
根土复合体材料的抗剪强度特性研究进展
2022-01-07 02:38陈飞施康钱乾罗特
有色金属科学与工程 2021年6期
陈飞, 施康, 钱乾, 罗特
(江西理工大学,a. 资源与环境工程学院; b.江西省矿业工程重点实验室,江西 赣州341000)
0 引 言
近年来, 随着我国基础设施建设的不断推进,由人类工程活动而产生的边坡显著增加。在对边坡的开挖中,不仅破坏了原有的植被覆盖层,而且对其土体结构造成了一定的破坏, 使得边坡的稳定性变差[1]。为了减少因边坡的失稳对人民生命财产造成的损害,对于边坡的治理就显得尤为重要。
传统的护坡方式(诸如挡土墙[2]、抗滑桩[3]、框锚结构[4]等)虽然对边坡能够起到及时加固和稳定坡体的目的,但由于这类护坡工程造价较高,而且破坏了当地的生态环境,使得工程所在地的本土植物难以恢复,从而导致人类工程建设与环境保护之间的矛盾越发严重[5]。 为了缓解这一矛盾,现阶段的生态护坡技术则是通过植物根系与土体的盘根错节来加固土体,起到强化边坡土体的抗剪强度,增强浅层土体稳定性的作用[6-7]。 另一方面,相比于无植被的坡体,在发生降雨的情况下,植物不仅能够起到一定的抗冲刷的作用,而且通过植物自身的蒸腾作用,可以增加土体的吸力,起到防止水土流失的作用[8-9]。
通过植被来维持边坡土体的稳定性至少可以追溯到我国的明朝,在1591 年,我国就有通过利用柳树来稳定堤防的记录, 而欧洲最早利用植被来护坡的记录是在19 世纪[10]。 根据相关文献的结果表明,植物护坡主要通过植物根系与土体之间的附着力以及根系之间的相互缠绕来加固边坡, 利用草本植物的加筋和木本植物的锚固作用实现对浅层土体的强化[11-13]。加筋作用主要表现为草本植物在浅层土体分布的细根能够显著的提升土体的黏聚力, 而黏聚力的提升能够增强加筋土体的强度,从而提高浅层边坡土体的稳定性。锚固作用则是木本植物通过根系将浅部欠稳定土体锚固到深部稳定土层中,从而对浅层土体起到强化作用。
为了进一步研究根土复合体的强化机理, 对于抗剪强度的研究是十分必要的, 大量研究表明,植物的种类,土体中根系的分布类型、根径大小、根数和土壤水分、干密度等因素都会对土体抗剪强度产生影响[14-17]。 另外由于植物根系与土壤的弹性模量不同,在土壤发生变形破坏时,先一步发生变形的土壤会带动根系发生变形破坏, 而根系为了抵抗这种破坏, 将土壤强加给自己的力分散到周边稳定土体中,进一步弱化土壤的变形应力,起到延缓土壤变形和增强土体强度的作用。 有学者通过直剪和三轴试验表明根土复合体能够有效减小侧向形变和提高抗剪强度[18]。
1 植物根系加强土体的抗剪强度特性
1.1 草本植物的抗剪强度特性
草本植物根系在边坡土体中分布相对较浅,通过在边坡表层土中盘结缠绕,对于土体能够起到一定的网兜作用,因此根系的存在不仅能够对浅层土体起到相对明显的加筋作用, 达到强化土体强度的效果,而且对于土体的变形能够起到一定的制约作用[19-20]。
郑启萍等认为草本植物根系对于土体内摩擦角的影响是微弱的, 但对于土体黏聚力的提升却是显而易见的[1]。 张兴玲等通过直剪摩擦试验表明含根土体的含水量与垂直压力保持一定时, 含根土体的内摩擦角变化幅度较小, 而其黏聚力却有着显著的增强[21]。 陈洁等表明土体干密度的增加对内摩擦角影响不大,却能够显著提高土体的黏聚力[16]。 夏振尧等通过试验发现,相比没有根系存在的无根土,具有紫花苜蓿和狗牙根的根系复合土体的黏聚力得到了显著的提高[22]。
1.1.1 根系含量对于土体强度的增强
杨亚川等通过复合体原型结构试样的直剪试验发现,复合体的内摩擦角与含根量相关性很小,而其黏聚力与含根量表现为正相关[23]。张谢东等以白三叶为试验对象,通过直剪试验,建立了植物根系的“锚固”公式[24]:
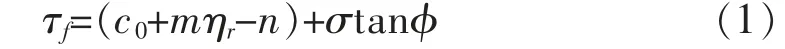
其中:τf为土壤抗剪强度(kPa);ηr为根的分布密度(kg/m3);m,n为强度作用系数,与根系特性有关;σ为剪切面法向应力(kPa);φ 为土的内摩擦角(°)。
该“锚固”公式表明,根系密度与土体的附加黏聚力成正相关关系,即土体中的根系越密,其对所在土体强度的提升越大。由于土体的黏聚力主要取决于土颗粒间的各种胶结作用,加上植物根系本身能够分泌有机胶结剂,其对于土颗粒之间的胶结具有一定的增强作用。 因而根系越密,其分泌的胶结物质更能有效地提升各个土颗粒间的结合强度,进而提高土体的黏聚力。另外,根系密度的增大,会导致根系与土颗粒接触面积的增大,从侧面也可以反映出根系对各个土颗粒的“网兜”作用会更加明显。 当土体承受荷载时,根系的“网兜”作用能够使根土复合体承受更大强度的荷载,进而对复合土体的强度产生一定的强化,由此对于土体抗剪强度的提升产生有利贡献。
1.1.2 植物浅层加筋与土壤密实度的关系
虎啸天等通过试验表明含有草本植物根系土体的黏聚力c值从表层土体向下呈逐渐降低的趋势[25]。从侧面验证了天然土体中植物的根系密度随着深度的增加呈现逐渐较小的趋势,也再次说明了植物根系的浅层加筋效应。单炜等通过三轴试验模拟植物根系的加筋效果,结果发现土体越密实,土体的加筋效果越明显[26]。
其原因为土体越密实, 根系与土壤的缠绕接触越紧密,当土壤发生剪切变形破坏时,根系更容易通过拉断产生抗拉强度来抵制这种破坏,而当土壤越松散时,根系则是通过被拉拔出来去抵抗这种变形破坏, 然而其产生的抗拔力是远小于抗拉断对于土体强度的贡献,因此,土体越紧密,植物的浅层加筋效果越明显。
1.1.3 根系分布形态对于抗剪强度的影响
孔纲强等通过开展高羊茅植物根系分别在水平、垂直、倾斜、相交及混合等分布形态下含根复合土的十字板剪切试验,结果表明,混合分布形态的根系增强最为明显[27]。 江锋等建立的复合土体力学模型,并推导出了在根的加筋作用下所增加的土体抗剪强度公式,其以狗牙根为研究对象,将大量的常规直剪试验的数据结果与公式得出的计算结果进行对比分析,发现土体和根—土复合体的抗剪强度随着含水量的增加而下降,而且斜根与直根对于根—土复合体的抗剪强度都会产生增强的效果, 但随着根条数的增加,斜根对于复合土体的增强作用更明显[28]。陈终达等以香草根为研究对象,通过室内直剪试验发现垂直加斜交等量布根的复合土体抗剪强度最高,而垂直加水平等量布根的复合土体抗剪强度最低,其他的布根形式介于上述二者之间,并表明当其复合体的根径达到某个阈值时,其抗剪强度达到最大值[29]。
由上述可得,与剪切面倾斜布置的根系对于抗剪强度的提升最大。这主要是由于倾斜的根系在进行直剪试验时, 能够有效地利用自身与土体的摩擦作用,通过根系与土颗粒之间摩擦力的分力来阻碍复合土体发生的剪切变形。而垂直布置的根系与土壤的摩擦力在抵抗剪切变形方向的分力为零,仅能通过根系自身的抗剪断来抵抗这种变形破坏。而且随着根数的增加,倾斜布置根系的优势会越来越明显,因此倾斜布置的根系对于抗剪强度的提升最高。
1.2 草本植物根系的抗拉特性
植物根系在土壤中的根系含量与根径大小在不同深度是不尽相同的,为了研究根系含量及根径的变化对于土体抗剪强度的影响,掌握根系含量以及根径大小随土体深度之间的变化函数对于量化其对于土体抗剪强度的贡献具有重要意义。
有研究表明,根系在土中的含量随着深度的增加而逐渐减小[30]。李建兴等以草本植物香根草、白喜草、狗牙根以及紫花苜蓿为研究对象,探讨其根系含量与土层深度之间的关系, 结果发现随着土壤深度的加深,根系的含量呈现以线性函数或指数函数的降低趋势[31]。刘昌义等通过研究草本植物与灌木植物单根抗拉力与根径之间的关系,发现这两者之间呈现幂函数关系,同时表明单根抗拉强度与根径之间表现为随根径增大,抗拉强度却减小的规律[32]。 Bischetti 等给出了根的抗拉强度与根径之间函数关系公式[33]:

式(2)中:α(MPa)和β(无量纲)是依赖于植物种类的经验常数;d为根径(mm)。
文伟等将野外挖取的狗牙根根系与海南地区红黏土随机混合,通过三轴试验发现抗拉强度与直径表现出幂函数的递减关系,而单根狗牙根的抗拉力与直径表现为线性函数的递增关系[34]。
根据上述学者的研究不难发现, 虽然植物根系的抗拉力随直径的增大而表现出线性增大的函数关系,但是其抗拉强度却表现为随根系直径的变化呈现反幂函数关系。 这主要是由于抗拉强度表达的是单位面积上的拉力,相比根径较大的根系而言,细小的根系具有更高的强度。而且根据Wu 氏的根土复合体力学模型,利用式(2)计算出的抗拉强度能够方便的得出根系对于复合土体的强度增量, 因此根系抗拉强度与根径函数的量化对于复合土体强度的研究贡献不言自明。
1.3 木本植物的抗剪强度特性
木本植物的粗深根通过扎进稳定的土层中,起到锚固浅层欠稳定土体的作用[35-36]。 肖宏彬等以草本植物香根草以及木本植物小叶女贞为研究对象,发现这两种植物根系都能提高根土复合体的抗剪强度,而且混交根土复合体的抗剪强度依旧符合库仑强度理论,并认定木本植物根系对于根土复合体的内摩擦角能够起到明显提高作用[37]。王月等为了研究根土复合体的抗剪强度特征, 选取小叶杨和白羊草为试验对象,发现土壤内摩擦角随着根长密度、根表面积的增大呈对数增长关系,并认为木本植物小叶杨增大了土壤的内摩擦角[38]。
1.3.1 根系的抗拉特性对于抗剪强度的影响
余芹芹等认为由于植物根系的存在,边坡土体的变形破坏不仅能够有效得到抑制,而且边坡土体的抗剪能力也能得到显著的增强, 并表明随着坡度的增加, 灌木根土复合体和素土的抗剪强度反而减小[39]。田佳等以贺兰山山杨、油松、青海云杉为研究对象,发现单根最大拉力与直径呈幂函数关系,表现为随着直径的增大而增大;单根抗拉强度、杨氏模量与直径成对数函数关系,随直径增大而减小[40]。张乔艳等发现当植物的生长情况保持一致时,根系的抗拉强度与根径的关系呈现为随着根径的增加总体呈下降趋势,并表明单根抗拉强度能够最大程度影响根土复合体抗剪特性,验证了细根和粗根搭配的根土复合体的抗剪强度高于单个细根或粗根组成的根土复合体[41]。
由此可得, 在根系抗拉强度与根径的关系上,草本植物与木本植物呈现出一致的规律性,即随着根径的增大,根系抗剪强度反而呈下降的趋势。 对于细根与粗根搭配的根土复合体的抗剪强度高于单独的细根或粗根组成的根土复合体这一现象,可从粗细根在复合土体扮演的角色进行解释。 在根土复合体中,较粗的根系主要起到支撑土体的作用。因为当土体发生剪切破坏时,较粗的根系能够有效阻碍土体的变形破坏, 其充当了降低复合土体所承受荷载的骨架作用。值得指出的是,由于较粗的根系与土颗粒间的接触面积相对较小,因此在土体发生变形破坏时,不能有效约束土颗粒产生的错动和脱离。 但就在这时,由于细根与土颗粒间的接触面积更大,细根对于土颗粒的网兜作用就开始逐步显现出来,能够起到稳定土颗粒的作用,所以细根的存在可以将土颗粒“固定”在粗根的周围,使其成为一个整体,对于土体的剪切破坏能够起到有效的阻碍作用。因此细根与粗根搭配的根土复合体对于土体的抗剪强度提升最大。
1.3.2 根的分布形态对抗剪强度的影响
为了研究根的分布形态对抗剪强度的影响, 吴鹏等通过试验变换根系与剪切面的夹角以及根系与土壤接触面积, 发现根系与剪切面斜交的复合土的抗剪强度大于根系与剪切面垂直的复合土[42]。 由此可得,根系与剪切面斜交对于土体抗剪强度的贡献更大。
丰田等表明当含水量保持一定,根系与剪切面是一个大于45°的锐角时, 复合土体的抗剪强度最高;且含水量越大抗剪强度峰值对应的角度也就越大,越接近垂直[43]。 曾红艳等以空心莲子草根系为试验对象,通过直剪试验发现, 随着根系与剪切方向倾角的增大,复合土体的抗剪强度不断增强,而且斜根比直根更易发生断裂[44]。
上述研究现象可以通过根系在剪切区与土体接触面积进行解释,斜根相比直根而言在剪切区与土体接触的面积更大,土体的剪应力分担到斜根根系上的应力更多,导致其更早地发挥其断裂时的拉断力。 周群华等通过数值模拟分析表明根系水平布置固坡效果最好[45]。 安然等基于岩土软件GeoStudio 进行二维有限元植被护坡模型分析,结果发现,在相同的附加凝聚力下,对于安全系数的提高效果,Tap 型(垂直根型)根系大于Plate 型(水平根型)的根系[46]。
为了方便众多学者在植物根系加强土体的抗剪强度特性的研究上有个直观的了解,将上述学者研究的植物类型和其得出的试验结果列表(见表1)进行归纳。 不难发现,大部分草本植物表现为仅增加黏聚力,对于土体的内摩擦角影响不大。另外,肖宏彬和王月2 位学者表达出草本植物增加黏聚力,木本植物增加内摩擦的观点。 至于是否适用所有的植物类型,尚需试验进行验证。
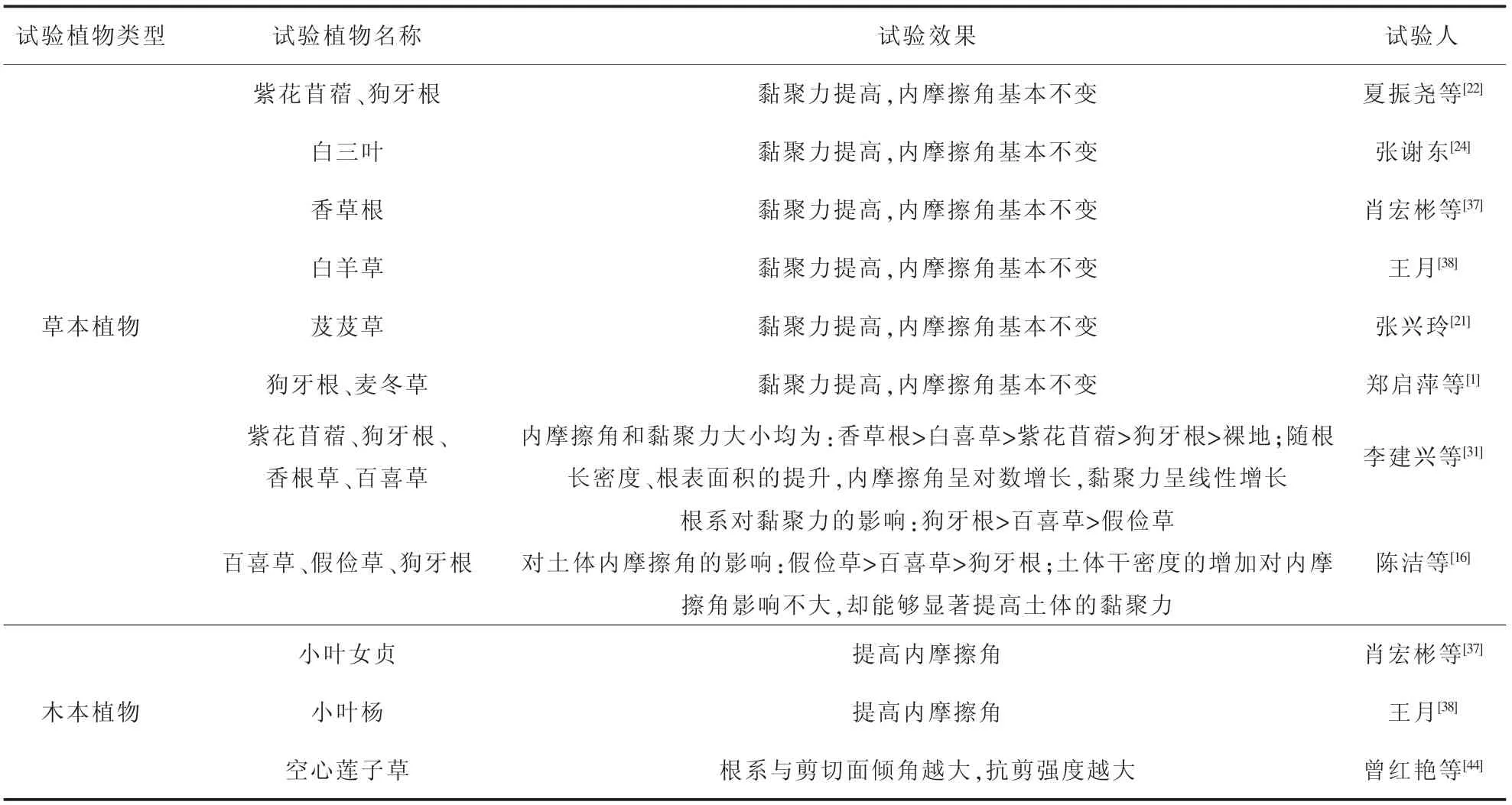
表1 植物根系对土体强度的作用Table 1 Effect of plant roots on soil strength
2 植物原状土与重塑土抗剪强度特性
在植物根土复合体抗剪强度试验中,以制备根土复合体原状试样做剪切试验居多[47]。赵丽兵等以豆科植物紫花苜蓿、草木樨、冰草和禾本科植物糜子这4种草本植物为研究对象进行野外剪切试验,发现对于土壤抗剪切强度的强化,紫花苜蓿根系的增强作用最大[48]。刘薇等考虑到在对植物根土复合体进行室内剪切试验时会造成土体震动,进而导致试验精度不高,于是设计了一种原位剪切试验装置用以解决该问题[49]。余芹芹等通过原位剪切试验验证在同一坡度下,根土复合体的抗剪力与剪切强度大于无根土体[39]。
2.1 重塑土与原状土强度的差异
由于原状土中的根系与土体间有着更为牢固的接触关系,在重塑土根土复合体与原状土复合土体的抗剪强度试验中,多表现为原状土根土复合体的抗剪强度更大。王元战等通过三轴试验发现在重塑加筋土的含根量达到原状土的4~6 倍时, 二者的破坏强度才基本相等[50]。 这说明在同等含根量的情况下,重塑土样的抗剪强度是小于原状土样的。刘子壮等通过研究发现在根径小于0.5 mm 的范围内,抗拉强度明显降低,这说明根径越细,单位横截面积根系抗拉能力越强,表明毛根对于根土复合体强度的提高发挥着极其重要的作用[51]。 张晓明等也验证在相同的植被、干密度、含水率以及同以垂直荷载下,原状土的剪切位移小于重塑土,原状土的抗剪强度大于重塑土[52]。 由此可得,原状土的根土复合体抗剪强度大于重塑土样的抗剪强度。 为了表明其中缘由,需要从多方面分析造成这种差异性的原因。
2.2 重塑土与原状土强度的差异性分析
重塑土与原状土的本质区别是在制作重塑土的过程中, 往重塑土里面添加的根系都是按照人为意愿修剪而放入土壤中的,与原状土相比,在重塑土样中添加的根系与土样的接触关系更为稀松, 根系与土样之间的咬合度更低, 并且最大限度地破坏了根系原本应有的结构。 而原状土则是最大限度地保留了根系自身的结构, 最为重要的是植物的细根也被保留下来。 当根土复合体的土壤发生剪切破坏时,由于细根与土壤颗粒紧密缠绕, 根系与土壤间的摩擦力较大, 导致其不易从土中拔出, 从而产生拉断破坏。 而粗根则易在土壤中被完整拔出,达不到自身所能发挥的抗拉强度, 因而粗根的抗拉强度对土体的加固贡献是不明显的[24]。而且在重塑土的制备中,多以添加粗根制备为主,这是导致重塑土的抗剪强度低于原状土的原因。
从活根的生物性质层面来看,由于细根具有活跃的生物学特征, 它在植物生长的过程中不断的生长,又不断地死亡,死去的根系为微生物提供了良好的生黏液质凝胶, 其能够将土壤颗粒有效地团聚起来,对土体的抗剪强度起到有利的贡献。 另一方面,活根本身也能分泌一定量的糖类、 有机酸等根际分泌物,这些分泌物对于土粒间的团聚也将起到较好的胶结作用[53]。而对于通过人为在土体中添加根系制备的重塑土样,其根系既无生物活性,也无原状土根系具备的大量毛细根系,从而造成了重塑土样的根土间结合强度较低,这也是导致重塑土样通过试验得出的加筋效果与原状土样产生较大差异的原因[54-55]。
3 植物加筋作用的抗剪强度理论基础
3.1 Wu 氏根土复合体力学模型
Wu 氏最早建立了根土复合体力学模型(见图1),其假定根系与剪切面垂直,当土体发生剪切变形破坏时,剪应力通过土体传递到根系上,根系为了抵抗土体的变形, 其所承受的力可以分解为平行于剪切面方向的水平应力trsinθ 和垂直于剪切面方向的法向应力trcosθ。 因此,根系对于土体的抗剪强度增量公式为[56]:
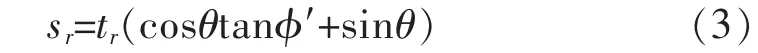
Wu 氏等通过敏感性分析发现 (cosθtanφ′+sinθ)对于θ 的值不敏感,并且当48°<θ<72°时,(cosθtanφ′+sinθ)接近于1.2[56]。 因此,式(3)可简化为:

根的面积比(简记RAR)是权衡根在土中含量的一个常用指标[30]。郭翬等表明在一定范围内,根土复合体的抗剪强度随着根面积比(RAR)的增大而增加,根系对土壤的加固作用也会越强[57]。根的总面积As与剪切区土壤的面积A的比值乘上单位面积根的拉力即为单位面积土中根产生的平均拉力,即Tr(As/A)=tr,因此,上式可转化为面积比下的抗剪强度增量:

图1 根系与剪切面垂直力学模型(Gray 等[58])Fig. 1 Vertical mechanical model of root and shear plane(Gray et al[58])

式(5)中:As为剪切面上所有根系截面积之和(cm2);A为 含 根 系 土 壤 截 面 积(cm2);As/A为 根 的 面 积 比(无量纲)。
式(5)表明土的抗剪强度增量只取决于根的平均抗拉强度Tr和根的面积比(RAR)。 因此当根系垂直于剪切面时,基于摩尔—库仑公式的复合土的抗剪强度公式为:
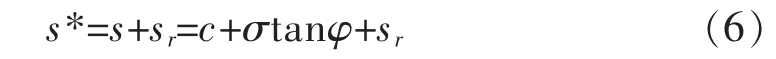
式(6)中:s为土的抗剪强度(kPa);σ 为剪切面上的法向应力(kPa)。
Gray 等基于Wu 氏的根土力学模型,推导出根系与剪切平面斜交时根系对于土的抗剪强度增量公式[58],其力学模型见图2。
根系与剪切面斜交时:

图2 根系与剪切面倾斜力学模型(Gray 等[58])Fig. 2 Mechanical model of root and shear plane inclination(Gray et al[58])
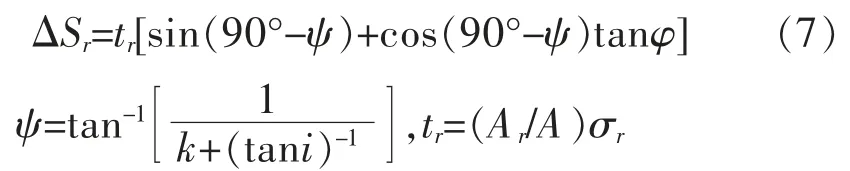
式(7)中:Sr为含根土的抗剪强度增量(Pa);tr为单位面积土中产生的抗拉强度(Pa);φ 为土的内摩擦角(°);ψ 为剪切变形角(°);i为根系延伸方向与剪切面的初始夹角(°);k为剪切变形率(k=x/z)(无量纲);z为剪切区的厚度(m);x为水平剪切位移(m);σr为剪切面上根系产生的拉伸应力(Pa)。
当根系与剪切面斜交时,复合土的抗剪强度公式为:
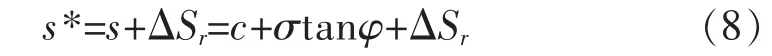
如果在面积为A的土体内,共有n个根,其中有m个垂直根,(n-m)个斜交根,根的抗拉力分别为T1,T2,…,Tn,垂直根的剪切变形角分别为θ1,θ2,…,θn,斜交根的延伸方向与剪切面的初始夹角为im+1,im+2,…,in,剪切变形比分别为km+1,km+2,…,kn,因此,根系加筋作用下的土体抗剪强度公式为[10]:
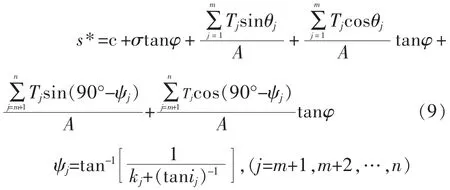
式(9)中:参数φ 可通过土的直剪试验获得;参数m、n和ij(j=m+1,…,n)可以通过截取不同的含根系土体纵剖面获得;θj(j=1,2,…,m)和kj(j=m+1,…,n)可以通过野外根系直剪试验获得,但实际上θj不易测得。对于根系抗拉力Ti,当根系被拉断时,则为根系的抗拉强度与横断面积的乘积,而当根系被拔出,则为根系与土体的摩擦力。
然而有学者认为Wu 氏模型高估了根系对于土壤的增强作用[59]。 Pollen 认为主要原因是Wu 氏假设所有的根部同时断裂,并且在断裂点发挥了全部的抗张强度[60]。 这与实际是不符的,因为根系在受力的时候不可能同时断裂,而是随根系抗拉强度的大小依次断裂。 根据抗拉强度与直径之间存在反幂函数的关系, 假设应力均分荷载到剪切区内的每个单根上,根系发生破坏时,根径最大的根最先发生断裂,而根径最小的根则最后发生断裂。再者根系也不可能完全发挥自身的抗拉强度,因为根系一旦被拉拔而与土体脱离,就仅仅只能发挥抗拔力对于土体的贡献,而抗拔力是小于根系通过断裂所发挥出的抗拉断力对于土体强度的贡献。
3.2 纤维束模型(FBM)
Pollen 应用广泛用于材料工业的纤维束连续断裂理论,提出根系渐进破坏模式。 该理论有效消除了Wu 氏模型由于假定根系同时断裂而高估土壤中根系提供的额外黏聚力。此模型假设根系在破坏的过程中, 根据每个根系的抗拉强度的不同而发生连续断裂,是比较符合根系实际的破坏情况。
对于该模型的基本原理,Thomas 做出如下叙述:含有n根平行纤维的纤维束受到载荷时,纤维束中的所有的纤维共同承担这个载荷,一旦载荷增加到某根纤维所称承受的最大抗拉强度时, 这根纤维就会断裂,而由这根纤维所承担的载荷就会被重新分配到剩下的(n-1)根完整的纤维上,致使每根纤维比初始状态承受更大的载荷,因此更有可能发生断裂。 如果这种载荷重分配造成更多的纤维发生断裂,重分配将会再一次发生,直到没有纤维发生断裂。 然后增加这个载荷,重复上述过程,直到所有纤维断裂[61]。
FBM 模型将载荷重分配到每根纤维后, 为了监测第n根纤维受到的载荷是否超过其最大抗拉强度,纤维模型的控制方程可写为:
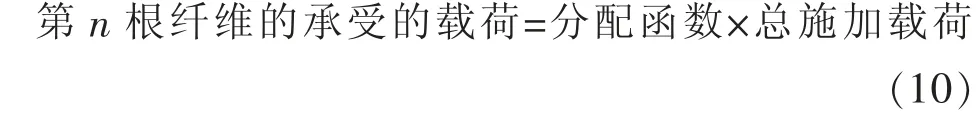
由式(10)可知为了监测第n根纤维承受的载荷是多少,分配函数的选取尤为重要。
Hidalgo 等提出了全局载荷分配GLS(Global Load Sharing) 和 本 地 载 荷 分 配LLS (Local Load Sharing)[62]。 GLS 假设来自断裂纤维的载荷在剩余的完整纤维之间重新分配,即重新分配的载荷与纤维的位置无关;LLS 假设载荷根据完整纤维与断裂纤维的接近程度进行重新分配,即重新分配的载荷与纤维的位置有关。 但由于纤维与纤维之间的位置难以确定,因此可采用GLS 模型确定分配函数。
FBM 最经典的应用是使用全局载荷分担(GLS),正如Pollen 等那样, 将应力均匀的重新分配到根束里的所有根系之间[63]。 Thomas 通过蒙特卡罗模拟发现,当施加到根束上的载荷被平均分配时,在不考虑根的直径或面积的情况下,最小的根首先断裂,最大的根最后断裂; 而当使每个根系承担相等的应力时,最大的根首先断裂,最小的根最后折断[61]。 由于抗拉强度与根径成反幂函数,因此当应力均分荷载到剪切区内的每个单根上,根系发生破坏时,根径最大的根最先发生断裂,而根径最小的根则最后发生断裂。 这与Thomas 模拟的结果一致。
然而根系在实际的破坏过程中, 土壤的性质、根系的直径、根系的生长方向和根系与土壤的摩擦都会影响根系的固土效果, 但FBM 模型并未考虑到这些因素。 因此, 为了解决上述问题,Schwarz 等提出了RBM 模型[64]。
3.3 根束模型(RBM)
RBM 模型是一个以位移控制加载过程的纤维束模型[65]。 本模型有效弥补了Wu 氏模型和FBM 模型的不足, 在计算根系对于土体抗剪强度的增强时,考虑了根的强度、根径、根的变形、根系与土的摩擦、根系土摩擦随土壤含水量和土壤类型的变化[66]。
RBM 模型认为根束的总抗拔力是位移的函数,可以表示为单根的抗拔力之和[63-64]。
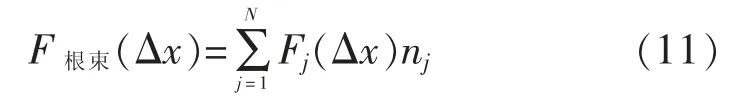
式(11)中:F根束(Δx)为整个根束的拔出力(N);Fj(Δx)为属于直径j类的单根拔出力(N);nj根束中直径j类束中的单根数量(个);N为直径分类的数量(个)。
整个根束拔出力与位移函数的变化可以采用微分方程来表示[63]:
式 (12) 中:Δxf和Δx0是2 种不同载荷下的位移(mm)。
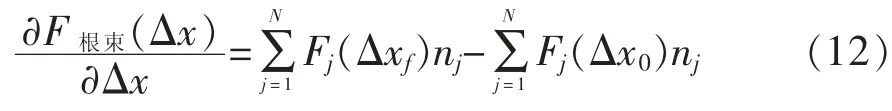
4 展 望
1)在植物护坡的研究方面,应当注意生物的多样性以及各种植物之间相互的竞争关系对于其在土壤强度贡献的制约性。 然而在研究植物的种类,根系的形状、根径、密度等对于土体抗剪强度的贡献方面,多采用以单一植物为研究对象,在多种植物类型混合种植对于土壤的强度增量方面研究不足,多种植物之间相互促进的补强关系少有研究。并且对于混合种植的植物,其得出的抗剪强度规律与单一植物测出的规律能否保持一致有待进一步的探索。
2)植物的生长受土壤和环境影响较大,同一种植物在不同季节、不同土壤之间的生长情况是不尽相同的, 这将导致根系的生长形态发生显著的变化。 因此研究植物在多个时期和地点的生长对于土壤强度的有利贡献是十分必要的,这对于植被护坡的可持续性具有重要意义,因此在以后的研究中应当着重强调植物多个时间段生长状况对于土体强化的影响。
3)根系的面积比(RAR)是衡量根系对于土的强度增量的一个重要指标。 大量研究表明随着RAR的增高,对于土壤抗剪强度的增加先增加,而后基本不变,即存在一个最优根系面积占比。 通过这一结论可以人为的布置植物的生长间距与数量,控制植物的根系在土壤中的占比尽量接近这个最优值,这样就能够最大限度地发挥出植物根系对于土体的加固潜能。因此,在今后的研究中要注意对该数据的获取与应用。
4)目前植被对于土体的强化,普遍认为通过增加土体的内聚力起到增强土体的作用。 而相关研究表明,对于不同类型的植物,其对土体的贡献不同,具体为草本植物对于土体强度的贡献表现为显著提高内聚力, 而木本植物则表现为通过提高土体的内摩擦角来增强土体的强度。 这将为采用草本与木本植物混种固土提供指导性意义,然而值得注意的是,得出这样结论的只是建立在以少量几种草本植物与木本植物混种为对象的基础上, 是否针对所有的草本植物和木本植物尚未可知, 今后可以通过试验进行验证。
猜你喜欢
发明与创新(2022年28期)2022-11-22
农业工程学报(2022年13期)2022-10-09
力学与实践(2022年3期)2022-07-02
大气科学学报(2022年2期)2022-05-12
聊城大学学报(自然科学版)(2022年3期)2022-02-14
作物学报(2022年3期)2022-01-22
今日自动化(2021年2期)2021-10-14
航空发动机(2020年3期)2020-07-24
珠江水运(2019年7期)2019-05-23
湖南大学学报·自然科学版(2014年6期)2014-09-27
