐胁迫反应。进一步通过转基因技术,获得NtIAA17基因过量表达的转基因株系,生理试验初步表明NtIAA17可以通过调控烟草的抗氧化能力,抵御盐胁迫带来的危害,增加转基因烟草的在盐胁迫下的生物量,影响烟株的生长发育;本研究为后续普通烟草耐盐胁迫分子机制研究和耐盐新品种的培育提供候选基因和理论依据。
猜你喜欢
学与玩(2022年10期)2022-11-23 08:32:00
今日农业(2022年3期)2022-06-05 07:12:08
生物学通报(2021年4期)2021-03-16 05:41:34
今日农业(2020年19期)2020-12-14 14:16:52
科学大众(2020年12期)2020-08-13 03:22:38
乡村科技(2019年9期)2019-06-26 03:52:04
数码世界(2018年7期)2018-12-24 15:06:05
创新科技(2015年1期)2015-12-24 06:23:21
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
