
黄龙山林区不同演替时期典型树种光诱导的气孔动力学研究
2022-01-15 05:35赵霖玉李秧秧
西北植物学报 2021年11期
赵霖玉,李秧秧
(1 西北农林科技大学 林学院, 陕西杨陵 712100;2 黄土高原土壤侵蚀与旱地农业国家重点实验室, 西北农林科技大学 水土保持研究所, 陕西杨陵 712100)
光是影响气孔开闭的主要环境因子。自然界中植物由于叶片遮挡、云层覆盖和太阳倾角的变化,用于光合作用的光量子强度(PPFD)一直在发生变化。在弱光→强光诱导的气孔开放过程中,气孔导度(gs)对光强变化的响应通常比光合速率(A)慢一个数量级,从而导致gs直接限制A;而在强光→弱光诱导的气孔关闭过程中,慢速气孔关闭导致水分的无效损失,最终影响植物的生产力和水分利用效率[1-3]。因此,加快光诱导过程中gs的响应可能是提高植物生产力和水分利用效率的一条有效途径。
不同物种[3-5]、同一物种不同基因型[6-9]或同一物种不同生长发育阶段[10]光诱导的gs动力学响应过程存在明显差异。此外,植物生长的环境条件,如土壤水分、光照、大气CO2浓度和相对湿度等亦影响其光诱导的gs动力学响应过程[7-8,10-11]。目前,关于植物光诱导的gs动力学变化机制并不清楚,一般认为gs响应快慢与气孔特征(如保卫细胞类型、气孔密度和大小等)有密切的关系。具有哑铃型保卫细胞的植物其gs响应速率快于具有肾形保卫细胞的植物[3],斑克木属植物中具有小气孔和高气孔密度的植物种具有更高的gs增加速率,且小气孔种达到响应幅度50%所需的时间更短[4]。但气孔特征无法解释在一系列植物上发现的气孔关闭速率和气孔大小无关[5,12]及具有肾形保卫细胞的不同物种间气孔动力学的差异[3],这表明其他因素,如保卫细胞结构(如细胞骨架和细胞壁弹性)及生物化学因素(如转运体或离子通道的数量和活性等)等可能在光诱导的gs动力学反应中起着十分重要的作用。
光和水分是影响黄土高原地区植被演替的主要因素,研究表明,植被演替早期种一般具有高的光合速率和水分利用效率(WUE)[13-15],尽管也有相反的报道[16]。这种高光合速率和高WUE有利于演替早期种进行快速生长及适应演替早期开放的光环境。演替早期种高的WUE是否与其快速的光诱导气孔动力学有关尚不清楚。对热带雨林2种演替早期种和9种演替后期种光诱导气孔动力学的研究表明,2种演替早期树种的气孔开放速率和T50%(达到稳态气孔导度50%所需时间)分别为0.06~0.26 mmol·m-2·s-2和5~14 min,而9种演替后期种的气孔开放速率和T50%分别为0.06~0.12 mmol·m-2·s-2和5~17 min,不同演替位树种的气孔开放速率和T50%存在重叠,并未因演替位的不同而存在差异[17]。在受水分限制的黄土高原地区植被演替中演替早期种和后期种的气孔动力学是否亦如此尚不清楚。为此,本研究对黄土高原黄龙山林区3种不同演替阶段树种幼苗光诱导的气孔动力学进行了探讨,试图回答以下问题:1)不同演替阶段树种光诱导的气孔动力学参数是否存在差异? 2)这种动力学参数差异是否与其气孔特征有关? 3)不同树种光诱导的气孔动力学差异是否可以解释其水分利用效率的差异? 对这些问题的回答,有助于阐释黄土高原地区不同演替阶段树种水分利用效率差异的生理基础,从而为半干旱区树种选择和水分调控提供一定的理论依据。
1 材料和方法
1.1 试验材料
试验用树种为黄龙山林区演替早期种山杨(Populusdavidiana)和白桦(Betulaplatyphylla)以及演替后期种辽东栎(Quercusliaotungensis)的实生苗,采用盆栽试验,在陕西杨陵西北农林科技大学水土保持研究所盆栽试验场进行。2021年3月下旬在陕西黄龙县蔡家川林场挖取3种树种的实生苗(1~3年生),每种树种大约15株,带回到西北农林科技大学水土保持研究所。挑选生长均一的实生苗栽植于塑料盆中,试验用土为蔡家川林场林地表层土壤,其土壤容重、有机质、全N、碱解N和Olsen-P含量分别为1.26 g·cm-3、12.56 g·kg-1、1.05 g·kg-1、78.26 mg·kg-1和30.5 mg·kg-1。土壤经风干、粉碎过筛后装入塑料盆(上底内径28.5 cm、下底内径21.0 cm、高24.5 cm),每盆装土14 kg,栽植树苗1株,每个树种栽植10盆。3种树种幼苗于2021年3月25日栽种,然后浇水至田间最大持水量。山杨和白桦4月初开始萌发生长,辽东栎4月中旬开始萌发生长。在生长过程中,土壤含水量一直维持在田间持水量的75%~85%,每隔2周,用Hoagland营养液进行养分补充,每次补充500 mL完全Hoagland营养液。于2021年6月20日(生长约2个多月后)每个树种挑选无病虫害的健康株8株,对冠层上部完全展开的成熟叶进行气孔导度动力学、气孔特征、光合能力和碳同位素分辨率(δ13C)等的测定。
1.2 测定项目及方法
1.2.1 气孔导度动力学参数使用LI-6400便携式光合作用系统(Li-COR, Lincoln, USA)测定最上部完全展开叶的气孔导度动力学参数。植物在室内恒定环境下提前适应一晚,整个测量过程按照McAusland等[3]的光诱导方法,涉及从弱光→强光诱导的气孔开放过程和从强光→弱光诱导的气孔关闭过程。即叶片先在100 μmol·m-2·s-1PPFD下平衡至A和gs达到稳定状态(一般持续0.5 h),然后将光强增加到1 200 μmol·m-2·s-1持续测定1 h后,再返回至100 μmol·m-2·s-1直至新的稳定状态(0.5 h左右)。测定时叶室条件设置为:叶温25 ℃,CO2浓度400 μmol·mol-1,相对湿度60%。整个测定过程在生长箱中进行,使用自动打点每隔30 s记录一次数据。考虑到植株昼夜生理节律的影响,所有测量均在8:00~15:00之间进行。每个处理重复4次。
1.2.2 气孔解剖特征测量完叶片气体交换参数后的叶片用来进行气孔解剖特征的测定,采用指甲油印迹法制成临时装片保存备用,每一片叶下表面2个装片。用显微数码相机进行图片拍摄,每个装片观察4个视野,用Image J软件测量;气孔密度(SD)和表皮细胞密度的观测在100×镜下进行,气孔长(保卫细胞长度,SL)、气孔宽(垂直于气孔长的保卫细胞最大宽度,SW)和气孔面积(SA)的观测在400×镜下进行,每片叶下表面气孔形态特征的观测不少于60个气孔。依据测定数据计算气孔指数[SI,气孔密度/(气孔密度+表皮细胞密度)×100%][7],表征气孔数占叶表面细胞总数的百分比。
1.2.3 叶片光合能力及δ13C值在剩余4株中与叶气体动力学参数测定相同位置选择叶片,进行CO2响应曲线测定。CO2由LI-6400光合仪携带的小气瓶提供,其设置梯度为400、300、200、100、50、200、400、600、800、1 000、1 200、1 500和1 800 mol·mol-1,测定前先在饱和光强下诱导30 min,测定过程中维持光强1 200 μmol·m-2·s-1,叶温大约25 ℃,相对湿度50%~60%。利用Sharkey等的方法[18]求得最大羧化速率(Vcmax)和最大电子传递速率(Jmax),连同大气CO2浓度下(400 μmol·mol-1)的光合速率(Aair)来表征植物叶片的光合能力。测定结束后采集叶片扫描测量叶面积,然后在75 ℃下烘干至恒重,称其干重,叶干重与叶面积之比即为比叶重(LMA)。烘干样粉碎过筛后,用MAT-251型质谱仪(美国Finnigan公司)测定叶片的13C/12C比,并以PDB(Pee Dee Belemnite)为标准计算叶片δ13C,质谱仪分析精度为±0.2‰。所有测定重复4次。质谱仪测定的叶片δ13C可间接反映C3植物水分利用效率的大小,该值越大,叶长期水分利用效率越大。
1.3 数据分析
1.3.1 气孔导度动力学模型及参数采用常用的指数曲线对气孔的开放与关闭过程进行模拟[18]:
气孔开放过程:gs=gsmax+(gsmin-gsmax)e-t/Ki
(1)
气孔关闭过程:gs=gsmin+(gsmax-gsmin)e-t/Kd
(2)
其中:gs为拟合的气孔导度,gsmin与gsmax分别为最小和最大稳态gs值,Ki和Kd分别为气孔开放和气孔关闭过程中的时间常数(即达到gs变异幅度63%所需时间),gsmax-gsmin为气孔开放和气孔关闭过程中gs的变化幅度(Δgsi和Δgsd),e为欧拉常数。用Vialet-Chabrand等提供的Excel菜单[19]对气孔动力学过程进行模拟。
1.3.2 统计分析用SPSS 20.0软件进行统计分析。处理之间gs动力学参数、气孔解剖结构及叶光合能力、叶δ13C值之间的差异显著性用单因素方差分析,若单因素方差分析达到显著后,用Tukey法进行处理之间差异显著性的多重比较(α=0.05)。
2 结果与分析
2.1 不同演替时期树种光诱导的气孔动力学
图1显示,3种树种光诱导的气孔导度动力学曲线变化趋势较相似;气孔开放过程中山杨和白桦气孔导度增加速率明显超过辽东栎,且达到最大值后存在一定程度的下降,而辽东栎这种过饱和现象较弱;辽东栎气孔关闭最先达到稳态,其次是白桦,而山杨最晚达到稳态。
进一步对图1气孔导度动力学过程进行模拟获得相应参数(图2)。其中,在弱光→强光诱导的气孔开放过程中,3个树种的时间常数(Ki)和气孔导度响应幅度(Δgsi)均存在显著差异,具体表现为:山杨和白桦的Ki显著低于辽东栎,但Δgsi显著大于辽东栎,而山杨和白桦间的Ki及Δgsi均无显著差异。在强光→弱光诱导的气孔关闭过程中,山杨和白桦气孔关闭的时间常数(Kd)均显著大于辽东栎,且山杨和白桦间无显著差异;山杨的气孔导度响应幅度(Δgsd)显著大于白桦和辽东栎,后两者无显著差异。山杨和白桦的Kd/Ki均接近1,并显著高于辽东栎,表明它们的气孔开放与关闭的时间相当,呈现出近似对称的响应;而辽东栎Kd/Ki远小于1,表明气孔关闭时间远快于气孔开放时间,呈现出明显的不对称响应。
2.2 不同演替时期树种的叶气孔解剖特征
3种树种叶片均只有下表面具有气孔。从表1可知,山杨和白桦的气孔密度显著低于辽东栎,两者的气孔密度仅分别为辽东栎的19.7%和32.7%;3种树种气孔长表现为山杨>白桦>辽东栎,且三者之间均存在显著差异;3种树种气孔宽度和气孔面积均表现为山杨和白桦显著大于辽东栎,且山杨和白桦之间无显著差异;3种树种气孔指数则表现为山杨<白桦<辽东栎,且三者之间均存在显著差异。以上结果说明演替后期种辽东栎表现出气孔密度大、气孔小和气孔指数大的特点。
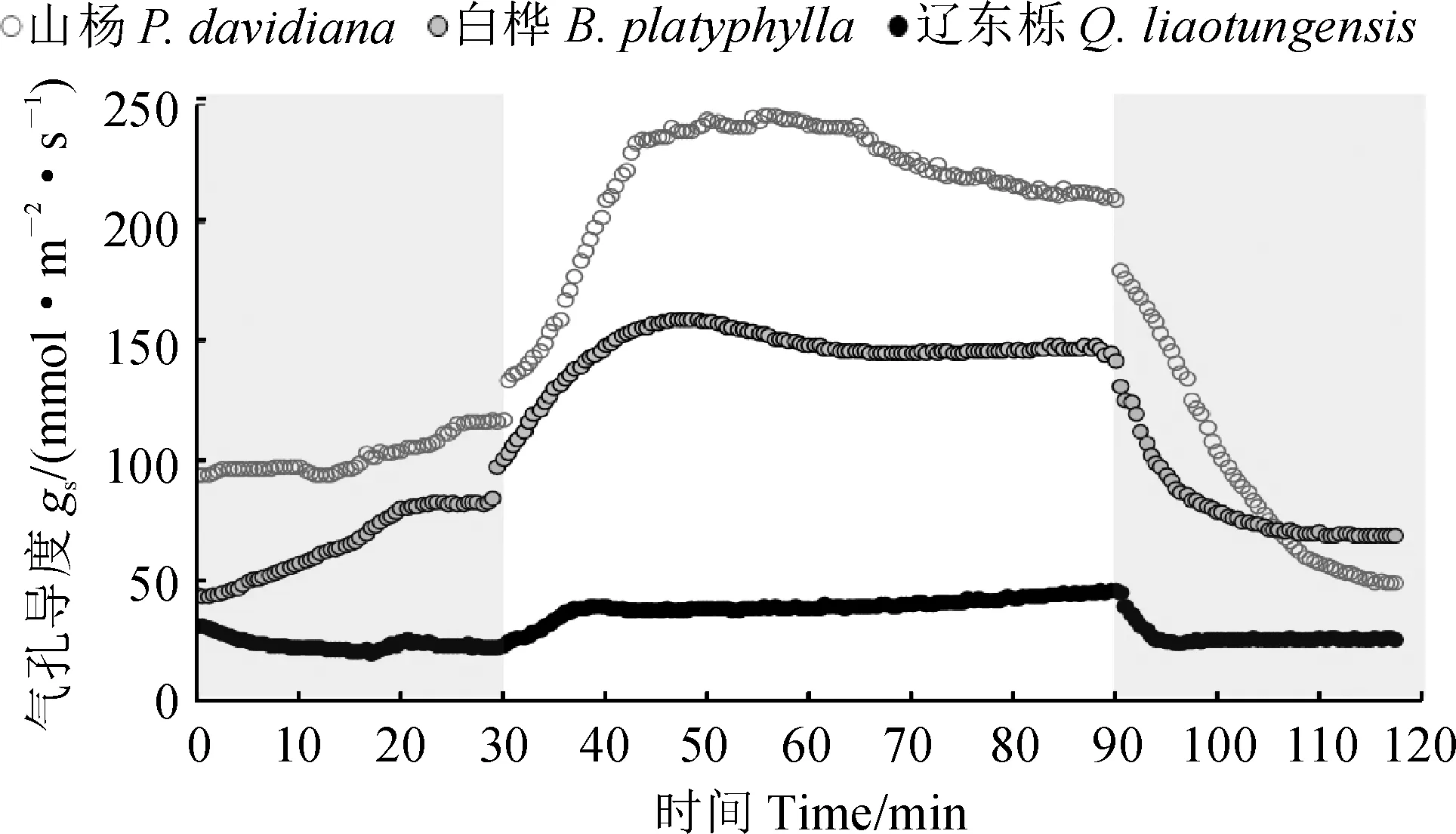
灰色部分为100 mol·m-2·s-1的弱光,白色部分为1 200 mol·m-2·s-1的强光图1 3种树种光诱导的气孔导度动力学曲线Grey and white color correspond to 100 and 1 200 mol·m-2·s-1 PPFD, respectivelyFig.1 Light-induced stomatal conductance dynamics in three typical species
2.3 不同演替时期树种的叶光合能力与长期水分利用效率
表2显示,3种树种的叶面积并无显著差异;辽东栎的比叶质量最高并显著高于白桦,但与山杨之间无显著差异,而山杨的比叶质量与白桦之间也无显著差异。演替早期种山杨和白桦的光合速率、最大羧化效率和最大电子传递速率显著高于演替后期种辽东栎,且山杨的最大光合速率与最大电子传递速率显著高于白桦。3种树种的叶δ13C值之间均存在显著性差异,表现为山杨>白桦>辽东栎。以上结果提示相对于演替后期种辽东栎,演替早期种山杨和白桦具有更高的光合能力和长期水分利用效率。

不同小写字母代表处理间在0.05水平下差异显著,下同图2 3种树种气孔开放和气孔关闭过程中光诱导的气孔导度动力学参数The different normal letters indicate significant difference among treatments at 0.05 level. The same as below.Fig.2 Dynamic parameters of stomatal conductance during light-induced stomatal opening and closing in three species

表1 3种树种叶的气孔解剖特征

表2 3种树种叶光合能力与δ13C值
3 讨 论
3.1 不同演替时期树种光诱导的气孔导度动力学参数间差异
本研究结果表明:在光诱导的气孔开放过程中,演替早期种山杨和白桦气孔开放明显快于演替后期种辽东栎,而气孔关闭过程则相反,演替后期种辽东栎的气孔关闭更快。一般认为,气孔的快速开放有助于叶快速达到最大光合速率,从而减轻气孔开放过程中气孔导度对光合作用的限制,而气孔关闭过程中快速的气孔关闭则有助于保持水分,从而减轻碳摄取降低时水分的无效消耗[3,6,10]。本研究中,演替早期种山杨和白桦一般生长在山坡底部,土壤水分条件较好且具有高的光合速率,因而快速的气孔开放反应有助于减轻气孔开放过程中的气孔限制,同时一定程度上补偿了演替早期种高蒸腾导致的水力梯度增大的缺陷[4],反映了演替早期种对演替早期变化的光环境的适应性。演替后期种辽东栎一般生长在水分限制的山坡上,因而快速的气孔关闭有助于维持水分。
植物气孔开放和气孔关闭的速率不同,一些植物气孔关闭快于气孔开放,如在McAusland等研究[3]的15种植物中,有7种的Kd显著小于Ki,在禾本科植物水稻[6]和小麦[10]也发现气孔关闭比气孔开放快,这种不对称性可能与这些植物水分保持优先于碳同化的策略有关。演替早期种山杨和白桦Kd/Ki值接近于1,气孔开放与气孔关闭快慢相近,可能与其生境中水分相对充足,植物优先碳同化来维持高生长速率有关,辽东栎气孔低的Kd/Ki值表明其在受水分限制的植被演替过程中,选择了保水优先于CO2摄取的策略,这种策略有助于增加其对长期水分亏缺的适应。
3.2 不同演替时期树种气孔动力学参数和气孔解剖特征之间的关系
由于研究的物种少(n=3),我们无法建立物种间气孔动力学参数与气孔解剖特征间的相关关系。前人研究表明:气孔密度大、气孔小及气孔指数高的物种其气孔反应更快[4,17]。从本研究看,演替后期种辽东栎气孔密度大、气孔小且气孔指数高,但其气孔开放最慢。其次,山杨和白桦的气孔长和气孔指数也具有一定差异,但二者气孔开放和气孔关闭快慢并无显著差异。因而我们认为,气孔特征之间的差异可能不是影响3种物种间气孔动力学快慢的主要因素,其他因素如副卫细胞数量和大小、叶生物化学、离子通道相关基因表达等可能影响更大[2,20]。
3.3 不同演替时期树种气孔动力学参数与水分利用效率间关系
气孔动力学研究显示:演替早期种山杨和白桦的气孔开放快于辽东栎,而气孔关闭过程中辽东栎的气孔关闭更快。演替早期种山杨和白桦同时具有更高的光合能力。快速的气孔开放有助于植物利用光斑,从而最大化光合作用[21],但在小麦上的研究表明稳态光合能力与气孔开放和关闭的时间常数呈正相关,即稳态光合能力越强,气孔开放越慢,稳态光合能力与气孔开闭快慢之间存在一种妥协关系[10]。本研究中演替早期种山杨和白桦快速的气孔开放可能有助于其取得更高的光合能力,但其慢速的气孔关闭则增加了气孔关闭过程中的蒸腾耗水,不利于水分利用效率的提高。叶片长期水分利用效率与气孔开放与关闭过程中光合速率和蒸腾速率变化均有关,尽管山杨和白桦气孔关闭过程中失水增加,但气孔开放过程中光合增加,导致其具有更高的水分利用效率。一些研究认为:快速的气孔关闭有助于保持水分,从而提高叶器官水平水分利用效率[3,6,10],但从本研究看,辽东栎快速的气孔关闭反应并未增加其水分利用效率,相反其长期水分利用效率低于山杨和白桦,原因可能与辽东栎慢速的气孔开放限制了其光合速率有关。
植物叶长期水分利用效率不仅与动态光强下气孔的反应有关,与稳态光下气孔的反应亦有关。在火炬松 (Pinustaeda)上的研究表明:动态光下慢速的气孔导度响应对总的日蒸腾速率、碳同化及水分利用效率影响不大[22],说明稳态光下气孔的反应可能更重要。此外,叶水分利用效率也受气孔对其他因素,如土壤水分、温度、蒸汽压亏缺和风等敏感性的影响。因此,今后需要通过气孔对环境因子的综合响应研究,以揭示3种植物气孔调节和叶水分利用效率差异的生理机制。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
辽宁省博物馆馆刊(2020年0期)2020-08-13
绿色科技(2019年19期)2019-11-26
东坡赤壁诗词(2019年3期)2019-07-05
中国边疆民族研究(2016年0期)2016-12-18
黑龙江生态工程职业学院学报(2013年4期)2013-11-20
陕西林业科技(2010年2期)2010-08-15
