
植物种胚败育机理研究进展
2021-12-30 07:35:18张冬野李胜男
东北农业大学学报 2021年11期
张冬野,高 萌,李胜男
(黑龙江大学现代农业与生态环境学院,哈尔滨 150080)
胚胎发育是植物生长发育过程中极为重要的步骤,但并非所有植物有性生殖过程均正常且顺利,并随之形成具有繁殖能力的后代。植物有性生殖过程繁杂、精细,受外界环境、花粉、胚囊、传粉媒介、授粉时机及授粉后发育环境等多种内外条件影响,无论哪一个步骤出现问题,均可能影响种子正常形成。因此,在植物有性生殖过程中,未能正常发育为成熟饱满种子的现象被称为种胚败育。种胚败育包括随机败育(Random abortion)和选择性败育(Selective abortion)[1]。在生长发育过程中,一些植物果实或种子败育发生不固定,不受果实或种子所在位置、果实种子数目等因素影响,这种败育现象属于随机败育;而一些植物在生长发育过程中,受自身生物学特性或外界环境因素影响,一部分发育中的幼果或种子有选择性出现败育现象,即为选择性败育,选择性败育在被子植物中发生尤为普遍,特别是在豆科、十字花科中最为常见[2],被认为是植物的一种进化机制。
许多植物存在胚败育现象(见图1)[3],尤其是一些农作物种胚败育问题造成严重的经济损失,粮食作物如水稻、小麦、玉米、谷子等;油料作物如大豆、花生、油菜等;果树如荔枝、葡萄等。胚败育问题一方面常因胚败育导致座果率降低造成减产,且严重制约杂交育种;另一方面,败育可形成无籽或无核果实,是一个优良的果实性状,如无籽葡萄、无籽西瓜、无核枣等,食用方便,深受消费者喜爱。因此,植物种胚败育机理研究一直是的研究热点,目前已在许多作物上展开深入研究。植物种胚发育是一个极为复杂的生物学过程,败育发生可能受生理生化、胚胎学、外界环境及分子调控等多方面因素影响。本研究归纳植物种胚败育胚胎学、生理生化及分子机理研究等方面进展情况,为进一步开展植物种胚败育机理研究、植物品种种质资源创新及无籽或无核果实性状育种应用提供理论基础。
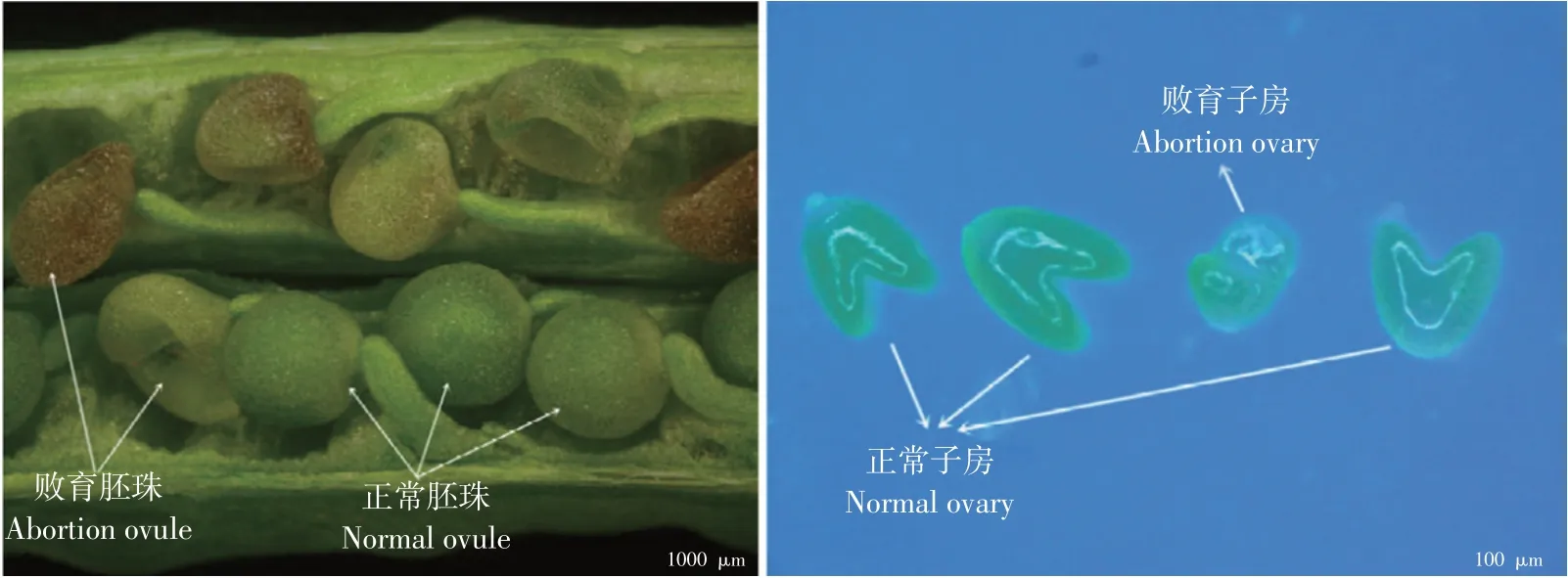
图1 甘蓝型油菜胚珠败育观察[3]Fig.1 Observation on ovule abortion of Brassica napus
1 胚胎学机制
种胚败育胚胎学机制具体包括雄性不育、雌性不育、授粉受精不良及胚中途败育等。
1.1 雄性不育
雄性不育是指雄性器官在发育过程中出现器官发育不完全、无法形成正常雄配子或产生有活力的花粉,导致败育的现象。雄性不育主要表现在雄蕊退化、花粉败育等。雄蕊退化主要表现为花药退化,花药发育畸形。水稻花药不育型材料表现为部分花药完全退化或不完全退化,完全退化的花药仅仅持有花丝结构,不完全退化花药表现扭曲且随后出现退化的现象,这两种情况均无法形成正常花粉[4]。甜橙类和柑橘类品种无核现象均与花粉退化密切相关[5]。而花粉败育主要表现为花粉母细胞减数分裂行为不正常、染色体变异或绒毡层结构异常。研究表明,单子叶植物花粉败育现象大多出现于接近双核期阶段,而双子叶植物花粉败育现象多出现在四分小孢子早期发育阶段[6]。毛白杨(Populus tomentosaCarr.)属于双子叶植物纲,其花粉败育即在小孢子发生过程中,具体表现为染色体结构缺失,同源染色体无法均匀分置于子代细胞中[7]。杏(PrunusarmeniacaL.)雄性不育发生于花粉发育末期,花粉分裂异常,绒毡层退化不适时,无法正常溶解前期花粉母细胞胼胝质壁,这些问题使酶或孢粉素无法正常达到小孢子,阻碍蛋白质合成,造成花粉败育[8]。
1.2 雌性不育
雌性不育是指由于雌性器官发育异常而导致的败育现象。雌性不育主要涉及花器官外部形态异常、胚囊发育受阻、珠被发育异常及雌蕊败育。花器官外部形态异常具体表现为子房干瘪,花柱外部形态异常。胚囊败育是导致植物雌性不育关键因素。研究李(PrunussalicinaLindl.)胚珠败育现象发现,雌配子体发育过程中胚囊发育受阻,珠心、珠被过早解体退化,这种发育异常的胚囊在发育胚中高达67%[9]。在大蒜(Allium sativumL.)雌配子体形成阶段,细胞发育不正常,珠被出现混乱,缺少珠孔,无法形成正常雌配子体和胚囊结构[10]。无核白葡萄种子败育关键在于内珠被、珠孔和胚囊组织形态异常[11]。而雌蕊败育则体现在雌蕊分化进程停滞或迟缓;花芽发育停滞,最终解体。
1.3 授粉受精不良
授粉受精在植物发育过程中要求极为严苛且复杂。授粉能否成功与授粉环境条件、花粉量多少、花粉与柱头亲和性及花期均有重要联系。授粉受精不良具体表现为:①花粉在柱头上无法正常萌发。例如越南无核橘‘Quyt Ngot’,其在自花授粉时自交不亲和,花粉粒在柱头上不发芽,花柱和子房中均未能观察到花粉管[12];②花粉在柱头上虽萌发,但花粉管在柱头上成团状或缠绕生长,无法深入花柱。导致沙糖桔无籽的关键原因为自花授粉时花粉管在柱头缠绕生长,无法进入胚珠[13];③花粉管在花柱中停止生长,无法到达胚囊,例如‘无核世纪梨’自花授粉时花粉管不进入胚囊[14];④花粉管到达胚囊后,精子未能顺利进入卵膜或进入卵膜后雌雄核不融合,或精子与极核无法正常融合,双受精失败,从而导致败育,形成无核果,例如用‘雪青’梨异花授粉产生的‘无核世纪梨’[14]。
1.4 胚中途败育
胚中途败育发生于植物授粉受精后,在合子发育至成熟胚形成阶段,由于一些原因造成胚发育停止在其中某个阶段。不同作物或同一作物不同品种间出现胚胎败育的时期也不同。无核白葡萄形成是由于胚乳核不分裂导致胚乳细胞解体、胚乳败育,幼胚中途发育停滞,无法形成具有繁殖能力的种子[11];‘南丰蜜橘’无籽主要原因也是胚胎早期败育,大概授粉4周后,大部分合子胚发育停滞、开始败育[15]。
2 环境因素
种胚败育原因除上述生理生化机制、胚胎学机制外,植物生长外部环境也严重影响种胚发育,例如温度、水分、光照及病虫害等。
2.1 病虫害
病虫害发生造成植物叶片光合作用异常,影响授粉阶段花粉供应,尤其是草食动物采食是植物生长发育过程中一大难关。害虫密度影响橄榄果实重量;甲虫采食也造成豆科植物结实率降低;被豆象(Bruchidae)长期侵害的豆科植物种子比较小[16]。
2.2 温度
温度过低或过高均不利于胚珠正常发育,增加种子败育率。荔枝品种大小孢子正常发育,但如果在授粉受精时遇到低温阴雨天气,无法顺利完成受精,便形成无核果;若天气晴好,受精正常,则发育为有核果[17]。开花前后温度骤降也是引起甘蓝型油菜胚珠败育关键因素[3]。温度过高或过低也造成许多豆科植物花和胚珠等发育不良,出现败育现象[16]。玉米从果穗形成至完全成熟过程中均对温度有要求,温度过低致使穗分化停滞,灌浆速度降低等,温度过高则影响正常开花及授粉过程[18]。低温亦导致李花粉萌发率显著降低,特别是花粉萌发速度及花粉管伸长速率,造成授粉受精质量下降,发生败育。此外,杧果、柑橘及樱桃等在花期如遇高温干旱或连续阴雨天气也可能影响授粉受精,形成种胚败育果实[19]。
2.3 光照
光照不足亦影响授粉受精,小花难以成熟,或授粉后因光合作用不充分、营养严重不足从而导致种胚败育。在玉米籽粒形成阶段如遇遮光,造成已受精籽粒全部败育[20]。同时,高粱不育系小花败育也受光照影响,对光温反应最为敏感[21]。此外,日照长短也影响植物种胚发育。长日照减慢谷子穗发育,短日照加快穗发育;短波促进穗分化,长波则抑制雌穗小花发育[22]。
2.4 水分
水分胁迫不仅抑制植株生长,也对种胚发育造成影响。干旱胁迫时,极易造成花期不遇;水分过于饱和造成花粉失活。研究表明,玉米在干旱情况下,植株光合作用受抑制,果穗籽粒营养不足,导致处在劣势位置的顶部籽粒大面积败育[23];若在小花分化期遭遇干旱,籽粒几乎全部败育。充足水分也减少野豌豆胚珠败育,促进种子成熟,极大提高野豌豆结实率[24]。
3 生理生化机制
3.1 内源激素含量
植物种胚发育受多种生理、生化指标综合影响。内源激素含量、保护酶活性、多胺和酚类物质及矿质营养元素等均影响胚正常发育。内源激素涉及植物生长、发育等各个过程,如组织和器官分化、开花和结实、成熟和衰老、休眠和萌发等方面。生长素(Auxin,IAA)刺激胚中途停止发育,发生败育问题。芒果种仁中IAA含量过低导致果核发育小且缓慢[19];辣椒雄性不育系叶片与花中内源IAA含量也均显著低于相应保持系[25];也有研究表明,种子若含有较高含量IAA促进种胚发育,且在生长后期,无核果实中果肉IAA含量大于正常果实[26]。研究表明,赤霉素(Gibberellin,GA3)活性影响种胚发育。在葡萄开花期施用GA3,可对授粉受精、胚囊发育造成一定影响,从而发生胚败育形成无籽葡萄[27];此外,外源GA3促使葡萄自由态多胺含量增加而发生败育[28]。脱落酸(Abscisic acid,ABA)在种子休眠、胚胎发育及果实发育等方面均发挥重要作用。伴随果实成熟,不同作物、不同品种及不同部位ABA含量变化也不同。在葡萄胚败育研究中发现种子发育调控果实中ABA含量,有籽葡萄、空壳品种及无籽葡萄果肉中ABA含量呈明显下降趋势[29]。在植物种胚发育过程中,也有多种激素协同调节发挥作用。前人研究表明,枣胚败育受枣胚和果肉之间所含激素浓度差异影响,当种子玉米素(Zeatin,ZT)、GA3和IAA含量低于果肉时,幼胚发生败育[30]。荔枝胚胎发育研究表明,败育胚胎中IAA、GA3及细胞分裂素(Cytokinin,CTK)含量较高,而正常胚胎中IAA、GA3、CTK含量低,且高水平ABA含量不利于幼胚正常发育[31]。此外,与对照相比,用生长素抑制剂青鲜素(Maleic hydrazide,MH)处理龙眼后,ABA/GA3及ABA/IAA值明显增高,极有可能是造成龙眼焦核关键因素之一[32]。于无核葡萄花前施用IAA、CTK类生长调节剂,可能改变胚败育过程中IAA与CTK之间平衡,从而促进胚发育产生残核[33]。多种激素协同作用调控植物种胚败育已得到证明。
3.2 保护酶活性
保护酶参与种子发育过程中的生理生化反应,保护酶活性变化可直接影响植物细胞内活性氧代谢和细胞程序性死亡程度。对葡萄种子相关研究中发现,与健康发育葡萄种子相比,败育葡萄中过氧化物酶(Peroxidase,POD)、超氧化物歧化酶(Superoxide dismutase,SOD)及过氧化氢酶(Catalase,CAT)活性均显著降低[34]。还有研究表明多酚氧化酶(Polyphenol oxidase,PPO)活性过低时,阻碍荔枝幼胚中多酚类物质氧化分解,IAA含量无法维持种胚正常发育,发生败育[35]。对无核葡萄种胚败育影响因子作主成分分析和灰色关联度分析得出,胚珠SOD活性与种胚败育关联度最高,丙二醛(MDA,Malondialdehyde)含量也是胚败育主要影响因子[36]。辣椒雄性不育材料POD、CAT、PPO活性与相应保持系相比差异显著[37]。对高羊茅育性相关研究表明,不育株花药中SOD、CAT活性显著低于同时期可育株,但POD活性及MDA含量高于可育株,抗氧化酶代谢紊乱及MDA积累可能是引起高羊茅败育主要生理因素之一[38]。
3.3 多胺与酚类物质
多胺与酚类物质也参与调节植物胚胎发育。与种胚败育相关多胺类物质具体指精胺(Spm,Spermin)、腐胺(Put,Putrescine)以及亚精胺(Spd,Spermidine)等。研究表明,与正常发育胚珠相比,败育胚珠中Spm、Put及Spd含量在发育阶段均显著降低[39]。无核葡萄种胚败育也受多胺类物质含量极速降低的重要影响[34],同时,在荔枝和龙眼种胚败育相关研究中也证实正常发育有核品种内源多胺含量和多胺含量比例显著高于种胚败育品种[35]。而酚类物质负向调控植物种胚发育。酚类物质含量在大多果树种胚败育品种与种胚正常发育品种中差异显著,较高的酚类物质含量造成植物体内促进生长类物质和抑制生长类物质的平衡遭到破坏,种胚败育;也有研究认为酚类物质可能参与调控DNA的合成作用,发生种胚败育现象[40]。
3.4 矿质营养元素
矿质营养元素也与植物种胚发育联系紧密。氮素营养失调是驼绒藜属植物种子败育直接原因[41];玉米生长发育过程中如果缺乏氮、磷、钾、锌、硼、锰等则会秃顶,子房发育情况也直接影响成粒情况。土壤中速效硼缺失,导致板栗胚珠败育[42];在板栗胚珠形成和胚囊发育期供给外源营养元素氮、磷、钾、硼、锌及镁等可有效缓解胚珠败育现象发生[43]。无核葡萄相关研究也表明,锌、铁也可能是发生胚珠败育现象影响因素[27]。
4 分子机制
早期植物种胚败育研究主要停留在生理、胚胎学上。随植物全基因组序列完善、基因定位克隆方法及蛋白组学发展,对植物种胚败育分子机制方面研究也逐渐增多。
在拟南芥(Arabidopsis thaliana)中,决定种子最终大小主要因素是快速增殖的胚乳和种皮细胞[44]。Orozco-Arroyo等研究表明,拟南芥有3条控制胚乳细胞化的途径:PcG(Polycomb group protein,多梳蛋白)途径、MADS(the MADS-box transcription factor)/AP2(APETALA2)途径及IKU途径(见图2)。这3个调控途径既相互独立,又在某些位点上协同作用。因此,胚乳发育是十分精细、复杂的过程。胚乳发育直接影响种子大小,甚至决定种子是否可育。
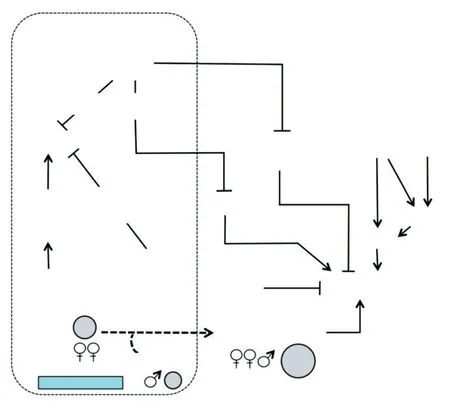
图2 拟南芥中控制胚乳细胞化的途径[44]Fig.2 Ways to control endosperm cellularization in Arabidopsis
4.1 种胚败育相关基因定位
基因调控机制也是影响种胚败育关键性因素。通过基因组测序、分子标记等手段对种胚败育基因定位十分关键。对谷子谷穗顶端败育的突变体sipaa1研究表明,谷穗顶端败育是单基因控制的性状变异;利用BSA(Bulked segregation analysis)混池测序将突变基因定位于1号染色体110 kb区间内,共有12个候选基因,其中6个在谷子穗部高表达,为下一步禾本科作物穗顶端败育相关通路研究提供基础[22]。基于多种分子标记和BAC(Bacterial artificial chromosome)克隆,成功从甘蓝型油菜C9染色体上分离得到一个正调控每角粒数的基因BnaC9.SMG7b,其在调节功能性雌配子体形成中发挥作用,决定可育胚珠形成,对油菜单株产量发挥重要作用[45-46]。也有研究表明,胚珠败育是甘蓝型油菜每角粒数变异最重要的决定因素,利用QTL(Quantitative trait locus)定位将一个控制胚珠败育的主效QTL精细定位到A8染色体80.1 kb区间内,并筛选得到两个关键候选基因,为下一步胚珠败育基因克隆和提高油菜产量奠定坚实基础[47]。利用简化基因组测序技术对番茄雄性不育基因作初步定位,得到149个雄性不育候选基因,为未来分子机制研究提供重要参考[48]。利用全基因组重测序及单核苷酸多态性(SNP,Single nucleotide polymorphism)研究榛子空壳果实突变体和野生种,筛选得到6个候选基因,其单核苷酸变异可能致使榛子胚败育,为进一步种胚败育分子机理研究及无核种质创新提供新的理论依据[49]。
4.2 组学在种胚败育中的应用
除对种胚败育相关基因定位外,从差异表达基因中挖掘、分离与种胚发育密切相关基因和蛋白也是研究种胚败育的有效方法。随测序技术发展,转录组学和蛋白组学在植物种胚败育研究上的应用十分广泛。基于转录组学和蛋白组学研究菊花胚发育,筛选得到140个差异表达基因和41个差异蛋白,为菊花胚发育下一步深入研究提供资源[50]。利用RNA-Seq比较分析‘冷白玉’枣败育高峰期正常胚与败育胚,败育胚中AUX/IAA(生长素诱导蛋白基因)、SAUR1(生长素早期响应基因)、PYR/PYL(脱落酸受体蛋白基因)、ERF1/2(乙烯应答转录因子)、GH3(吲哚乙酸酰胺合成酶)等基因显著下调表达,再次证实激素与枣胚发育密切相关,从分子生物学角度揭示枣胚败育与内源激素变化的相互关系[51]。与之相似,对谷子穗顶端败育突变体sipaa1和野生型Yugu1的转录组测序结果表明,差异基因显著富集通路主要是激素信号转导、外界胁迫响应等,再一次证明植物激素在种胚败育过程中发挥重要作用[22]。对葡萄种胚败育的研究中,通过RNA-Seq筛选得到无籽相关候选基因,且研究表明HD-Zip转录因子与葡萄胚败育密切相关,例如VvHDZ28与VvHDZ27[52]。还有相关研究对假单性结实无核葡萄‘无核白’和有核葡萄‘黑比诺’胚珠发育不同时间点作转录组测序分析,在无核白葡萄胚珠SA合成通路中两个关键基因VvEDS1和VvSARD1表达水平明显较高,而进一步研究表明,VvHDZ28正调控VvEDS1表达,而VvHDZ28过表达导致胚珠败育及内源SA含量增加,这也为葡萄胚珠败育机理研究及假单性种子败育型葡萄遗传改良提供重要依据[53]。基于对珙桐转录组测序构建的unigene库,对调控种子败育关键基因CesA作克隆和表达模式分析,为进一步研究珙桐种子败育分子机制奠定理论基础[54]。分析板栗早期子房内可育胚珠和败育胚珠蛋白组学表明,败育胚珠在早期蛋白质合成功能受损,且参与活性氧代谢途径的过氧化物酶表达量高;同时,作为糖酵解和糖异生的关键酶GAPDH(Glyceraldehyde-3-phosphate dehydrogenase),在败育胚珠中下调表达,造成细胞能量代谢系统紊乱,并进一步引起细胞衰老及死亡,这也为板栗胚珠败育机制进一步研究及为提高板栗产量奠定重要理论基础[55]。利用比较蛋白质组学对胚败育诱导的甜樱桃小果脱落机制开展研究得出,植物激素生物合成、细胞壁代谢等通路显著富集,胚败育可能导致植物激素合成紊乱,扰乱信号转导途径,控制细胞壁降解相关基因,导致小果脱落[56]。利用分子标记检测得出,MADS-box家族基因VvAGL11是控制葡萄种子败育关键基因[57]。基于对无核葡萄转录组学和基因组重测序的联合分析鉴定并分离两个差异表达基因VvMADS28以及VvMADS39,并通过功能验证得出VvMADS28参与种子发育,VvMADS39与葡萄无核相关[58]。在玉米、番茄、棉花及荔枝等作物上研究发现,对糖代谢关键酶(酸性转化酶、蔗糖合成酶等)作RNAi沉默或超表达导致种子发生败育或种子重量提高[59-61]。
5 展望
胚胎发育关系到农作物产量和品质等重要农艺性状,因此种胚败育机理研究,特别是遗传和分子机制探究尤为关键。目前关于种胚败育机理仍未清晰。因此,随着分子生物学发展,全基因组序列不断完善、测序技术升级,未来对种胚败育的研究重点应对更多种胚败育相关基因进行精细定位、克隆等,明确调控通路及之间互作过程,并结合生理生化、胚胎学等全面解析植物种胚败育机理,为农作物品种种质资源创新及无籽或无核果实性状育种应用提供理论依据。
猜你喜欢
生物技术通报(2023年2期)2023-03-07 12:54:40
哈哈画报(2022年8期)2022-11-23 06:21:20
蔬菜(2022年7期)2022-07-29 15:47:37
阅读(低年级)(2020年11期)2020-12-28 02:26:35
女报(2020年10期)2020-11-23 01:42:42
山东农业大学学报(自然科学版)(2020年2期)2020-05-21 00:18:06
中国蜂业(2018年6期)2018-01-21 15:44:41
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30 08:05:59
小青蛙报(2016年2期)2016-10-15 05:05:56
文理导航·科普童话(2016年6期)2016-09-12 23:24:08
