
八倍体小偃麦与普通小麦杂交后代中小麦染色体相互易位类型的鉴定
2022-01-06 03:03李军帅黄美瑕付体华
麦类作物学报 2021年11期
李军帅,黄美瑕,宋 敏,付体华
(四川农业大学农学院,四川成都 611130)
小麦是世界上主要粮食作物之一,同时也是我国第三大粮食作物,其生产的安全性对保障中国粮食安全和社会稳定具有重要的意义[1]。在长期的小麦育种中,由于注重骨干或核心高产品种资源之间的杂交,当今广泛种植的小麦栽培品种遗传基础日益狭窄,遗传多样性逐渐降低,因此,开发新的小麦遗传资源对解决小麦育种当前问题以及对小麦种质资源的创新至关重要[2-3]。
染色体在生物进化进程中会发生变异,其中易位是其重要变异方式之一。染色体易位可以在物种内自然产生,也可以人工诱导产生,包括利用物理化学诱导剂、远缘杂交等染色体工程技术。染色体易位不仅使遗传连锁群发生改变,而且影响基因的表达,生物性状也随之变化。染色体易位也是物种进化进程中新基因组产生的重要驱动力量,新物种的形成往往伴随着染色体易位的发生[4-5]。另外染色体易位可使物种在特定环境中具备更好的适应性,如小麦中4A、5A与7B染色体之间的环状易位,4AL-5AL易位早在二倍体水平上就已经发生,之后在四倍体小麦中进一步与7B染色体发生易位,形成4AL-5AL-7BS易位,前人根据该易位,提出了有关染色体倒位与易位的起源进化假说[6-12]。因此,发现更多染色体水平的易位,不仅可以验证相关易位理论,还会丰富小麦的遗传变异[13-14]。
Huang等[15]对我国373个栽培小麦品种进行了荧光原位杂交(fluorescence in situ hybridization,FISH)分析,发现49个品种中含有小麦相互易位染色体;Badaeva等[16]从世界不同区域的406种多倍体小麦中检测出大量的不同染色体易位类型,且欧洲小麦中广泛存在5B与7B染色体的易位。研究表明,染色体易位提高了小麦的黄锈病抗性以及对当地环境的适应性[17-18];除此之外,多个带有优良农艺性状的小麦品种(如萨达姆27、扬麦3号、丰抗13、Eshimashinrik、川麦62等)都被鉴定出小麦染色体发生了相互易位[19-23]。这些结果表明,染色体易位在现代小麦品种中分布十分普遍,在生产实践上发挥了重要作用。
自20世纪以来,育种学家们进行了大量有关小麦与其近缘物种远缘杂交的研究,极大地丰富了小麦的遗传资源库。远缘杂交产生的杂种后代中,也会出现多种小麦染色体之间的易位变异[24]。然而,大多数研究者主要关注于小麦与外源染色体之间所产生的变异及其对小麦性状的影响,而对小麦染色体之间易位的相关研究报道还比较少,因此,其遗传育种价值未得到有效利用。本课题组前期在利用八倍体小偃麦品系12-1179与普通小麦川麦107杂交转移偃麦草优异基因的过程中,获得了一些性状发生明显变异的小麦染色体之间易位的材料。本研究利用非变性FISH(ND-FISH)技术对4个小麦染色体相互易位品系(TC、19Y-145、19Y-185、19Y-200)进行分析鉴定,以期为创新小麦优异种质资源奠定基础。
1 材料与方法
1.1 试验材料
本研究所鉴定的4个小麦品系(TC、19Y-145、19Y-185、19Y-200)皆为八倍体小偃麦品系12-1179与普通小麦品种川麦107杂交后再与川麦107回交,然后至少连续自交10次以上的稳定品系,由四川农业大学农学院实验室提供。由于品系TC对条锈病表现为免疫,并且大穗多花、籽粒饱满、千粒重较高,因此重点对其农艺性状与其亲本川麦107进行对比,同时与高感条锈病小麦品系SY95-71杂交得到F1代,F1代自交得到F2代,对其条锈病抗性进行遗传分析。
1.2 试验方法
1.2.1 ND-FISH分析
染色体制片按照Han等[26]的方法进行。ND-FISH分析所采用的寡核苷酸探针包括能够区分小麦染色体的Oligo-pSc119.2-1和Oligo-pTa535-1[27]、区分小麦B和D染色体组的Oligo-B和Oligo-D[28]以及着丝粒特异探针Oligo-CCS1[27]。其中Oligo-pSc119.2-1、Oligo-D、Oligo-CCS1三种寡核苷酸探针以6-FAM-5-dUTP(绿色)进行标记,Oligo-pTa535-1、Oligo-B以Tamra-5-dUTP(红色)进行标记,探针由上海英骏生物技术有限公司合成。杂交程序参照Fu等[29]的方法。在每管探针粉末中用 1×TE缓冲液(pH=7.0)溶解为100倍的探针母液,用2×SSC和1×TE缓冲液进一步再稀释10倍成为探针工作液。每张载玻片上滴加10 μL探针工作液,置于37 ℃恒温水浴锅中杂交2 h,杂交完后用2×SSC溶液清洗,晾干后向每张载玻片上滴加10 μL DAPI复染液,黑暗环境下放置10 min后,用荧光显微镜Nikon ECLIPSE 80i进行观察并拍照。
1.2.2 农艺性状及抗病性调查
将供试材料种植于四川农业大学农场。品系TC与亲本川麦107各种植4行,2个重复,成熟后各行随机取10株对其有效分蘖、小穗数、穗长、千粒重等农艺性状进行调查。品系TC与高感条锈病品系SY95-71杂交F1和F2代在田间分别种植1行和8行,每行15株左右,以SY95-71作为对照,于分蘖期接种含有CYR31、CYR32、CYR33、CYR34、水源11等条锈菌混合菌种,待对照叶片全部感染后对F1和F2代进行观察并记载反应型(infection type,IT),按0、0;、1、2、3、4六级标准记载,其中,0、0;、1、2为抗病型,3和4为感病型[25]。
2 结果与分析
2.1 4个小麦品系的亲本核型
八倍体小偃麦品系12-1179是以中国春为母本,与中间偃麦草杂交得到F1代杂种,再与小麦栽培品种绵阳26回交得到F2代杂种,从F2代杂种连续自交过程中筛选结实性好的植株,繁殖至少8代而形成,杨 园等[30]对其染色体结构进行了研究,结果表明,其染色体构成为2n=56,含有 12条中间偃麦草染色体和44条小麦染色体,其中5D染色体为4条,没有发现小麦之间易位的染色体。本研究采用混合寡核苷酸探针Oligo-pSc119.2-1和Oligo-pTa535-1对亲本川麦107进行ND-FISH分析,对照Tang等[27]的小麦标准ND-FISH核型,也未发现其存在小麦之间易位的染色体(图1),表明所鉴定材料发生的易 位均在八倍体小偃麦与普通小麦杂交过程中 产生。
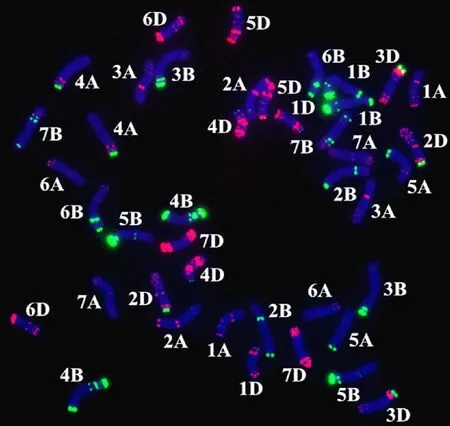
绿色和红色分别表示Oligo-pSc119.2-1和Oligo-pTa535-1探针信号。Green and red indicate the signals of Oligo-pSc119.2-1 and Oligo-pTa535-1,respectively.图1 川麦107的ND-FISH杂交结果Fig.1 ND-FISH hybridization result of Chuanmai 107
2.2 4个小麦品系的核型
2.2.1 品系TC的核型
随机选取10粒TC的种子,待发根后进行染色体计数和ND-FISH分析,结果表明,其染色体数目为2n=42。利用寡核苷酸探针Oligo-pSc119.2-1和Oligo-pTa535-1对染色体进行分析,发现TC在7B和3D染色体之间发生了相互易位(图2A),易位染色体在短臂之间和长臂之间相互结合。采用混合探针Oligo-B和Oligo-D也证实了这种结果(图2B)。进一步用着丝粒探针Oligo-CCS1明确品系TC的着丝粒位置,并与混合探针Oligo-B和Oligo-D的杂交结果进行对比,发现易位染色体断裂点在着丝粒区域(图2C和2D),说明小麦品系TC具有一对3DS·7BS和一对3DL·7BL相互易位纯合染色体。

A:以Oligo-pSc119.2-1(绿色)和Oligo-pTa535-1(红色)作探针;B和D:以混合的Oligo-B(红色)和Oligo-D(绿色)作探针,图B和图D分别表示不同的细胞;C:以Oligo-CCS1(绿色)作探针。图4和图5同。白色箭头指示3DL·7BL易位染色体,红色箭头指示 3DS·7BS易位染色体。A:Oligo-pSc119.2-1(green) and Oligo-pTa535-1(red) were used as ND-FISH probes;B and D:Mixed Oligo-B(red) and Oligo-D(green) were used as ND-FISH probes,figure B and figure D indicate the different cells;C:Oligo-CCS1(green) was used as the ND-FISH probe.The same in figure 4 and figure 5.White and red arrows indicate 3DL·7BL and 3DS·7BS translocation chromosomes, respectively.图2 品系TC的ND-FISH分析Fig.2 ND-FISH analysis of line TC
2.2.2 品系19Y-145的核型
随机选取10粒19Y-145的种子,待发根后进行染色体计数和ND-FISH分析,结果表明,19Y-145的染色体数目为2n=42条。用寡核苷酸探针Oligo-pSc119.2-1和Oligo-pTa535-1对染色体进行分析,发现19Y-145在2D与3B染色体之间发生了相互易位(图3A),易位染色体在短臂之间和长臂之间相互结合。采用混合寡核苷酸探针Oligo-B和Oligo-D也证实这种结果(图3B)。进一步用着丝粒探针Oligo-CCS1明确品系19Y-145的着丝粒位置(图3C),并与混合探针Oligo-B和Oligo-D的杂交结果进行比较,发现易位断裂点也处于着丝粒区域,说明品系19Y-145具有一对2DS·3BS和一对2DL·3BL相互易位染 色体。

A:以Oligo-pSc119.2-1(绿色)和Oligo-pTa535-1(红色)作探针;B:以混合Oligo-B(红色)和Oligo-D(绿色)作探针;C:以Oligo-CCS1(绿色)作探针。白色和红色箭头分别指示2DL·3BL和 2DS·3BS易位染色体。A:Oligo-pSc119.2-1(green) and Oligo-pTa535-1(red) were used as ND-FISH probes. B:Mixed Oligo-B(red) and Oligo-D(green) were used as ND-FISH probes. C:Oligo-CCS1(green) was used as the ND-FISH probe. White and red arrows indicate 2DL·3BL and 2DS·3BS translocation chromosomes,respectively.图3 品系19Y-145的ND-FISH分析Fig.3 ND-FISH analysis of line 19Y-145
2.2.3 品系19Y-185的核型
随机选取10粒19Y-185的种子,待发根后进行染色体计数和ND-FISH分析,结果表明,该品系的染色体数目为2n=42。使用探针Oligo-pSc119.2-1和Oligo-pTa535-1对染色体进行分析,发现19Y-185在7B与4D 染色体之间发生了相互易位,易位染色体在短臂之间和长臂之间相互结合(图4A)。利用混合寡核苷酸探针Oligo-B和Oligo-D也证明了这种结果(图4B)。进一步用着丝粒探针Oligo-CCS1明确品系19Y-185的着丝粒位置,并与混合探针Oligo-B和Oligo-D的杂交结果比较,发现19Y-185易位染色体的断裂点位于7B染色体的长臂和4D染色体的短臂上(图4C和4D),具有一对4DS-7BS·7BL和一对4DL·4DS-7BL相互易位染色体,但具体断裂点还需采用分子生物学方法进行进一步确定。
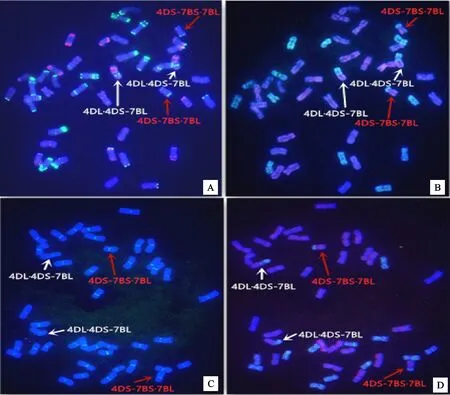
白色和红色箭头分别示4DL·4DS-7BL和4DS-7BS·7BL易位染色体。White and red arrows indicate 4DL·4DS-7BL and 4DS-7BS·7BL translocation chromosomes,respectively.图4 品系19Y-185的ND-FISH分析Fig.4 ND-FISH analysis of line 19Y-145
2.2.4 品系19Y-200的核型
对19Y-200进行染色体计数和ND-FISH分析发现,19Y-200的染色体数目为2n=42条。用寡核苷酸探针Oligo-pSc119.2-1和Oligo-pTa535-1对染色体进行分析,发现19Y-200在3B与3D染色体之间发生了相互易位(图5A)。使用混合寡核苷酸探针Oligo-B和Oligo-D也证实了这种结果(图5B)。用着丝粒探针Oligo-CCS1明确品系19Y-200的着丝粒位置,并结合混合探针Oligo-B和Oligo-D的杂交结果,发现19Y-200易位染色体的断裂点在3B和3D染色体的长臂上(图5C和5D),具有一对3BS·3BL-3DL和一对3BL-3DS·3DL相互易位染色体,但具体断裂点还需通过分子生物学方法进一步确定。

白色和红色箭头分别示3DL-3BS·3BL和3DS·3DL-3BL易位染色体。White and red arrows indicate 3DL-3BS·3BL and 3DS·3DL-3BL translocation chromosomes. respectively.图5 品系19Y-200的ND-FISH分析Fig.5 ND-FISH analysis of line 19Y-200
2.3 品系TC的农艺性状和条锈病抗性
从外观形态来看,品系TC与亲本川麦107具有明显差异,TC对条锈病表现免疫(IT=0),而川麦107表现为感病(IT=3~4),同时TC的每穗小穗数、穗长和千粒重显著或极显著高于川麦107,因此品系TC具有比川麦107更优异的农艺性状(表1和图6)。

A:植株;B:麦穗;C:籽粒。在每个小图中,左为品系TC,右为亲本川麦107。A:Plant; B:Spike; C:Grain. In each picture,TC is on the left,and Chuanmai 107 is on the right.图6 品系TC与亲本川麦107的形态特征Fig.6 Morphology features of line TC and Chuanmai 107
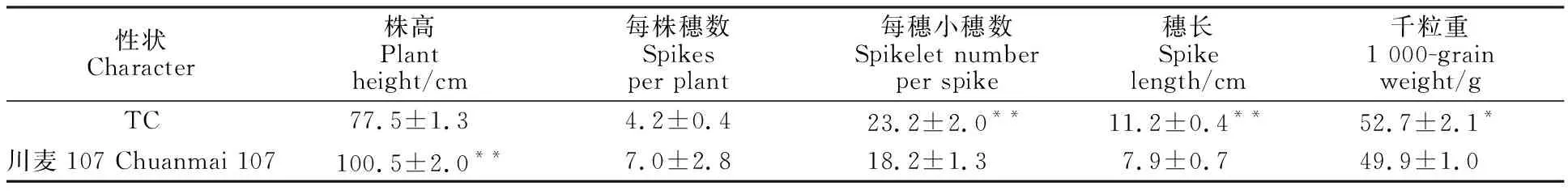
表1 品系TC与CM107的农艺性状比较Table 1 Comparison of agronomic traits between line TC and Chuanmai 107
对TC与SY95-71杂交后得到的F1代和自交获得的F2代的条锈病抗性进行调查,结果表明,在SY95-71重度感染(IT=4)的情况下,杂种F1代和TC均对条锈病表现为免疫。杂种F2代共有109株,其中抗病株数为74株,感病株数为35株,经χ2测验,符合3∶1分离比例,说明品系TC的条锈病抗性是受一对显性单基因控制。
3 讨 论
染色体易位是物种进化过程中基因组形成的重要驱动力,易位后的基因拥有更高的进化速度,其不仅改变了基因的连锁群及其位置(表现出位置效应等遗传现象),而且基因的重组频率也发生改变[31]。杨财容[32]通过研究不同区域的六倍体披碱草,发现高海拔的六倍体披碱草属物种具有更复杂的染色体组间易位,推测可能与其所处环境有关,染色体易位使其更能适应高海拔环境。易位也是植物改变染色体近着丝粒部分连锁情况的主要方式,在小麦的进化中发挥了较大作用[33]。
普通小麦作为异源六倍体作物,在长期进化和人工育种进程中,染色体之间存在大量的易位变异。其中,5B-7B、3B-3D、4A-6B、2A-3A、2B-3B、3B-7B和2D-7B是常见的染色体易位类型[34]。但是由于受技术限制,多数易位的断裂点还不确定,仅有5B-7B、5B-6B、4B-6B等少数几个易位类型由于有明显的Giemsa C带带纹特征,通过Giemsa C带技术可确定是染色体之间的臂间易位[20,35-36]。本研究4个小麦品系(TC、19Y-145、19Y-185、19Y-200)均是八倍体小偃麦和川麦107通过远缘杂交创制的染色体易位材料。采用ND-FISH和特定的重复序列为探针进行多重FISH分析,发现TC和19Y-145分别是7B-3D和3B-2D通过着丝粒断裂而形成的臂间易位纯合品系,品系TC含有一对3DS·7BS和一对3DL·7BL相互易位染色体,品系19Y-145含有一对2DS·3BS和一对2DL·3BL相互易位染色体;而品系19Y-185中易位染色体的断裂点在7B染色体的长臂和4D染色体的短臂上,形成的是一对4DS-7BS·7BL和一对4DL·4DS-7BL染色体相互易位的纯合体;品系19Y-200中易位染色体的断裂点位于3B和3D染色体的长臂上,形成的是一对3BS·3BL-3DL和一对3DS·3DL-3BL染色体相互易位的纯合体。这4种易位类型除3B-3D染色体易位类型[37]外,其余3种未见前人进行过相关报道,是新的小麦染色体核型变异类型。
Qi等[38]研究表明,普通小麦染色体的B基因组比A和D基因组更容易进行重排。本研究中所鉴定的四种染色体变异均为B和D染色体组之间的易位,而未发现A基因组的易位。但前人研究认为,A和B染色体组之间更易发生易位[34],原因可能是本研究鉴定到的4个小麦品系为八倍体小偃麦和普通小麦杂种后代的高世代材料,而在八倍体小偃麦和普通小麦杂种后代的低世代材料中,由于存在大量单价染色体的断裂与融合,既会产生小麦与中间偃麦草染色体的易位,也会出现小麦自身不同染色体的易位。Law等[22]报道,不管是自然杂交品种还是育成品种,小麦自身染色体易位的发生具有一定地域特性。如5B-7B染色体易位更常见于西方国家的小麦;埃塞俄比亚四倍体小麦中发现了新的染色体易位T2A·4B、T1B·6B和T5B·6B[16,39];两份中国云南特有的云南铁壳麦中发现了1BS·1BL-2DL和2DS·2DL-1BL染色体易位[40]。本研究中鉴定到的小麦染色体4种易位类型仅发生B和D基因组染色体之间,推测可能与四川的环境、气候相关,小麦B-D染色体易位增加了其在当地环境中的适应性。
本研究的4个染色体易位材料中,品系TC表现出良好的大穗多花、籽粒饱满、千粒重高等农艺特征,而且对条锈病表现为高抗。对其抗病性分析发现,其受一对显性基因控制。因此品系TC可能是一个优良的育种材料。这些良好的农艺性状和高抗条锈病的特性是否涉及染色体易位或连锁群改变,还需进一步研究。
猜你喜欢
山西医科大学学报(2022年8期)2022-09-27
中国计划生育和妇产科(2022年7期)2022-08-02
湖南农业科学(2022年1期)2022-06-06
福建轻纺(2022年1期)2022-02-18
广西蚕业(2021年3期)2021-10-28
今日农业(2021年7期)2021-07-28
农民致富之友(2020年12期)2020-05-11
农民致富之友(2019年20期)2019-07-27
农民致富之友(2019年16期)2019-07-01
云南农业科技(2019年2期)2019-04-04
