
修剪措施对紫叶紫薇花蕾中内源激素和碳氮营养含量的影响
2022-01-15 05:35:38罗雪梦王晓明曾慧杰李永欣
西北植物学报 2021年11期
罗雪梦,王晓明,曾慧杰,李永欣,唐 丽
(1 中南林业科技大学 林学院,长沙 410004;2 湖南省林业科学院,长沙 410004)
紫薇(LagerstroemiaindicaL.)属于千屈菜科(Lythraceae)紫薇属(Lagerstroemia)植物,是中国夏季重要的园林观赏树种。大量关于果树生长的报道中,常采用修剪措施以调整树枝结构、改变树体营养分配、增加果实产量等[1-2]。关于紫薇采用修剪措施的报道已有很多,如剪除第一次开花的花穗实现北京地区紫薇花期持续至国庆节前后[3],通过修剪4/5当年生枝与施肥结合实现厦门地区紫薇盛花天数显著增加[4],达到实现二次开花、花期延长的目的。还有学者研究紫薇一年多次花开的技术措施,如沙飞等[5]研究证明紫薇在夏季采用轮次修剪处理后能够获得一年花开四度的最佳状态。
修剪措施对不同植株不同器官内的生长激素会产生不同程度的影响,通过改变内源激素比例促进或延迟成花进程[6]。苏曼琳等[1]发现文冠果(XanthocerassorbifoliaBunge)经不同程度短截后不同部位芽内源激素含量有所变化,脱落酸(ABA)含量下降,而赤霉素(GA3)、吲哚乙酸(IAA)和玉米素(ZT)含量均上升。艾沙江·买买提等[7]研究‘富士’苹果(Malus×domesticaBorkh.)枝条不同方式修剪后芽内源激素平衡,发现ZR和GA3含量显著提高,促进萌发成枝。修剪后植株各部分之间建立新的营养代谢关系,碳素主要供给生殖生长,氮素主要供给营养生长,如红花玉兰(Magnoliawufengensis)不同程度地夏季修剪均能提高可溶性蛋白和可溶性糖含量[8],不同程度短截后薄壳山核桃(CaryaillinoensisK.Koch)新枝叶片及枝条的碳氮比值随短截程度的增大而提高[9]。紫薇夏剪是目前重要的花期调控措施和生产实践经验,目前关于紫薇修剪的报道基本集中在处理后树势生长和开花特征等方面,鲜见紫薇修剪后激素水平和营养代谢变化的有关报道。
紫叶紫薇(purple-leafLagerstroemiaindica)枝干直立性强,花色大红色,成熟叶黑紫色至紫红色,是极具观赏价值的选育良种。本研究通过对紫叶紫薇主花枝进行重度修剪,探究修剪措施对花芽内源激素和碳氮代谢的影响,旨在为紫叶紫薇花期调控实践提供进一步理论依据,以支持紫薇修剪技术的实践应用与推广。
1 材料和方法
1.1 试验地概况
田间试验地位于湖南省长沙市雨花区湖南省林业科学院林木新品种创制中试基地,地处113°01′E,28°06′N,属低山丘陵地貌,地势较平缓,坡度15~20°。属亚热带季风性湿润气候,年均气温17 ℃,年均降雨量1 400 mm,年均日照时数1 500 h。试验地土壤为四纪酸性红壤,pH 4.0~5.5。
1.2 材料与处理
试验材料为5年生紫叶紫薇品种‘丹红紫叶’(Lagerstroemiaindica‘Ebony Embers’)。平均树高约3 m,树势中等,长势良好。选取生长势一致的‘丹红紫叶’设2个处理,一是每植株的双杈花枝的其中一个枝条进行重度修剪(剪去当年生枝长度的2/3),二是以植株不进行修剪为对照(CK)。每个处理3株,单株小区,3次重复。修剪处理于夏季一次花末花期进行,修剪后常规养护管理。
1.3 样品采集与处理
对照植株于花芽膨大期、花蕾成熟期、初花期、盛花期、末花期各采样一次,处理植株枝条于修剪后当年二次开花同样的花发育阶段各采样一次;在修剪处理枝条及对照植株上随机摘取顶端花蕾,用于内源激素含量测定的样品液氮速冻后存于-75 ℃冰箱中待测,用于可溶性糖、淀粉、全氮含量测定的样品在105 ℃烘箱中杀青30 min,80 ℃继续烘干48 h,粉碎机研磨过80目筛,装入密封袋待测。
1.4 测定指标及方法
1.4.1 内源激素含量称取鲜样0.5 g,液氮研磨为粉末状,激素的分离纯化参照饶丹丹[10]的方法进行,采用高效液相色谱法(HPLC)测定样品中赤霉素(GA3)、吲哚乙酸(IAA)、脱落酸(ABA)、玉米素核苷(ZR)的含量,高效液相色谱仪为岛津LC-20AT,色谱柱为Hypersil BDS C18色谱柱,流动相为甲醇-0.75%冰乙酸水溶液,色谱柱温度为30 ℃,进样量为10 μL,检测波长为254 nm,流速为1.0 mL·min-1。
1.4.2 可溶性糖和淀粉含量称取样品粉末0.1 g,采用蒽酮比色法测定样品中可溶性糖和淀粉的吸光度,再根据葡萄糖标准曲线计算可溶性糖和淀粉含量(mg·g-1)[11],并计算非结构性碳水化合物含量(可溶性糖含量+淀粉含量)。
1.4.3 全氮含量称取样品粉末0.2 g,采用硫酸-双氧水消解,凯氏法测定样品中全氮含量[12],并计算碳氮比(C/N,非结构性碳水化合物含量/全氮含量)。
1.5 数据分析
采用Excel2010进行数据统计与作图,SPSS25进行数据分析。
2 结果与分析
2.1 修剪处理对紫叶紫薇花蕾内源激素含量的影响
2.1.1 玉米素核苷(ZR)含量由图1,①可知,修剪处理紫叶紫薇在开花过程中花蕾的ZR含量均比同期不修剪对照显著提高,其在花蕾成熟期ZR含量较对照显著增加了263.98%(P<0.05)。修剪处理组花蕾中ZR含量在花芽膨大期至花蕾成熟期上升,之后逐渐下降,盛花期至末花期再次上升,但整个开花过程ZR含量变化并不显著(P>0.05)。对照组花蕾中ZR含量随生育期逐渐下降,从花芽膨大期至初花期变化不显著,至盛花期开始显著降低,盛花期和末花期ZR含量分别比初花期显著降低了57.82%和67.23%,而盛花期和末花期之间无显著差异(P>0.05)。说明紫叶紫薇经重度修剪后会显著提高花蕾中ZR水平,也明显改变了其在花芽发育过程中动态变化规律。
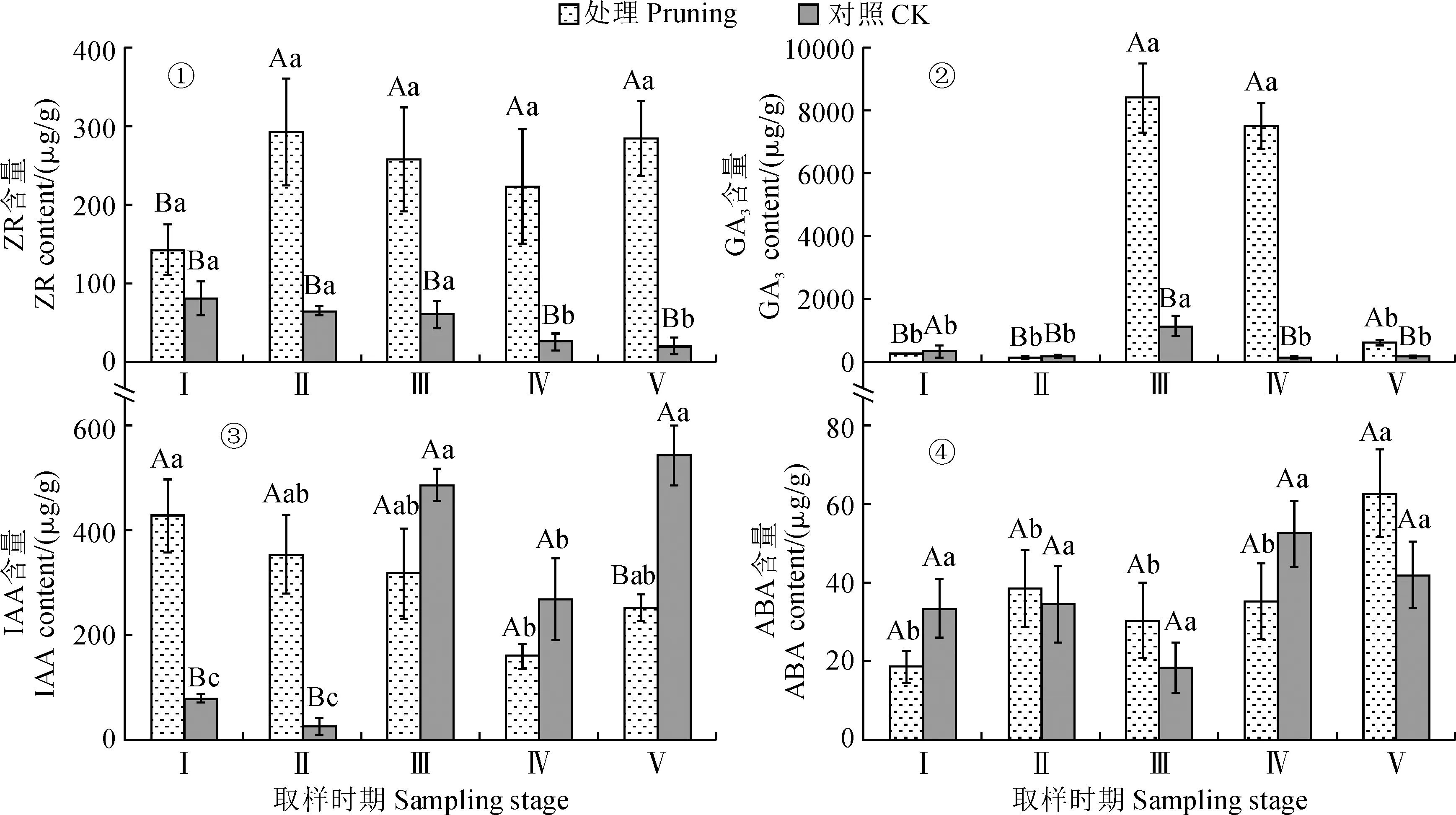
Ⅰ.花芽膨大期;Ⅱ.花蕾成熟期;Ⅲ.初花期;Ⅳ.盛花期;Ⅴ.末花期;处理内不同小写字母表示不同取样时期间在0.05水平存在显著性差异;同一取样时期内不同大写字母表示不同处理间在0.05水平存在显著性差异(P<0.05)。下同图1 修剪处理对紫叶紫薇花蕾内源激素含量的影响Ⅰ. Flower bud expansion period; Ⅱ. Flower bud maturity period; Ⅲ. Initial flowering period; Ⅳ. Full-flowering period; Ⅴ. Late flowering period;Different lowercase letters indicate a significant difference at the 0.05 level at different sampling time points for the same treatment, and different capital letters indicate a significant difference at the 0.05 level at same sampling time point for the different treatments(P<0.05). The same as belowFig.1 Effect of pruning treatment on endogenous hormone contents in flower buds of purple-leaf Lagerstroemia indica
2.1.2 赤霉素(GA3)含量紫叶紫薇花蕾GA3含量随开花过程大致呈现“降-升-降”的变化趋势(图1,②)。其中,修剪处理的紫叶紫薇花蕾GA3水平从花芽膨大期至花蕾成熟期降低约47%;然后在初花期迅速上升,并在初花期出现GA3含量峰值,此时较花蕾成熟期增加了约60倍;随后在盛花期GA3含量虽略降低,但仍维持在较高水平;末花期GA3含量比盛花期显著下降了91.81%(P<0.05)。对照组花蕾中GA3水平从花芽膨大期至花蕾成熟期同样降低约47%,然后在初花期比前一时期大幅显著增加约5倍,但初花期至盛花期GA3含量显著下降了88.62%(P<0.05),盛花期与末花期之间GA3含量变化不显著(P>0.05)。同时,在紫叶紫薇花芽分化、开花过程中,修剪处理组花蕾中GA3含量在花芽膨大期显著低于对照组,在初花期、盛花期、末花期GA3含量较对照组显著升高了2~56倍(P<0.05)。说明紫叶紫薇经重度修剪后会先显著降低花芽分化前期花蕾中GA3水平,而后显著提高开花时期花蕾中GA3水平。
2.1.3 吲哚乙酸(IAA)含量图1,③显示,修剪处理的紫叶紫薇花蕾IAA含量在花芽分化期显著高于不修剪对照4.4~12.8倍,其在初花期及盛花期低于对照组但差异不显著,其在末花期显著低于对照53.63%(P<0.05)。修剪处理组花蕾中IAA含量在花芽膨大期为最高值,之后在花蕾成熟期及初花期小幅度下降,其在初花期至盛花期显著降低49.84%(P<0.05),而在末花期比盛花期显著上升了58.01%。对照组花蕾中IAA含量随开花过程呈现“降-升-降-升”变化趋势,其在花蕾成熟期较花芽膨大期降低但未达到显著性水平,在初花期显著升高,而后在盛花期显著下降并处于中等水平,其在末花期再次显著升高(P<0.05)。说明紫叶紫薇经重度修剪后于花芽分化期提高花蕾中IAA水平,而在开花期降低花蕾中IAA水平。
2.1.4 脱落酸(ABA)含量由图1,④来看,修剪处理和不修剪对照紫叶紫薇花蕾ABA含量在开花过程中均大致表现为“升-降-升”的动态变化趋势。其中,修剪处理组花蕾中ABA含量变化从花芽膨大期至盛花期均较稳定,无显著性差异,其在末花期较盛花期显著增加78.11%(P<0.05)。对照组花蕾中ABA含量在花芽膨大期和花蕾成熟期小幅波动,其在初花期呈最低水平(18.30 μg·g-1),至盛花期又上升至52.48 μg·g-1,但变化并不显著(P>0.05)。同时,修剪处理的紫叶紫薇花蕾ABA含量在花芽膨大期低于对照组44.38%,在花蕾成熟期及初花期分别比对照提高10.28%和39.80%,至末花期高于对照组49.55%,但整个开花过程中修剪处理组与对照组之间的ABA含量差异不显著(P>0.05)。以上结果说明重度修剪对紫叶紫薇花蕾中ABA含量的影响不显著。
2.2 修剪处理对紫叶紫薇花蕾非结构性碳水化合物含量的影响
2.2.1 可溶性糖含量图2,①显示,紫叶紫薇植株修剪处理组花蕾中可溶性糖含量随花发育进程逐渐升高,且花蕾成熟期及初花期比花芽膨大期分别显著上升了20.39%、62.48%,其在初花期与盛花期间无显著差异且处于较高水平(12.3~12.8 mg·g-1),至末花期比盛花期显著降低38.31%(P<0.05)。紫叶紫薇对照组花蕾中可溶性糖含量随花发育进程持续上升,各发育时期比花芽膨大期含量显著上升了23.92%~106.81%(P<0.05)。修剪处理组花蕾中可溶性糖含量在花芽膨大期至盛花期比对照组高6.94%~17.46%,在花蕾成熟期与初花期比对照组显著增加,其却在末花期比对照组显著降低43.06%(P<0.05)。说明紫叶紫薇经重度修剪后花芽分化期及开花前期的花蕾中可溶性糖含量增加,花期末期的花蕾中可溶性糖含量降低。
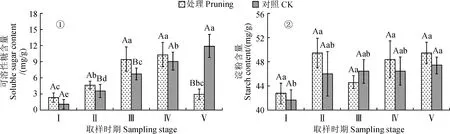
图2 修剪处理对紫叶紫薇花蕾可溶性糖和淀粉含量的影响Fig.2 Effect of pruning treatment on soluble sugar and starch contents in flower buds of purple-leaf L. indica
2.2.2 淀粉含量从图2,②可知,紫叶紫薇修剪处理组花蕾中淀粉含量从花芽膨大期至花蕾成熟期上升,在初花期下降,在盛花期与末花期再次逐渐升高;修剪处理组花蕾淀粉含量在花发育进程中处于42.75~49.46 mg·g-1之间,但其在各发育时期间均未表现显著性差异(P>0.05)。对照组花蕾淀粉含量在花发育进程中呈现逐渐上升趋势,其从花芽膨大期的41.64 mg·g-1上升到末花期的47.40 mg·g-1,显著上升了13.83%(P<0.05)。同时,修剪处理花蕾中的淀粉含量在初花期比不修剪对照降低3.96%,在其他时期增加2.66%~7.49%,整体花发育进程中处理组与对照组间差异均并不显著(P>0.05)。说明紫叶紫薇经重度修剪后花蕾中淀粉含量小幅度增加但受到影响并不显著。
2.3 修剪处理对紫叶紫薇花蕾全氮含量的影响
修剪处理组和对照组紫叶紫薇花蕾中全氮含量随着花芽分化和开花时期均逐渐降低(图3)。其中,修剪处理组花蕾中全氮含量从花芽膨大期至花蕾成熟期显著降低6.18%(P<0.05),而其在初花期至末花期未表现出显著性变化。对照组花蕾中全氮含量从花芽膨大期至末花期下降率在1.08%~2.99%,均不表现显著性差异(P>0.05)。同时,修剪处理组花蕾中全氮含量在花发育各阶段均高于对照组,比对照组分别增加了5.76%、1.54%、0.37%、1.91%和1.67%,但两者间未表现显著性差异(P>0.05)。说明紫叶紫薇经重度修剪后能够稍微提高花蕾中全氮含量,但增幅并不显著。
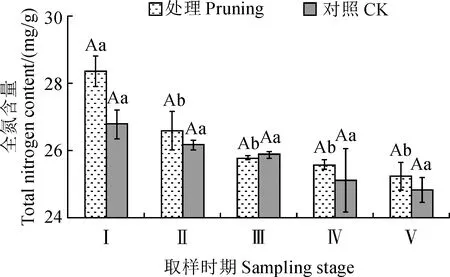
图3 修剪处理对紫叶紫薇花蕾全氮含量的影响Fig.3 Effect of pruning treatment on total nitrogen content in flower buds of purple-leaf L. indica
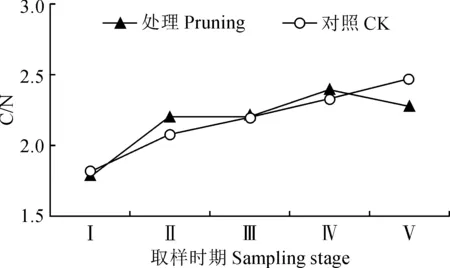
图4 修剪处理对紫叶紫薇花蕾C/N的影响Fig.4 Effect of pruning treatment on C/N in flower buds of purple-leaf L. indica
2.4 修剪处理对紫叶紫薇花蕾碳氮代谢的影响
修剪处理及未修剪的紫叶紫薇植株的花蕾中碳氮比总体上均呈现上升趋势(图4)。其中,修剪处理组的花蕾碳氮比值从花芽膨大期至花蕾成熟期上升较快,初花期比值波动小,然后在盛花期逐渐增大,最终在末花期逐渐下降至2.27。对照组的花蕾碳氮比值从花芽膨大期至末花期持续小幅度上升。同时,修剪处理的紫叶紫薇花蕾中碳氮比值在花芽分化期至盛花期高于不修剪对照,但其间碳氮比值差值小(0.030~0.126);其在末花期低于不修剪对照,碳氮比值相差约0.198。可见,紫叶紫薇经重度修剪会小幅度提高花芽分化期及开花前期花蕾中的碳氮比值。
3 结论与讨论
夏季修剪通过剪除部分老枝老叶及残花败果,可阻碍夏季高温多雨引发的叶部疾病,刺激腋芽抽发新枝及新生叶片的营养生长,促进内源激素重新分配及激素平衡调节,增强碳水化合物积累,为花发育进程提供能量。紫薇常采用夏季修剪的方式达到再度开花、延长花期的目的。从花的分化形成到再次开花的整个过程,始终发生着内源激素相互协调、碳水化合物代谢等各种生理生化反应。前人研究发现:巴拉多葡萄(Vitisvinifera‘Balado’)经花穗整穗修剪后测得花穗在盛花期及落花期细胞分裂素(CTK)含量提高,在葡萄幼果期ABA含量显著升高,盛花期和幼果初期IAA含量降低,盛花期至幼果期GA含量均降低。表明花穗整形可以减少巴拉多葡萄落花落果,增强座果率[13]。本研究发现,紫叶紫薇经过重度修剪后花蕾中ZR含量显著提高,并且自初花期至末花期GA3含量显著增加,说明紫叶紫薇修剪后使得植株再次成花、花期延长可能与ZR及GA3含量上升有关。李建军等[14]也发现忍冬(LonicerajaponicaThunb.)大白期时间延长与花蕾ZR水平升高有关。杨晓红等[15]表示植物组织处于衰老状态时,内源细胞分裂素水平会下降以减少水分的消耗,寿命长的花比寿命短的花具有更高的ZR水平。因此高含量ZR可能有利于水分的快速吸收,使植株花期延长。
关于GA诱导开花的理论认为GA浓度越高,表现出越大的促进作用,不存在超最适浓度的抑制作用[16],因此可能修剪处理后的紫叶紫薇花蕾GA3含量在开花期升高促进了植株花期延长。有学者发现短截后从根部运输而来的养分使枝条剪口下部的芽GA3含量大幅度提高[7],大量研究表明GA可以向上或向下进行双向运输[17],本研究中GA3水平的上升可能是因为枝条中上部被剪除后GA通过韧皮部积累于侧芽中,根部合成的GA通过木质部不断输送,使二次花GA3累积量增加。孟雯雯[18]对高羊茅(FestucaarundinaceaSchreb.)进行不同修剪高度处理后发现其根系赤霉素含量与对照组相比显著增加。其次,本研究发现,紫叶紫薇经过重度修剪处理后IAA含量在花芽分化期显著高于对照,在初花期至末花期较对照降低,贾玥等[13]也发现葡萄花穗经整穗修剪后IAA含量在开花中后期显著低于对照,其他时期IAA含量较稳定。Nito等[19]在报道中认为IAA含量高低在同一花穗中的分布有所不同,枝条上部IAA水平更高,中部及下部IAA水平较低,因此紫叶紫薇开花期IAA含量降低可能是IAA含量更高的部分被剪除而导致。另外,在本研究中,花芽分化期修剪组ABA含量低于对照组但无显著影响,据黄丛林等[20]研究发现葡萄果实中的ABA大部分来源于叶片,因此叶片可能是紫叶紫薇花蕾中ABA的主要来源,修剪处理使早期花芽内ABA来源不充足,ABA含量低于对照。重度修剪处理对紫叶紫薇花蕾中ABA含量整体情况的影响不显著,与刘仁道等[21]为促进甜樱桃(Prunus.aviumL.)生殖生长对直立生长的新梢进行拿枝处理的报道中情况相似,结果发现拿枝后ABA含量与对照差异不显著。在本研究中,对照组与修剪组在盛花期和末花期ABA含量均处于较高水平,ABA含量的增加可能与ABA具有增强代谢的作用有关,盛花期至末花期紫叶紫薇开始产生果实,需要更多的营养,ABA含量上升促进了代谢库活性[22]。有学者认为ABA在成花的不同阶段可能有不同作用[23],本研究的紫叶紫薇修剪处理组ABA含量在花芽分化期低于对照组而在末花期高于对照组,推测高水平ABA可能与紫叶紫薇植株修剪后花期延长相关,但修剪后ABA含量在末花期升高对紫叶紫薇花期延长的影响效应有待进一步探究。
在花的形成与开放过程中,碳水化合物和氮代谢起着重要的作用。孙颖等[24]认为油茶在简化整形修剪模式下很大程度地提高了叶碳水化合物的累积,从而提高油茶产量与品质。本研究发现,紫叶紫薇花蕾中可溶性糖含量随开花进程逐渐上升,但变化量不大,这与高小俊等[25]对芒果(MangiferaindicalL.)短截后花芽内可溶性糖含量变化的研究结果类似。可溶性糖作为关键的能量来源和结构物质,其含量的增加可促进细胞液浓度,是花发育进程的保障。修剪处理组可溶性糖含量在花芽分化期高于对照组,可能是修剪处理解除了顶端优势,可溶性糖大多集中于附近侧芽,促进腋芽发育成花芽[26]。前人研究中关于番红花(CrocussativusL.)[27]、枣(ZiziphusjujubaMill.)[28]等在花芽分化期其可溶性糖含量增加,我们可以认为修剪处理使紫叶紫薇可溶性糖含量提高与紫叶紫薇再次成花相关,高可溶性糖含量有助于紫叶紫薇花芽分化。末花期修剪处理组可溶性糖含量显著低于对照组,田璐[29]同样发现忍冬花蕾可溶性糖含量随花芽分化进行逐渐上升,至花盛放时达最高值,开放后由于能量的大量释放糖含量开始降低,因此可以认为紫叶紫薇再度开放后末花期有较多果实产生,消耗了植株大部分养分导致糖含量快速下降。本试验中修剪组紫叶紫薇花蕾中淀粉含量先升高后下降,然后再次升高,与环剥处理下毛棉杜鹃(RhododendronmoulmainenseHook.)花芽分化过程叶淀粉含量变化规律相似[30]。紫叶紫薇初花期淀粉含量的降低趋势可能是因为淀粉在植株内作为能量补充的角色,并不直接被利用,因此淀粉水解为可溶性糖为生长代谢提供能量,淀粉累积量较低[25]。淀粉作为贮藏性营养物质,在后期成为主要的花发育能量提供者,因此开花中后期淀粉含量继续升高。
本研究中,紫叶紫薇修剪处理组与对照组的全氮含量表现为逐渐降低趋势,且修剪组全氮含量略高于对照组,与罗丽[31]短截处理川香核桃(Juglansregia‘Chuanxiang’)全氮含量变化类似。而张翔[9]的研究认为薄壳山核桃在不同程度短截处理后其全氮含量均低于对照,推测这与植物的试验地区、当地气候条件及植物品种关系密切。本试验中紫叶紫薇修剪组的全氮含量较对照组增幅并不显著,在夏季修剪对红花玉兰生理影响的研究中同样发现全氮及可溶性蛋白含量仅在一定程度上增加[8]。修剪处理使得全氮含量在花发育进程提高的响应特征有待进一步探究。
碳氮代谢是植株最基本的代谢活动,碳和氮的比例适当,营养充足,有利于花芽分化,开花也多。碳氮比协调对紫叶紫薇花发育生理过程起到推动作用。本试验结果表明,紫叶紫薇花蕾碳氮比值随花发育时间呈逐渐上升趋势,修剪组碳氮比值在花芽分化期及开花盛期小幅度高于对照组。碳氮比学说认为高碳氮比状态有利于花芽分化,反之则抑制花芽的形成。前人对短截处理后的薄壳山核桃的碳氮代谢研究也发现碳氮比提高,从而说明适度修剪处理可以促进薄壳山核桃成花[9]。夏黑葡萄(Vitisvinifera×Vitislabrusca‘Summer Black’)采用喷药处理促进二花的试验中,其可溶性糖含量增加,使得碳氮比增加,表明二次成花与碳氮比提高有密切关系[32]。因此,紫叶紫薇经重度修剪后碳氮比值高于对照可能与紫叶紫薇再次成花相关。在本研究中,紫叶紫薇修剪组碳氮比值在末花期下降,且比值低于对照组,原因可能是末花期气温降低及光照强度减弱,叶片逐渐衰老,植株光合能力受到影响进而非结构性碳水化合物积累下降;也可能是花果期即将结束,非结构性碳水化合物向其他储存器官转移为休眠期做准备[33]。
综上所述,内源激素的作用方式及碳氮代谢的协调程度与紫叶紫薇整个花发育进程的关系极其复杂,紫叶紫薇经过重度修剪处理后,花蕾中ZR、GA3含量显著升高,IAA含量先上升再下降,ABA含量变化较稳定;可溶性糖、淀粉、全氮含量增加;碳氮比值小幅度提高,然后在末花期比值降低。多种因素共同调控紫叶紫薇的二次成花进程。本试验只研究了紫叶紫薇花发育过程中的生理变化(内源激素和碳氮含量),尚未探讨内含物质变化的内因,有关影响紫叶紫薇花发育过程中这一系列变化的基因调节机制有待深入研究。另外,本研究为小区域试验,且品种单一,研究结果具有一定的局限性,还需要增加品种数量及植株数量进一步探究与充实。
猜你喜欢
气象水文海洋仪器(2022年2期)2022-07-08 09:48:48
农产品市场周刊(2022年4期)2022-05-21 15:15:28
农产品市场周刊(2022年4期)2022-03-23 22:34:20
作文小学中年级(2021年11期)2021-12-11 09:09:52
北京园林(2020年2期)2020-01-18 03:30:56
小学生优秀作文(低年级)(2018年11期)2018-11-14 02:08:08
大东方(2018年1期)2018-05-30 01:27:23
现代园艺(2017年13期)2018-01-19 02:28:02
花卉(2016年1期)2016-02-17 07:10:18
上海质量(2015年2期)2015-12-02 02:26:44
