
浙江省毛竹秆形结构特征
2021-12-27 04:22:58汤孟平
浙江农林大学学报 2021年6期
杨 帆,汤孟平
(1. 浙江农林大学 省部共建亚热带森林培育国家重点实验室,浙江 杭州 311300;2. 浙江农林大学 环境与资源学院,浙江 杭州 311300)
毛竹Phyllostachys edulis为禾本科Gramineae刚竹属Phyllostachys单轴散生型竹类植物,具有生长更新快,自然成熟期短等优点,是中国分布最广、面积最大的经济竹种[1-2]。毛竹独特的生长习性决定了秆形结构因子及其在竹秆上的分布具有一定的规律性。毛竹的主体是竹秆,为提高毛竹林的经营利用水平,长期以来学者们十分重视毛竹秆形结构研究[3-5]。毛竹的秆形结构通常用竹节数、节长、节间周长(或直径)等因子描述。毛竹的竹节数在笋芽分化时期就已经确定,出土后不再增加新节[3]。毛竹的高生长主要依靠竹节之间的拉伸[4],节间伸长是从基部开始,靠居间分生组织细胞分裂和细胞伸长而逐节伸长的[6-8]。周芳纯[9]较早研究了节间长度、围度等在竹秆上的分布规律,依据毛竹竹秆上自近中央节位向基部和梢部两端推移,毛竹节间长度逐渐变小的特征,用正态分布近似描述节间长度的分布规律,表明节间长度通过正态分布难以反映节长的分布规律。谢芳[10]和甘代奎[11]则对标准竹竹节进行编号,赋值基部第1节竹节竹号为1,并自下而上依次编号,分析秆形结构因子在竹秆上分布规律,表明从基部向上,节长依节号的变化呈抛物线型,竹节周长与节号呈线性负相关。但是,不同毛竹的竹节数往往不同,以节号作为自变量,得出秆形结构因子与节号的关系方程不具有普遍意义。可见,现有描述毛竹秆形结构因子及其在竹秆上的分布规律仍是未解的科学问题。本研究以浙江省受人为干扰较少的毛竹林为研究对象,以竹节所处高度与竹高的比值(即相对高,取值范围0~1)代替实际高,分析秆形结构因子依赖于相对高的分布规律,旨在为毛竹林经营利用提供理论依据。
1 研究区与方法
1.1 研究区概况
浙江省位于 27°01′~31°10′N,118°01′~123°08′E,年均气温为 16~19 ℃,四季分明,光照充足,雨水充沛,是典型的亚热带湿润季风气候。浙江省是中国毛竹的主产区之一,有“七山一水二分田”之说,山地和丘陵占74.63%,因而比较适合毛竹的生长,毛竹广泛分布于海拔400~800 m的丘陵、低山山麓地带。根据《2018年浙江省森林资源及其生态功能价值公告》,浙江省森林面积为607.82万 hm2,森林覆盖率为59.71%,竹林面积为92.70万 hm2,占森林面积的15.25%,其中毛竹林面积为81.67万 hm2,占竹林面积的88.10%。
1.2 样地设置及样竹调查
2014-2018年,根据浙江省森林资源一类调查系统抽样样地中的毛竹林样地分布较多的区域,共选择10个县市(区)为调查区域,包括庆元县、泰顺县、常山县、武义县、诸暨市、宁海县、黄岩区、安吉县、临安区和余姚市(表1)。在调查区域内,选择近5 a没有采伐、垦复、施肥和钩梢等人为经营管理措施干扰的近自然毛竹林,每个区域随机设置10 m×10 m的样地至少4个,共52个样地。根据样地每木调查结果,按照胸径分布范围,确定大、中、小径级,并在大、中、小径级的样竹中,各随机选取1株梢头完整且无病虫害的样竹,共159株,总竹节数为9 431节。用锯子在竹秆基部齐地伐倒,用皮尺测量竹高。自竹秆基部向上,将竹节从小到大依次编号,称之为节号。用围尺测量每个竹节的竹节长和竹节中央直径。用游标卡尺测量竹秆基部,胸高处,及1/2高处东、南、西、北4个方位的壁厚,取4个方位壁厚的平均值作为对应位置的壁厚。

表1 研究区概况和样地样竹调查Table 1 Study area overview and sample setting
1.3 统计分析
采用SPSS 20.0对毛竹秆形结构因子在区域间的差异性进行了方差分析,对秆形结构因子之间的关系进行了相关性分析,对秆形结构因子间的主导因子进行了因子分析。
2 结果与分析
2.1 秆形结构因子描述性统计及区域差异性分析
由表2可知:10个地区毛竹的竹节数最小为39节,最大为76节,平均为59节。竹节数和1/2高节号的变异系数最小,均为0.12。胸高节号的变异系数最大,为0.20。最长竹节长与1/2高节长相近。10个研究区间,毛竹的竹节数、1/2高节长、最长竹节长、基部壁厚、1/2高壁厚、竹高和胸径间差异显著(P<0.05),其他秆形结构在10个地区间差异不显著。
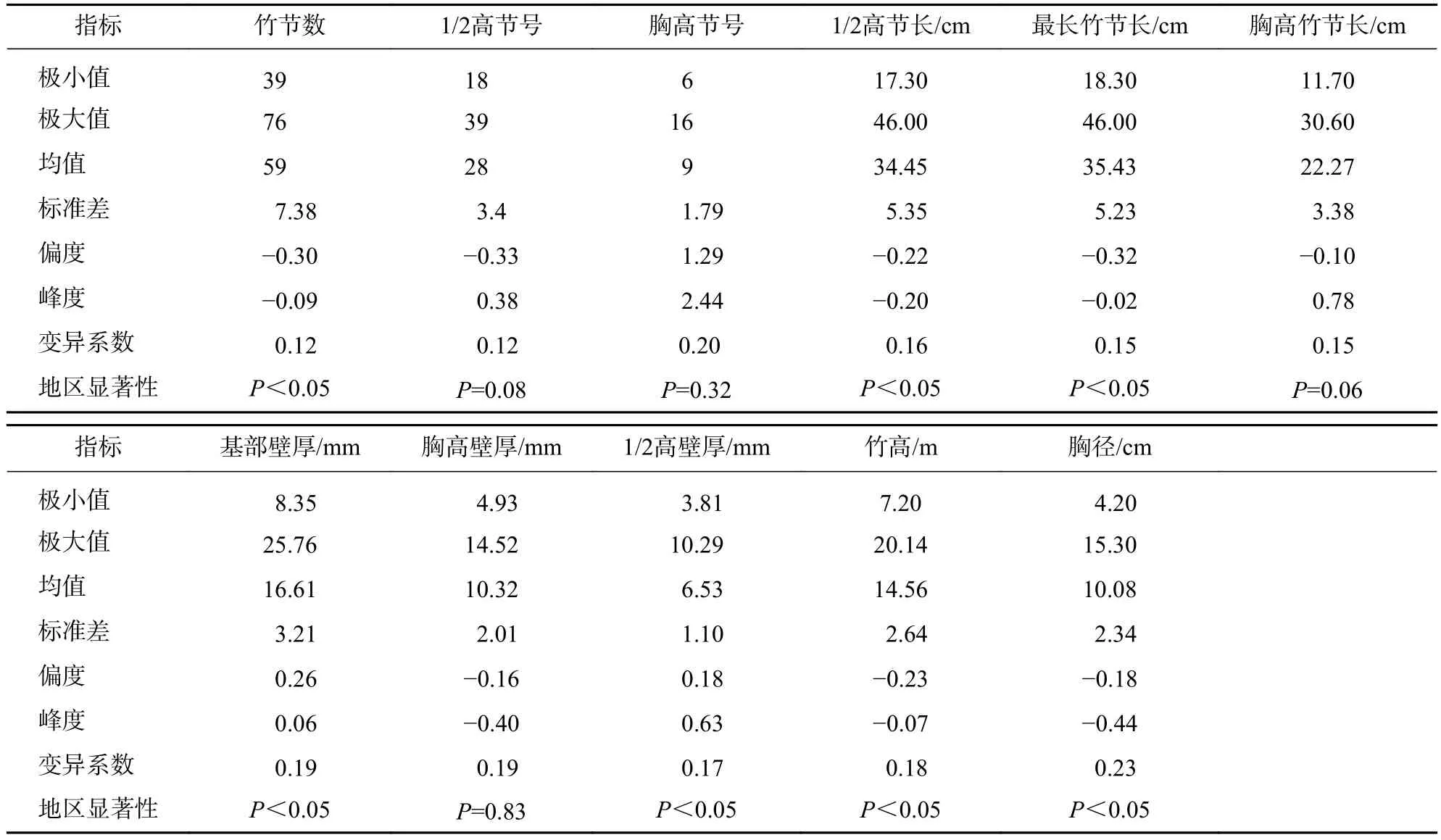
表2 毛竹秆形结构因子描述统计特征Table 2 Description statistical characteristics of factor of culm form
2.2 竹节数的分布特征
以5节竹节为竹节数级距,分析所采集样竹的竹节数频数分布特征。从图1可知:毛竹的竹节数主要集中在53~67节,且毛竹竹节数的频数分布符合正态分布(P<0.01)。从图2可见:随着径阶的增大,平均竹节数有增大的趋势,即粗壮竹子的竹节数较细小竹子多。
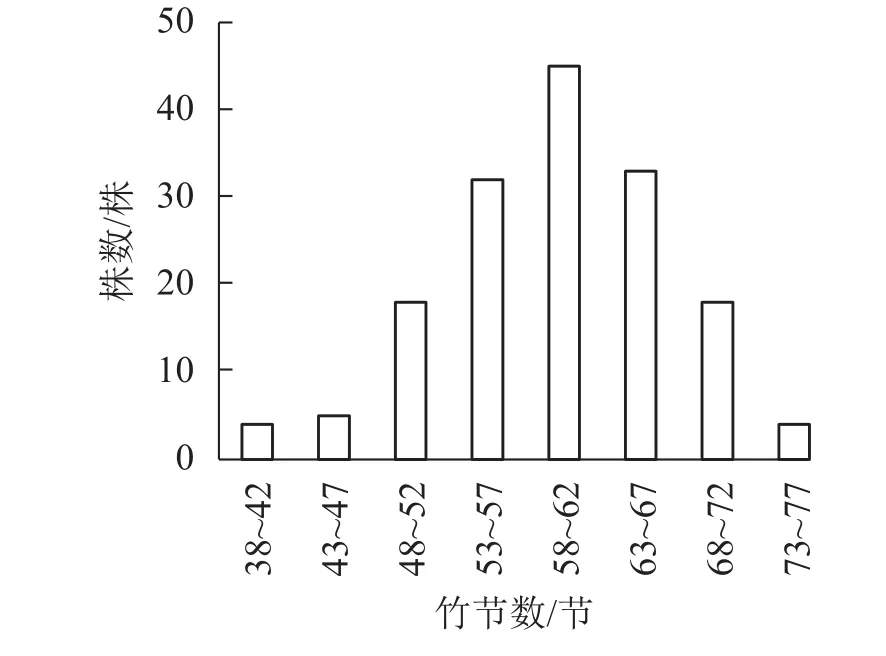
图1 竹节数的分布频率Figure 1 Frequency of number of bamboo node

图2 不同径阶竹节数Figure 2 Number of bamboo node in different diameter class
2.3 竹节长的分布特征
不同毛竹的竹节数、竹节长和竹高往往不同。为消除不同竹高的影响,以竹节所处高度与竹高的比值(即相对高,取值为0~1)代替实际高,分析竹节长在竹秆上的分布规律。从图3可见:随着竹节相对高的增加,竹节长自基部至梢头呈先增大后减小的趋势,并且竹节长在竹秆上的分布具有对称性。因此,可用抛物线描述竹节长在竹秆上的分布规律。根据函数拟合结果,在相对高为0.48时,即约1/2竹高处时,最长竹节为35.46 cm(图3)。

图3 毛竹竹节长与相对高的关系Figure 3 Relationship between relative height and length of node
为比较不同径阶毛竹竹节长在竹秆上分布的差异性,绘制了不同径阶的关系图(图4),并拟合抛物线(表3)。经检验,拟合方程均达显著水平(P<0.01)。根据方程拟合结果和不同径阶最长竹节长对应相对高的实测值,可以解析不同径阶最长竹节长对应的相对高,以及某一相对高对应的理论节长的分布规律。从表3可知:不同径阶毛竹最长竹节对应的相对高差异不大,取值均为0.47~0.52,但最长竹节长随着径阶的增大有逐渐变大的趋势。

图4 不同径阶毛竹竹节长在竹秆上的分布Figure 4 Distribution of length of bamboo node on culm in different diameter class

表3 不同径阶毛竹竹节长分布特征拟合结果Table 3 Fitting results of distribution of length of node on culm in different diameter class
2.4 竹节中央直径的分布特征
竹节中央直径可反映竹节的大小。从图5可见:随着竹节相对高的增加,竹节中央直径逐渐下降,呈明显的线性负相关关系。不同径阶毛竹竹节中央直径与相对高的关系拟合直线见图6,拟合参数见表4。经检验,拟合方程均达显著水平(P<0.01)。利用拟合方程,可推算不同径阶毛竹在某一相对高对应的理论竹节中央直径。从图6可以看出:随着径阶的增大,拟合函数斜率逐渐变小。但不论毛竹径阶怎么变化,竹秆相对高每增加10%,竹节中央直径约下降10%。
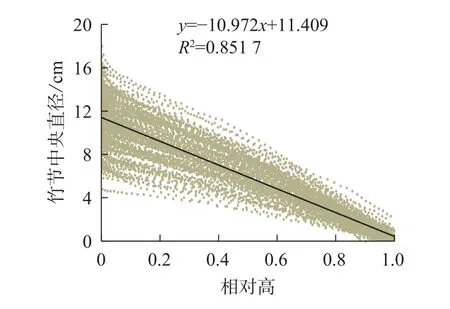
图5 毛竹竹节中央直径与相对高的线性关系Figure 5 Relationship between relative height and central diameter of bamboo node

图6 不同径阶毛竹竹节中央直径在竹秆上的分布Figure 6 Distribution of central diameter of node on culm in different diameter class
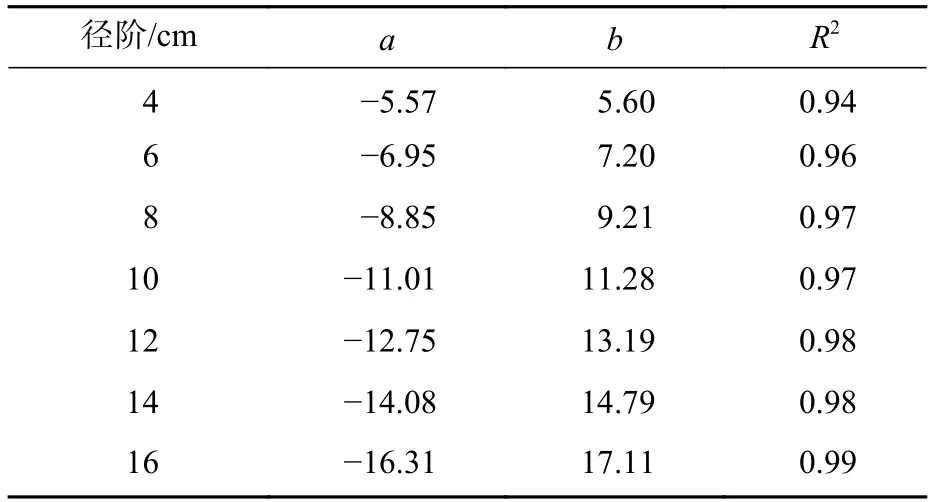
表4 不同径阶毛竹竹节中央直径分布特征拟合结果Table 4 Fitting results of central diameter of node on culm in different diameter class
2.5 毛竹秆形结构因子相关性分析
为分析毛竹秆形结构因子之间的相互关系,选择11个具有代表性的秆形结构因子进行相关分析。由表5可知:竹节数与胸径、竹高呈显著正相关,相关系数分别为 0.667 (P<0.01)和 0.640 (P<0.01)。胸径与1/2高壁厚、1/2高节长呈显著正相关,相关系数分别为0.756 (P<0.01)和0.416 (P<0.01),说明竹子越粗壮,1/2高对应的壁厚越厚、节长越长。胸径与胸高节号呈显著负相关,相关系数为-0.551 (P<0.01),说明竹子越粗壮,达到胸高所需的节数越少,即胸高以下的平均节长越长。竹节数和1/2高节号呈显著正相关,相关系数为0.739(P<0.01),主要原因是竹节长在竹秆上的分布具有对称性(图3)。1/2高节长和最长竹节长的相关系数为0.979 (P<0.01),主要原因是竹节长自基部往梢部有先增加后减少的趋势,且大约在1/2高时竹节最长 (图3)。
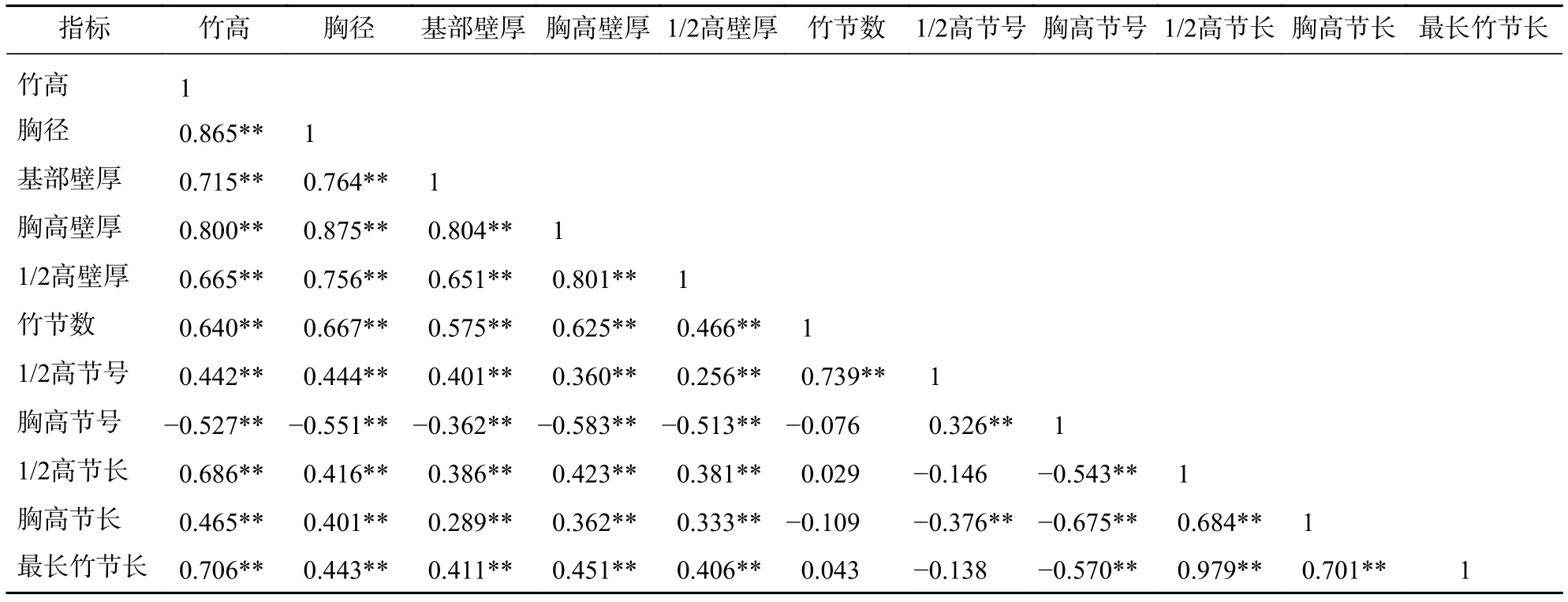
表5 毛竹秆形结构因子的相关性分析Table 5 Correlation analysis of factor of culm form
2.6 毛竹秆形结构的主导因子分析
由表6可知:前4个主因子含有原始数据90.66%的信息,第1主因子解释了秆形结构27.33%的方差,第2、3、4主因子分别解释了秆形结构26.00%、22.64%和14.69%的方差。与第1主因子相关性较大的是1/2高壁厚、基部壁厚和胸高壁厚,而壁厚因子两两之间相关性较高,均达显著水平(P<0.05,表5),表明第1主因子是反映壁厚的综合因子。与第2主因子相关性较大是1/2高节长和最长竹节长,而最长竹节长与1/2高节长之间的相关性达0.982(表5),表明第2主因子是反映1/2高节长的综合因子。与第3主因子相关性最大的是1/2高节号,其次是竹节数,而1/2高节号与竹节数之间的相关性达0.739,表明第3主因子是反映竹节数的综合因子。与第4主因子相关性最大的是胸高节号,其次是胸高节长和胸径,表明第4主因子是反映胸高处秆形结构的综合因子。因子分析结果表明:壁厚因子、1/2高节长、竹节数和胸高处秆形因子是反映毛竹秆形结构的主要因子。
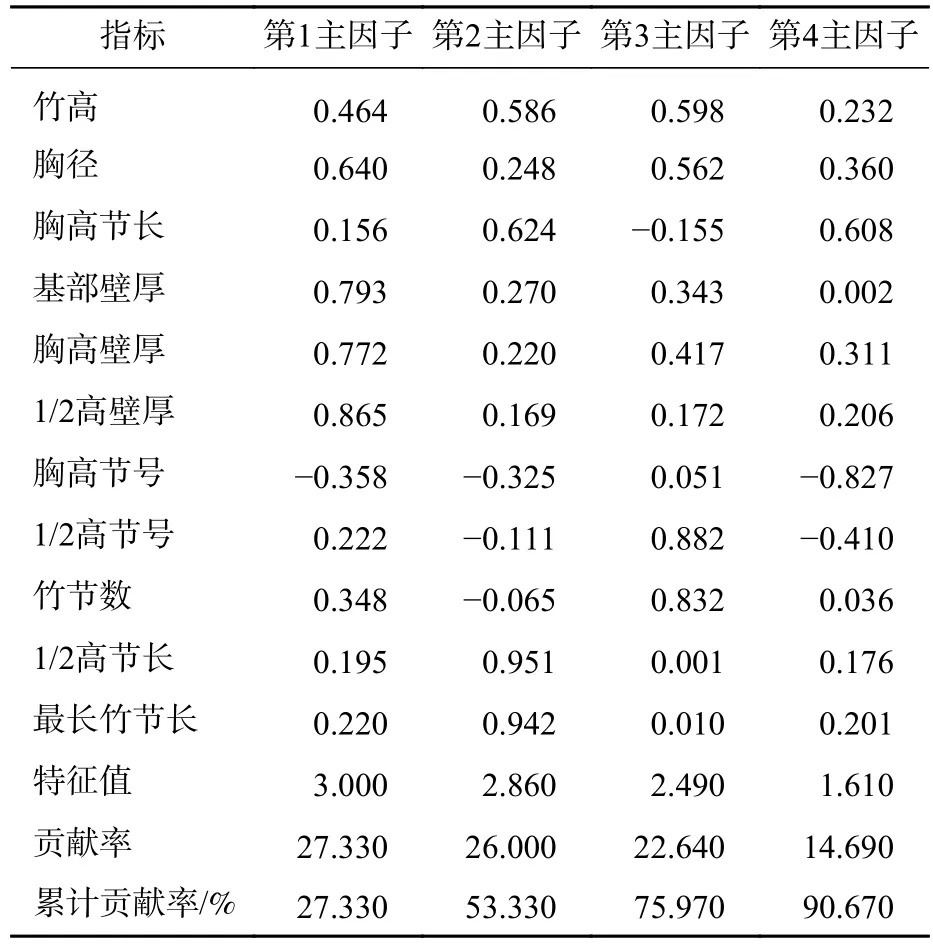
表6 毛竹秆形结构因子分析Table 6 Factor analysis of culm form factor
3 讨论
3.1 毛竹秆形结构的稳定性
稳定性是毛竹秆形结构的特点之一。毛竹的竹节数相对稳定,集中在53~67节,平均59节。抛物线可以较好地拟合竹节长与相对高在竹秆上的分布,但不同径阶毛竹最长竹节的相对高没有明显差异,均约在1/2竹高处。这种稳定性主要是由遗传结构和生长特点决定的[12],竹秆中部居间分生组织较两端分生组织活动期长、生长量多,因此中部竹节节间较长,基部和梢部节间较短[4,13-14]。
3.2 毛竹秆形结构的差异性及其影响因素
差异性是毛竹秆形结构的另一个特点。随着径阶的增大,竹节数和最长竹节长有增大的趋势,这与周芳纯[9]的研究结论一致。胸径越大,胸高以下平均竹节长越长,汪阳东[12]的研究也得到相近的结论。差异性则主要受母竹大小和地理生态因素的影响[15-17]。在出笋前期,母竹消耗了大量的营养物质,导致在出笋中后期的竹笋营养匮乏,生长竞争压力增大[18-20],其生长发育受到一定程度影响。此外,母竹发挥其整合作用,优先将营养供给较大的竹子,较小的竹子由于营养竞争其秆形结构的生长受到了影响[21]。
林木的生长性状不仅与物种遗传特性有关,同时也受环境条件的影响[5,22-23]。在出笋成竹期和孕笋期,气候因子异常,降水量少、气温偏低、寒流持续时间过长等影响竹子的正常生长发育,也会导致毛竹的秆形生长异常,秆形生长量明显变小[16-17,24-25]。本研究方差分析表明:竹节数、1/2高节长、最长竹节长、基部壁厚、1/2高壁厚、高和胸径在10个区域间差异显著。相关分析表明:年均降水量、年均气温与各秆形结构因子均呈正相关,即随着年均降水量和年均气温的增加,毛竹相同位置对应的秆形指标均有逐渐增加的趋势。这与周文伟[26]的研究结论相似,该研究表明:降水量的增加能促进毛竹的生长,水热条件较高的庆元比水热条件较低的安吉的毛竹产量高。实际上,较好的水热条件可以降低毛竹之间的竞争,促进毛竹的生长[16,27-28]。可见,年均降水量和年均气温是影响毛竹秆形生长的重要因素。
4 结论
毛竹竹节数服从正态分布,竹节长与竹节相对高呈抛物线关系,且不同径阶毛竹最长竹节的相对高没有明显差异,均约在1/2竹高处。竹节中央直径与竹节所处相对高呈线性负相关关系。随着径阶的增大,最长竹节长和竹节数有逐渐增大的趋势。壁厚因子、1/2高节长、竹节数和胸高处秆形因子是反映毛竹秆形结构的主要结构因子。
猜你喜欢
课外生活(小学1-3年级)(2022年8期)2022-07-05 18:52:25
作文大王·中高年级(2022年3期)2022-03-21 08:38:12
课堂内外·创新作文小学版(2022年2期)2022-01-28 12:45:37
安徽农学通报(2020年17期)2020-10-09 10:46:23
热带农业科学(2017年9期)2017-10-23 22:56:24
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
农村农业农民·B版(2016年7期)2016-10-21 10:44:23
防护林科技(2016年8期)2016-08-29 01:28:17
贵州科学(2016年2期)2016-06-13 00:58:58
华南农业大学学报(2016年3期)2016-06-01 09:27:59
