
大麦旗叶长和宽的QTL分析
2022-01-06 03:03姚琦馥魏佳泰李水琴
麦类作物学报 2021年11期
姚琦馥,魏佳泰,李水琴,刘 航,马 建
(1.铜仁学院农林工程与规划学院/贵州省梵净山地区生物多样性保护与利用重点实验室,贵州铜仁 554300; 2.四川农业大学小麦研究所,四川成都 611130)
大麦(HordeumvulgareL.,2n=14,HH)是世界上仅次于小麦、玉米和水稻的第四大重要谷类作物[1]。较小麦而言,大麦产量低、面筋含量低,且易倒伏[2],但大麦却含有种类丰富的营养物质,如高蛋白、高膳食纤维、高维生素、低脂肪、低糖等[3-4],因此大麦在保健方面具有很大的潜在价值。在种植面积有限的情况下,提高大麦单位面积产量意义重大。
植物叶片的形状、大小可以调节植物的生长发育,如株高、粒长、粒宽、分蘖、产量等[5-6],旗叶长和宽是旗叶大小的关键组成因素[7-9],是谷类作物提高产量的两个重要农艺性状,对产量具有重要作用[10-11]。旗叶长和宽均属于数量性状(quantitative trait locus,QTL),通常受多个基因控制,受环境影响也较大[12-14]。与旗叶长和宽相关的QTL已经在小麦、水稻等谷类作物中被广泛报道[15-17]。Farooq等[18]利用水稻材料IR64及其基因导入系对旗叶长和宽进行了QTL定位,在2号和4号染色体共定位到2个控制旗叶长的QTL;在1号和4号染色体上共定位到2个控制旗叶宽的QTL。Bing等[19]利用水稻品种珍汕97和IRAT109构建的包含180个株系的F9代重组自交系群体,在2、3、4、5、6、7和10号染色体上共检测到8个控制旗叶长的QTL,在4、5和6号染色体上共检测到5个控制旗叶宽的QTL。Ma等[11]于11个不同生态环境下在小麦2D和5B染色体上共检测到2个控制旗叶长的位点QFlw.sicau-2D和QFll.sicau-5B.3,在2D染色体上检测到1个控制旗叶宽的位点QFlw.sicau-2D,可解释4.09%~35.68%的表型变异。Liu等[6]对大麦旗叶形态相关的QTL进行了鉴定,但目前尚未见有人发现稳定表达的主效QTL,严重影响了通过调节旗叶形态来优化大麦株型结构,进而来实现大麦产量提升的目标。因此,本研究利用DArT标记对1个大麦重组自交系(RIL)群体进行分析,并基于多环境对该群体进行旗叶形态鉴定,以期获得控制旗叶长和宽的稳定表达的主效位点,为解析大麦旗叶形态遗传机制和分子辅助育种奠定基础。
1 材料与方法
1.1 试验材料
供试材料为1个通过单粒传方法获得的F10代大麦RIL群体。该群体包含125个株系,其亲本为经过多年多点鉴定表现稳定的J36528和BMJ89材料,均由四川农业大学小麦研究所保存。母本J36528旗叶较父本BMJ89长,而父本BMJ89旗叶较母本J36528宽。
1.2 试验方法
将RIL群体和亲本分别于2016、2017、2018和2019年种植在四川农业大学崇州试验基地(分别用2016CZ、2017CZ、2018CZ和2019CZ表示),于2018和2019年也种植在四川农业大学雅安试验田(分别用2018YA和2019YA表示)进行表型测定。试验田按照随机区组设计,包含2个重复。每个株系播种2行,行距0.3 m,行长1.5 m,株距0.1 m。试验材料均按单粒播种法进行播种,田间管理方式按常规管理方法进行。
1.3 旗叶长和宽表型数据的测定
于小麦开花期,每个株系选取长势良好且均一的单株用直尺进行旗叶长和宽的测定(不取每一行两边的单株,排除边际效应)。每个株系选取6个单株进行测量,同时计算出每个株系旗叶长和宽的平均值,用于后续分析。
1.4 遗传图谱的构建和QTL定位
按照Wenzl等[20]的方法利用DArT标记(https://www.diversityarrays.com/)扫描J365283/BMJ89 RIL群体125个株系中的92个单株和亲本,并利用已有SSR标记[21-23]对92个株系的群体进行分型[24]。对于存在共分离标记的位点,仅保留1个标记进行图谱构建。利用JointMap 4.0构建连锁图谱。利用IciMapping 4.0软件进行QTL检测,找到LOD>2.5的位点,同时计算出每个QTL的表型贡献率和加性效应。将在2个环境以上检测获得且表型贡献率大于10%的QTL视为主效稳定表达的位点。用Triticeae Multi-omics Center网站(http://202.194.139.32/blast/viroblast.php),将本研究鉴定到的以及前人检测到的QTL侧翼标记序列与大麦基因型‘Morex’参考基因组[25]进行比对(默认参数),获得QTL对应的物理位置。
1.5 数据分析
使用Excel 2019整理数据;使用SAS 9.4软件计算旗叶长和宽的最佳线性无偏预测(best linear unbiased prediction,BLUP)值和遗传力(H2);使用SPSS 17.0软件分析旗叶长和宽在不同环境及BLUP值的相关性;使用Origin 9.3软件绘制旗叶长和宽的频率分布图。
2 结果与分析
2.1 亲本及RIL群体旗叶长和宽的表型
对6个环境表型数据处理后发现,除了缺失数据外,J36528的旗叶长均极显著高于BMJ89,J36528的旗叶宽均极显著低于BMJ89(表1)。RIL群体在不同环境的旗叶长和旗叶宽均出现超亲分离现象,且旗叶长和宽均具有较高的遗传力,分别为0.93和0.82(表1),表明这2个性状主要受遗传因子影响,受环境影响较小。旗叶长和宽在每个环境下均呈现近似正态分布,符合数量遗传性状的特点,适合进行QTL定位(图1)。
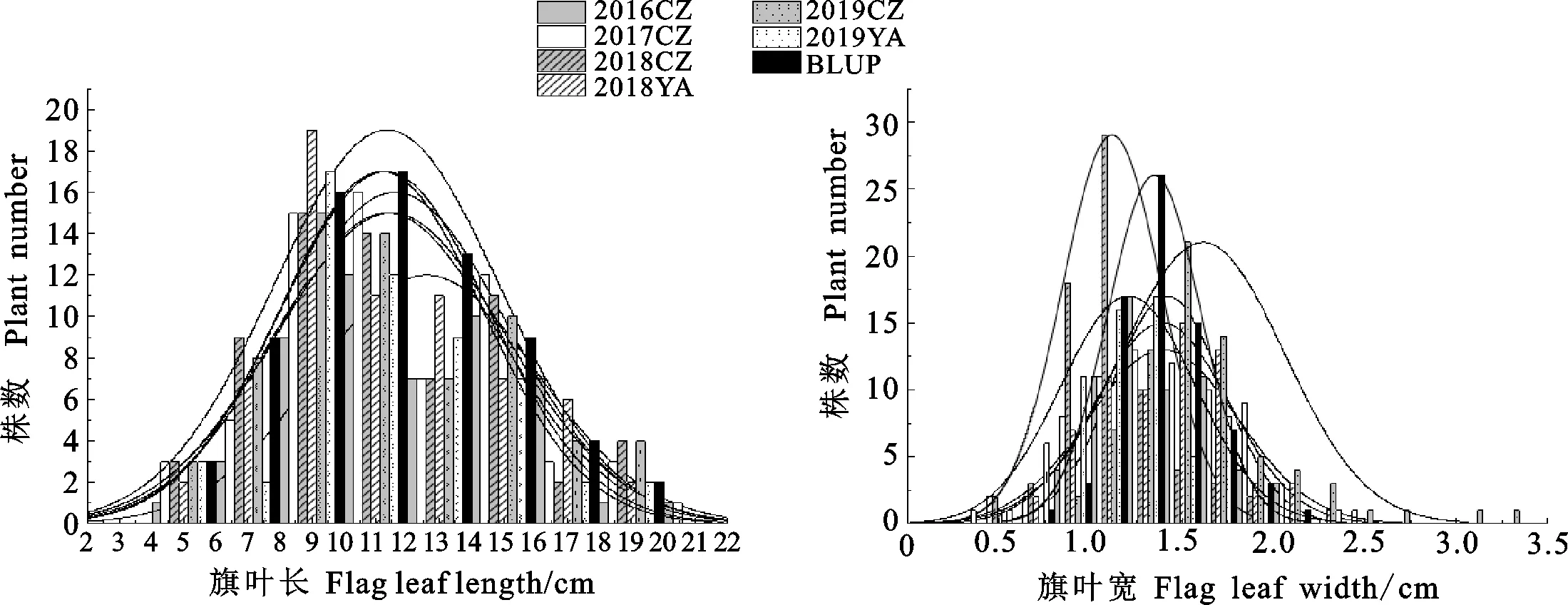
图1 不同环境下旗叶长和宽的表型频率分布Fig.1 Frequency distribution of flag leaf length and width of the RIL population in different environments

表1 不同环境下亲本及RIL群体旗叶长和宽的表型统计结果Table 1 Phenotypic statistics of flag leaf length and width of the parents and RIL population in different environments
2.2 旗叶长和宽在不同环境下的相关性
从表2可以看出,旗叶长和宽在不同环境及BLUP值之间均显著相关,旗叶长的相关系数介于0.975~0.997之间,而旗叶宽的相关系数介于0.955~0.994之间,进一步说明旗叶长宽受环境影响较小。
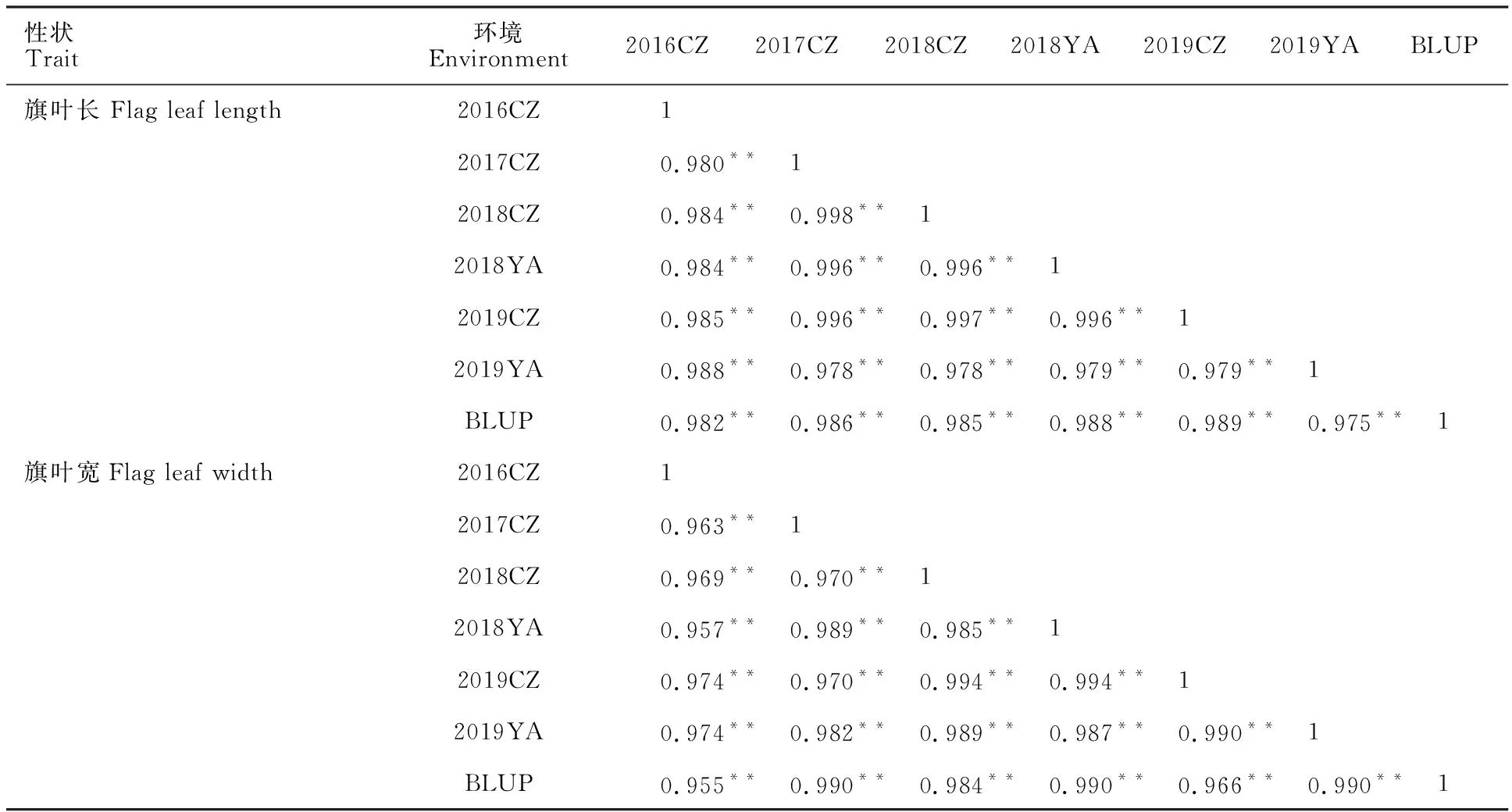
表2 旗叶长和宽在不同环境下的相关性Table 2 Correlation coefficients of flag leaf length and width in different environments
2.3 遗传图谱的构建
基于447个DArT和8个SSR标记构建大麦群体的遗传连锁图谱,分析结果(表3)显示,染色体上标记数量介于26~99之间,平均每条染色体有65个标记;图谱包含7个遗传连锁群,总长1 034.96 cM,每条染色体图谱长度介于94.96 cM ~218.80 cM,平均每条染色体图谱长度为147.85 cM。标记密度介于0.96~6.70 cM/标记之间,平均标记密度为2.27 cM/标记。
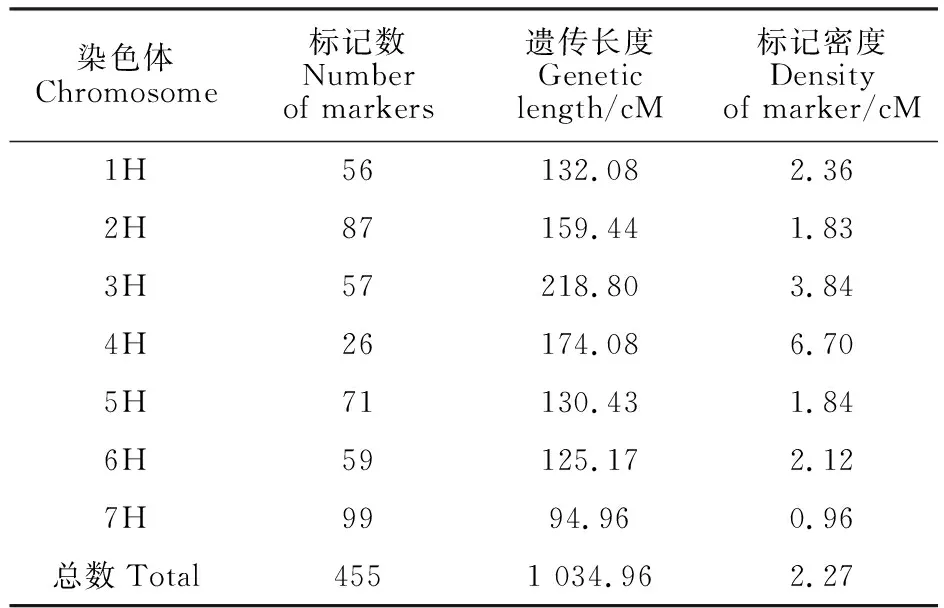
表3 构建的大麦遗传图谱中标记在染色体上的分布Table 3 Distribution of markers in the constructed barley genetic map
2.4 旗叶长和宽的QTL检测结果
基于遗传连锁图谱和测量获得的4年6个环境的表型数据,通过IciMapping 4.0软件共检测到12个控制旗叶长和宽的QTL(表4)。其中,检测到7个与旗叶长相关的QTL,分布在2H、5H、6H和7H染色体上。其中,5H染色体上的QFll.sicau-JB-5H.2在3个环境及BLUP值中被检测到,可解释12.02%~19.95%的表型变异,LOD值为3.14~3.61,为稳定表达的主效QTL,加性效应为正值,表明加性效应来自母本J36528。6H染色体上的QFll.sicau-JB-6H.1在2个环境及BLUP值中被检测到,可解释 16.69%~16.99%的表型变异,LOD值为 2.89~4.49,为稳定表达的主效QTL,加性效应为负值,表明其加性效应来自父本BMJ89。QFll.sicau-JB-2H.2和QFll.sicau-JB-7H在2个环境中被检测到,加性效应分别来自亲本J36528和BMJ89可分别解释9.81%~10.31%和 14.28%~ 14.45%的表型变异。其余3个QTL只在单一的环境中被检测到,可解释13.79%~ 28.84%的表型变异。
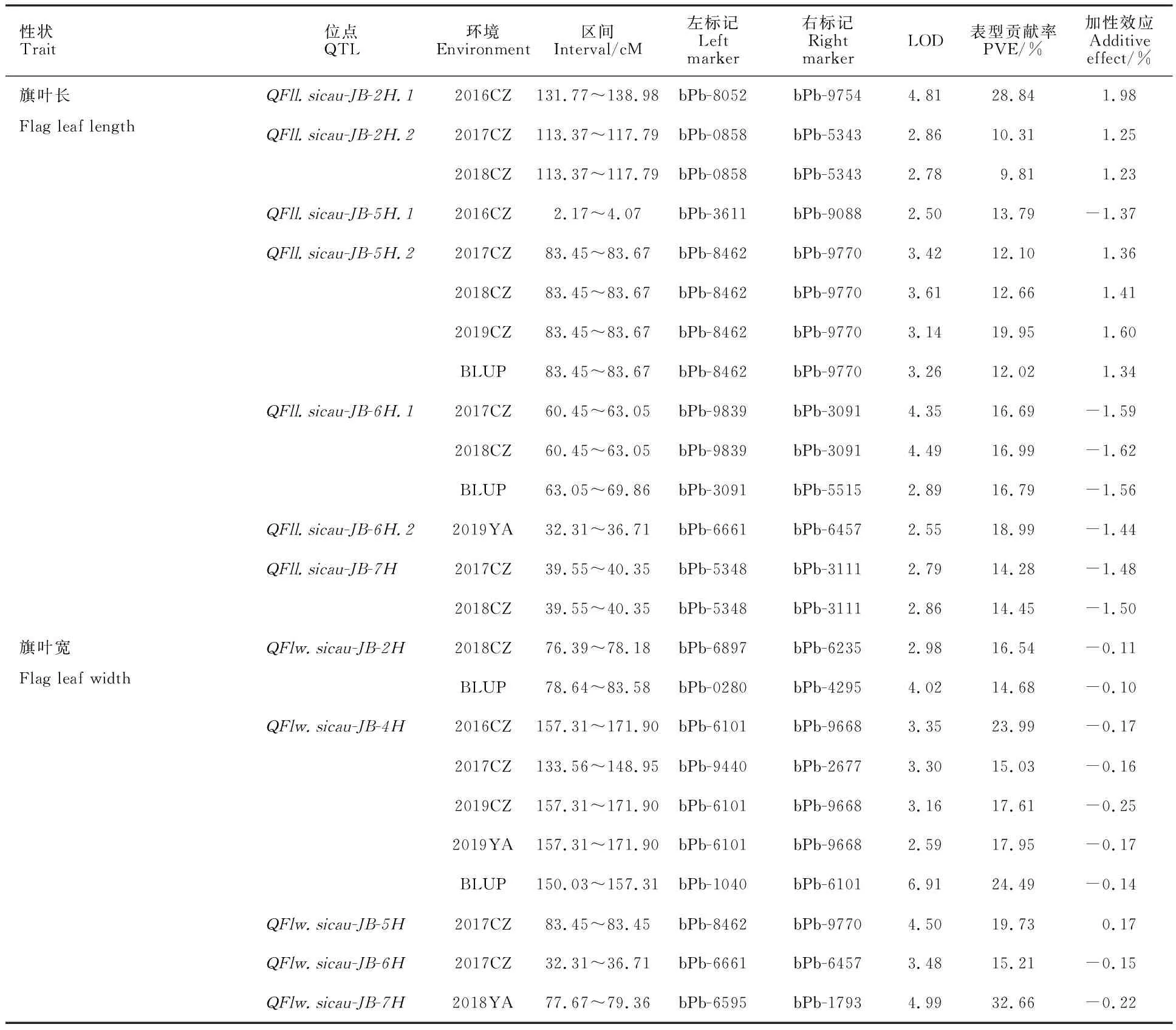
表4 基于大麦RIL群体检测到的与旗叶长和宽相关的QTLTable 4 QTLsfor flag leaf length and width detected in the J36528/BMJ89 (JM) population
检测到5个与旗叶宽相关的QTL,分布在2H、4H、5H、6H和7H染色体上。其中QFlw.sicau-JB-4H在4个环境及BLUP值中被检测到,可解释15.03%~24.49%的表型变异,LOD值为2.59~6.91,为稳定表达的主效QTL,加性效应为负值,表明其加性效应来自父本BMJ89。其余4个QTL均只在单一环境中被检测到,可解释了14.68%~32.66%的表型变异。
为了进一步揭示3个稳定表达的主效位点(与旗叶长相关的QFll.sicau-JB-5H.2和QFll.sicau-JB-6H.1以及与旗叶宽相关的QFlw.sicau-JB-4H)的遗传效应,根据以上3个位点的侧翼标记,将RIL群体中的92个株系分为2种类型,一种是携带相应QTL增效位点的株系,另一种是携带相应QTL减效位点的株系。进一步对2种类型进行检测,结果(图2)表明,对于3个QTL而言,含有各自QTL增效位点株系的旗叶长或旗叶宽在各个环境下都显著或者极显著高于含有减效位点的株系(除了QFll.sicau-JB-5H.2位点的2019YA环境)。
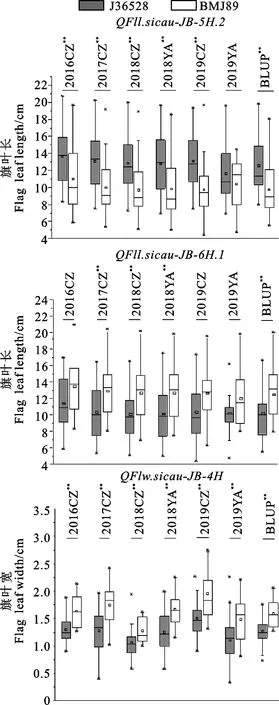
*和**分别表示各环境下亲本J36528和BMJ89之间在0.05和0.01水平上差异显著。白色框代表带有BMJ89等位基因的纯合株系,灰色框代表带有J36528等位基因的纯合株系。*代表异常值;□代表平均数。*and**indicate significant differences between J36528 and BMJ89 at the 0.05 and 0.01 levels,respectively.White box represents homozygous lines with BMJ89 allele,and gray box represents homozygous lines with J36528 allele.* represents outlier;□ represents mean value.图2 旗叶长和宽主效QTL的遗传效应Fig.2 Genetic effect of major QTLs for flag leaf length and width
2.5 与旗叶长相关的2个主效且稳定表达QTL的聚合效应
本研究检测到2个控制大麦旗叶长的主效且稳定表达的QTL,即QFll.sicau-JB-5H.2和QFll.sicau-JB-6H.1。为了进一步分析这2个QTL的聚合效应,本研究根据QTL的侧翼标记对含有这2个QTL不同效应位点的株系进行分析。结果(图3)发现,含有2个QTL增效位点的株系(20个)的旗叶长显著或极显著高于不含任何位点株系(12个)以及仅含有QFll.sicau-5H.2或QFll.sicau-6H.1增效位点的株系,增幅分别为73.28%、14.33%和20.60%。仅含有QFll.sicau-5H.2或QFll.sicau-6H.1增效位点株系的旗叶长也极显著高于不含任何位点的株系,增幅分别为51.58%和43.68%。但含有QFll.sicau-5H.2和QFll.sicau-6H.1增效位点的株系之间旗叶长无显著差异。
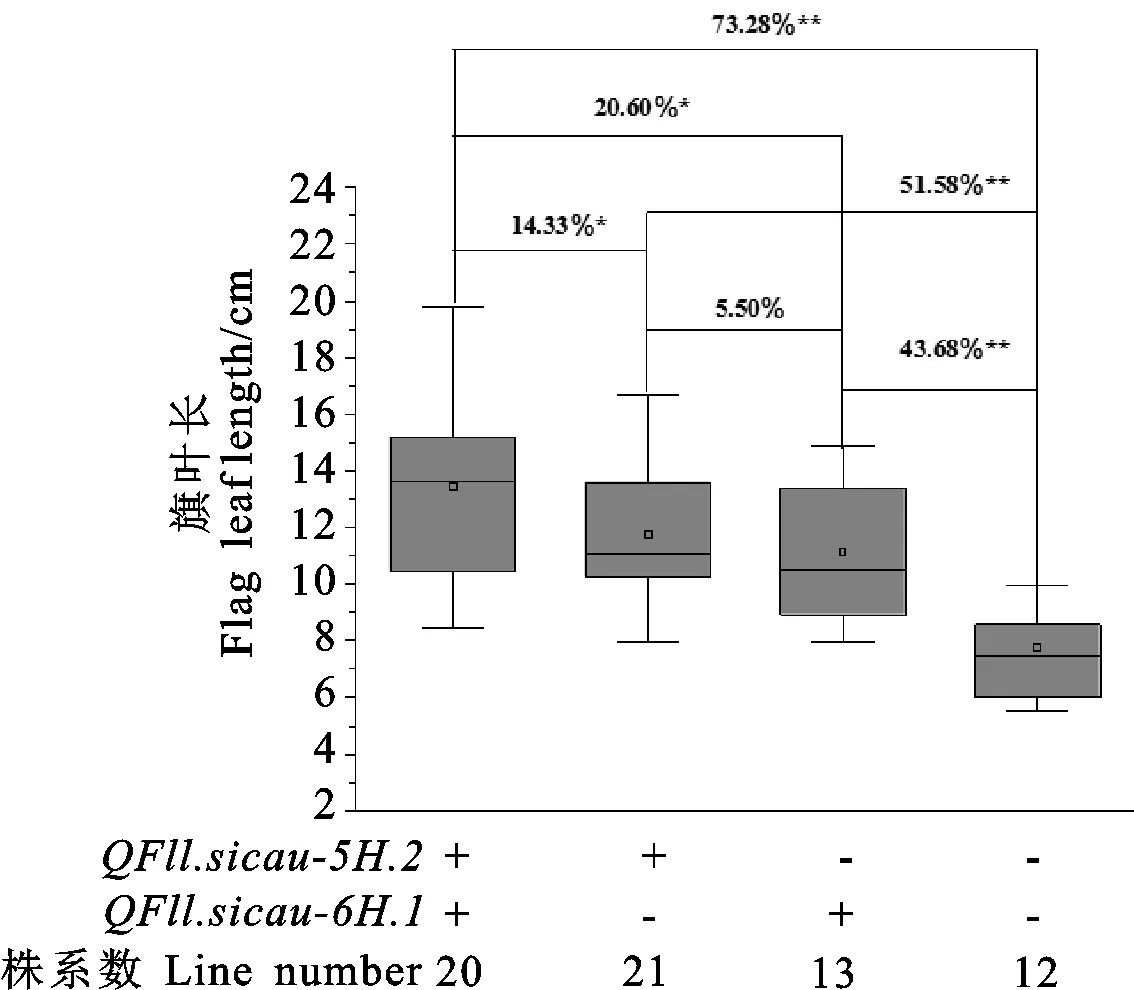
+和-分别代表携带和不携带目标QTL增效位点的株系。图中百分比数据表示两个株系之间旗叶长的变幅。*和**分别表示株系之间在0.05和0.01水平上差异显著。+ and - represent lines with and without the positive alleles of the target quantitative trait loci (QTL),respectively.The percentage data in the figure represent the variation of flag leaf length between two lines. * and ** indicate significant differences between lines at 0.05 and 0.01 levels,respectively.图3 两个旗叶长主效QTL的聚合效应Fig.3 Multiplying effect of the two major QTLs on flag leaf length of the RIL population
2.6 主效QTL在大麦参考基因组的物理位置
将主效位点QFlw.sicau-JB-4H、QFll.sicau-JB-5H.2和QFll.sicau-JB-6H.1的侧翼标记序列锚定到大麦参考基因组上,发现QFlw.sicau-JB-4H位于大麦4H染色体长臂584.05~612.36 Mbp区间,QFll.sicau-JB-5H.2位于5H染色体长臂379.10~540.62 Mbp区间,QFll.sicau-JB-6H.1位于6H染色体短臂32.39~ 33.59 Mbp区间(图4)。
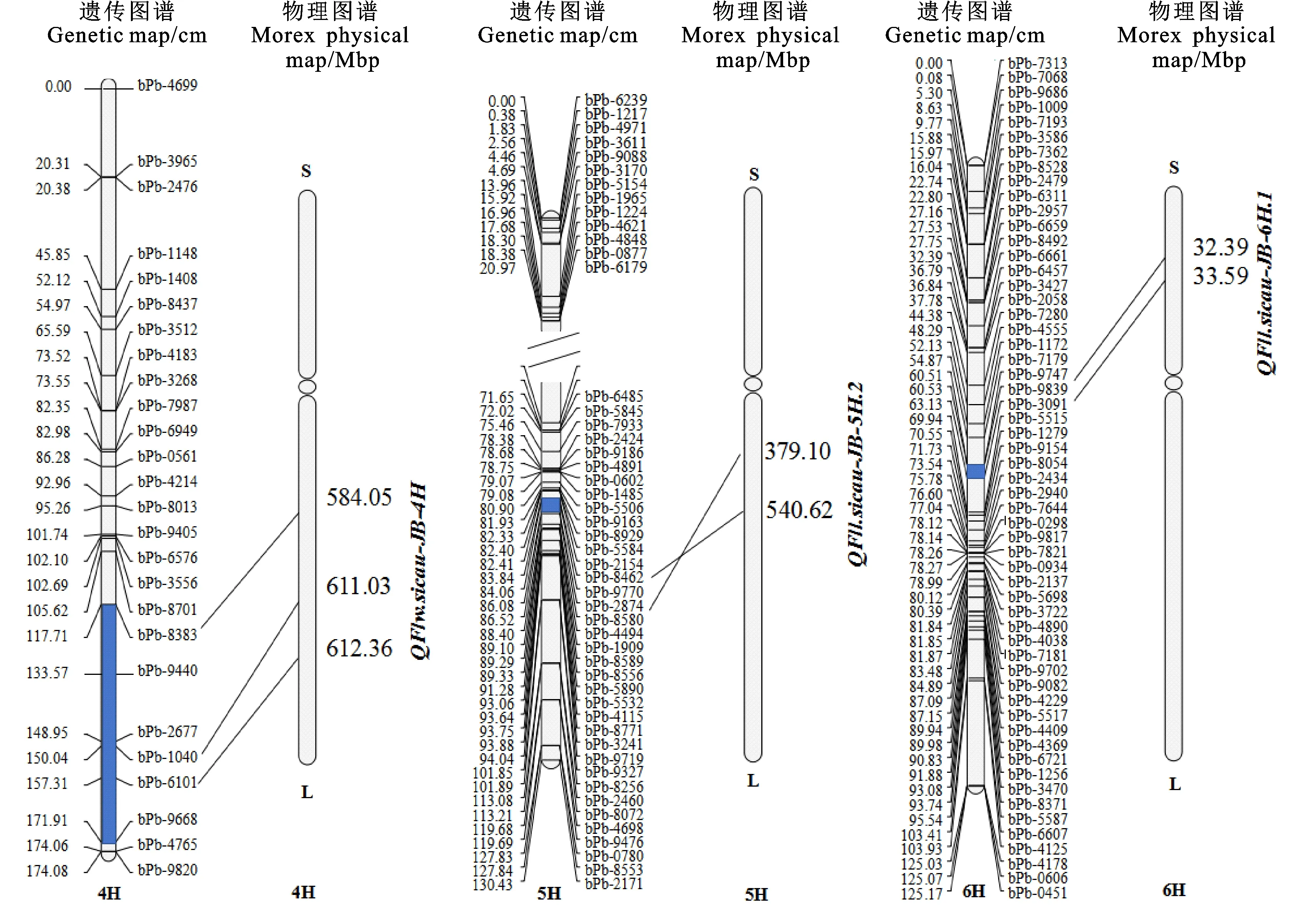
图4 大麦旗叶长和宽3个主效QTL的遗传图谱和物理图谱Fig.4 Genetic maps and physical maps of the three major QTLs for flag leaf length and width
3 讨 论
3.1 加快推进挖掘大麦旗叶形态调控位点
大麦作为第四大谷类作物,其产量增长对人们的生活有着重要意义。旗叶对于谷物产量的积极作用已有很多报道,例如Sakamoto等[9]发现,旗叶较直的水稻新品种可以增加光合作用所需的光捕捉量,进而增加籽粒灌浆所需的氮素储存量,最终提高籽粒产量;Ma等[11]发现,旗叶大小和形态是小麦“理想型”的两个重要因素,适当的旗叶长和宽可以增加小麦产量;Simon等[12]研究也发现,小麦的旗叶角度和旗叶面积是决定产量的重要性状。除了与产量相关的报道外,在小麦、水稻和玉米中,还有很多关于旗叶形态调控位点鉴定的报道[13-15,26]。而在大麦中,前人对于大麦旗叶的研究多集中在光合特性等生理生化指标方面[27-29],而鲜有关于大麦旗叶相关性状位点的报道,到目前为止,对于大麦旗叶遗传调控机制的认识还很有限。大麦作为重要粮食作物和饲料加工作物,提升其产量和品质非常必要,本研究获得的大麦旗叶相关性状定位结果,为进一步推进大麦旗叶形态的遗传研究和精细定位奠定了基础,也为产量的提升提供了更多的可能。
3.2 大麦RIL群体旗叶受环境的影响较小
前人研究发现,旗叶形态受环境影响较大[11,30],而本研究发现,大麦旗叶长和宽在不同环境下极显著相关,即受到环境的影响较小,原因还有待进一步探究。本研究也检测到3个主效且稳定表达的QTL,说明这些位点在实际育种中具有潜在利用价值。
3.3 与旗叶形态相关的稳定表达的主效QTL
本研究对RIL群体125个株系中的92个单株进行了标记扫描,构建的图谱准确可信,基于该图谱成功鉴定到3个主效且在多个环境稳定表达的旗叶长和宽QTL。迄今为止,关于大麦旗叶长和宽位点鉴定的报道极少。Liu等[6]在4H染色体长臂上鉴定获得1个控制大麦旗叶宽的位点qFLW4-18,经过比对发现,其位于4H染色体601.76 Mbp处,与本文鉴定到的旗叶宽位点QFlw.sicau-JB-4H位于同一个区间,因此,推测两者可能为等位基因。而Liu等[6]鉴定获得的大麦旗叶长QTL主要分布于2H和3H染色体上,在其他染色体未见报道。因此,推测QFll.sicau-JB-5H.2和QFll.sicau-JB-6H.1可能为控制大麦旗叶长的新位点。值得注意的是,在小麦第5部分同源群染色体长臂已检测到主效且稳定表达的旗叶长位点,如QFll.sicau-5B.3[11]、QFll.sau-SY-5B[10]和QFll.cau-5B[31]。由于小麦和大麦染色体具有一定的线性相关性,因而,QFll.sicau-JB-5H.2有可能与这些位点具有同源性。此外,小麦春化基因Vrn-B1也位于5B染色体长臂上[32-33]。这些位于小麦和大麦第5部分同源群长臂的旗叶相关QTL与春化基因的相关性有待进一步揭示。
3.4 与旗叶长相关的主效且稳定表达QTL的聚合效应
对控制同一性状的不同位点进行聚合,通常会起到一定的叠加效应[34]。本研究对控制旗叶长的主效位点QFll.sicau-JB-5H.2和QFll.sicau-JB-6H.1进行聚合分析后发现,二者叠加后旗叶长极显著增加。这为大麦育种中通过聚合不同位点来改良旗叶性状提供了理论依据。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
中国现代医生(2022年21期)2022-08-22
种子(2022年6期)2022-08-03
休闲读品·天下(2021年2期)2021-10-08
疯狂英语·新读写(2021年2期)2021-02-25
天津医科大学学报(2021年1期)2021-01-26
上海师范大学学报·自然科学版(2020年5期)2020-12-18
江苏农业科学(2020年14期)2020-08-28
三农资讯半月报(2020年2期)2020-03-09
南方农业学报(2018年9期)2018-09-10
