
祁连山北坡植物群落空间分布格局与多样性研究
2021-12-17 01:27:04贺国宝
草业学报 2021年12期
贺国宝
(甘肃省肃南县草原工作站,甘肃肃南734400)
山地植被的物种多样性和空间分布格局能够反映植物利用环境资源的状况和种群在群落中的作用与地位[1],也可以揭示植物个体之间相互依存、相互制约的复杂关系[2],对认识山地植物的生态适应性和维持植被群落稳定性机制具有重要的意义[3]。植物群落空间分布格局是群落内部种群个体在空间水平位置的分布模式[4];物种多样性可以表征生态系统内物种组成、结构多样性和复杂程度,同时也是生物群落对生物和非生物环境综合作用的外在体现[5];植物群落中建群种和优势种的分布格局最能代表植被的空间格局变化[6],是研究群落结构、种群间相互作用及种群与环境关系的重要手段。山地植被对土壤属性、温度和水分等外界因子具有很强的敏感性和直观性[7],其中,海拔梯度通过对热量、光照、水分等资源的再分配[8],影响着山地植物的分布范围、群落动态及植被类型,有关海拔梯度下植物群落的多样性[9-10]及物种间和物种内植物功能性状已有大量研究[11-12],但是有关西北干旱区山地植物群落随海拔梯度的分布格局和多样性的研究相对较少。祁连山是祁连山国家公园的主体,也是西北干旱区主要的水源涵养区和重要的生态屏障,高寒草地植被群落的生态变迁对河西走廊生态、文化和经济的发展发挥了极其重要的作用。近年来,学术界对祁连山地东段植物群落特征、土壤性状[13]、地形[14]、气候[15]、坡向[16]等地理环境因子的响应等方面做了大量研究,但海拔梯度下祁连山地北坡植物群落的多样性及分布格局的变化机理尚不明确。鉴于此,本研究以祁连山北麓自然保护区植被为对象,采用TWINSPAN 和DCA 等方法,对植物群落的空间分布及其与环境之间的关系进行分析,揭示植物种、植物群落与环境之间的生态关系,为祁连山北坡高寒植被群落物种多样性保护提供理论依据。
1 材料与方法
1.1 研究区概况
祁连山北坡地处河西走廊与青藏高原的过渡带,具有典型的大陆性气候特征,生态环境脆弱,植被类型以荒漠和草原为主,在特定的地形地貌和气候条件下主要发育并形成了荒漠草原(1600~2450 m)、典型草原(2450~2800 m)和草甸草原(2800~3800 m),植物以旱生和中生草本以及灌木为主。研究区位于祁连山北坡的肃南县大河乡和康乐乡(37°51′36.9″N,101°50′22.9″E),海拔 1600~3500 m,年均温 1~2 ℃,≥0 ℃的年积温为2450 ℃,相对无霜期80 d,年降水量230~450 mm,年蒸发量1480~2120 mm,相对湿度65%,土壤类型主要以亚高山灌丛草甸土、山地灰褐土、山地栗钙土等为主,该区物种丰富,天然植被以草地、灌木林地和乔木林为主,受气候、海拔和坡向的多重影响,有明显的垂直分异规律,主要植物有:盐爪爪(Kalidium foliatum)、合头草(Sympegma regelii)、灌木亚菊(Ajania fruticulosa)、西北针茅(Stipa sareptana)、扁穗冰草(Agropyron cristatum)、鬼箭锦鸡儿(Caragana jubata)、高山柳(Salix cupularis)、嵩草(Kobresia myosuroides)、苔草(Carex tristachya)等。
1.2 试验设计
1.2.1 样地设置 按照祁连山地天然草地类型分布特征,结合多年的野外调查工作经验,从祁连山与河西走廊过渡带至祁连山高山区,在海拔1600~3440 m 内,布设两条间隔5 km 的平行样带,在两条平行样带间海拔每间隔100 m 布设一个10 m×10 m 的样方,共布设32 个样方。对灌木群落进行调查时将灌丛样方划分为4 个5 m×5 m 的灌木样方,记录所有灌木种的种名、高度和盖度等;进行草本植物群落调查时,用交叉对角线法在灌丛样方内选取5 个1 m×1 m 的草本小样方,详细记录每个样方内所有植物的高度、盖度、密度、频度和地上生物量等。用观察法描述样地基本概况,使用GPS 测定样地位置信息。
1.2.2 数据采集 2019 年7-9 月,选择天气状况相对稳定的期间进行取样。首先,在选定的样地中按已划分梯度设置的样方,监测样方内灌木和草本生境状况,调查记载样方内的所有植物种类、数量、盖度、高度、密度、生物量等特征,重复3 次;野外不能鉴别的植物种类,采集标本带回室内鉴定;然后齐地刈割每个草本样方地上所有植株,带回实验室,于80 ℃烘箱中烘干48 h,测量草本样方内物种的地上生物量。
1.2.3 物种多样性分析 选用重要值(important value,IV)来描述物种在群落中的重要性。其计算公式如下[17]:

采用 Shannon-Wiener 多样性指数(H′)、Pielou 均匀度指数(J)和Simpson 优势度指数(C),物种丰富度指数(R)来表示植物个体数量在种间的分配情况。计算公式分别为:

式中:S为植物种数;N为群落中物种总数量之和;Pi表示第i个物种重要值与群落物种总重要值之比[18-19]。
1.3 数据处理
基于野外调查和室内实验获取的植物群落特征和生长环境数据库,采用Wintwins 2.3 软件对所有植被进行TWINSPAN 数量分类;使用Canoco for Windows 4.5 软件进行去趋势对应分析(detrended correspondence analysis,DCA)[20],确定与植被类型相关性最显著的环境因子,用Excel 2016 和Origin 2016 软件绘图。
2 结果与分析
2.1 植物群落类型划分
对祁连山北坡植物群落的32 个样方的重要值矩阵进行TWINSPAN 等级分类(图1)。结合研究区域实际生态意义,采用第5 级的划分结果,将研究区植物群落划分为9 个群丛类型(表1)。该分类结果较为客观地对祁连山北坡植被群落进行分类,比较准确地揭示出植物群落类型与环境梯度之间的关系,同时所划分群落的指示种也较为充分地反映了群落的生境特征。

表1 祁连山北坡9 个植物群丛类型的群落特征及其主要物种成分Table 1 Community characteristics and main species components of nine plant association types on the north slope of Qilian Mountains
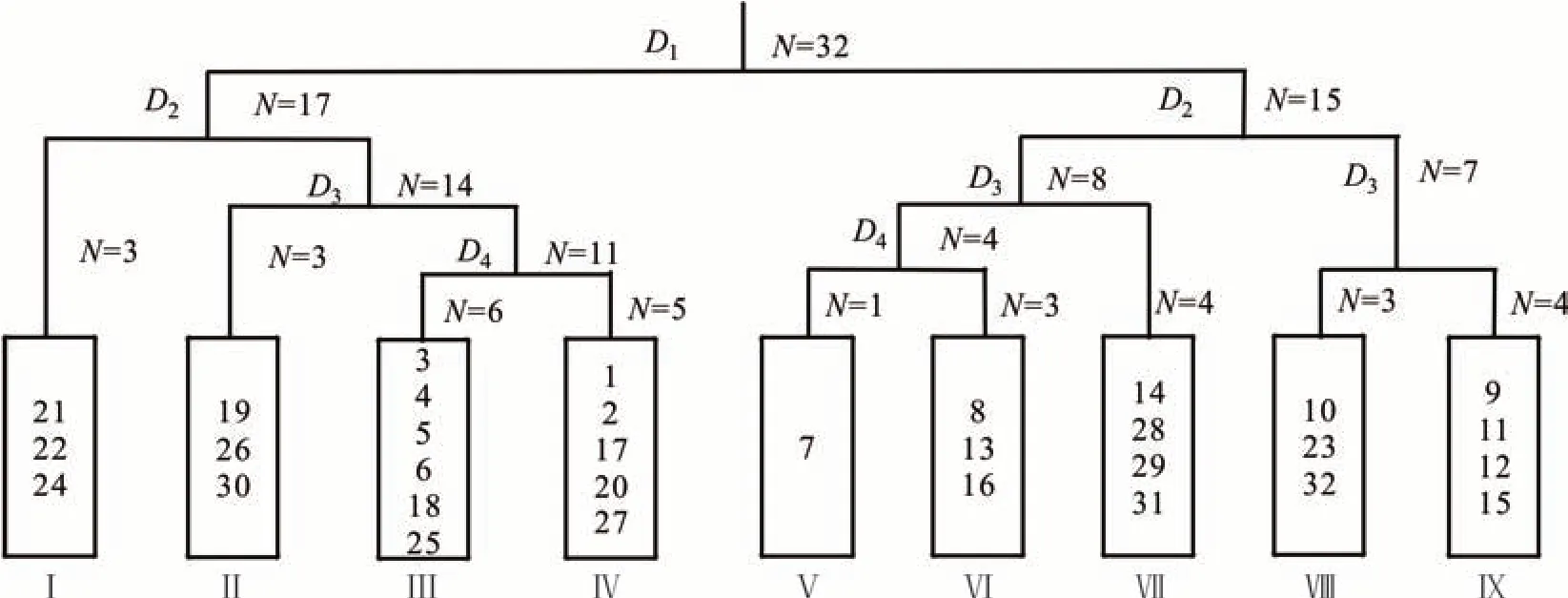
图1 祁连山北坡植物群落32 个样方的TWINSPAN 分类Fig.1 TWINSPAN taxonomic tree of 32 plots of community on the north slope of Qilian Mountain
2.2 DCA 排序
对祁连山北坡 32 个灌丛样方进行 DCA 分析(图 2),4 个排序轴的特征值分别为 0.906、0.372、0.252、0.174,其中第1 轴特征值最大,包含的生态信息量也最多。由TWINSPAN 分类得到的9 个群丛类型在DCA 排序图上的分布更加紧凑和集中,按各群丛生境的不同划分为4 类,对应的植被类型为荒漠、荒漠草原、典型草原和草甸草原,各植被类型之间体现出明确的分布范围和界线,说明DCA 很好地反映出各植物群落之间及群落与环境之间的关系。
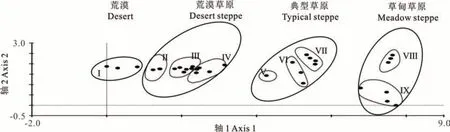
图2 祁连山北坡32 个样方的植物群落DCA 二维排序Fig.2 Two-dimensional DCA ordination diagram of 32 quadrats of community on the north slope of Qilian Mountain
从排序图看,第一轴主要反映各植物群丛所在的海拔梯度,从左到右海拔逐渐升高,群丛类型逐渐由荒漠植被群丛Ⅰ至荒漠草原植被群丛Ⅱ、Ⅲ和Ⅳ过渡为典型草原植被群丛Ⅴ、Ⅵ和Ⅶ,最后演化为草甸植被耐寒灌木群丛Ⅷ和Ⅸ。群丛类型沿第一轴的转换也体现了生境的变化规律,沿第一轴从左向右,植被类型也逐渐由荒漠演化为草甸草原。第二轴基本体现了地形因子的变化,在相同海拔梯度下,群丛Ⅷ排在群丛Ⅸ的上面,表明这些群丛多处于山的阴坡地带,群丛Ⅷ中生长有鬼箭锦鸡儿、高山柳等灌木,而群丛Ⅸ中植物主要为喜好生长在山坡阳面的苔草、嵩草等草甸植被和山沟林下阴湿处的唐松草、嵩草等草本植物。
2.3 植物群落多样性特征
随着海拔由低至高的转变,群丛类型逐渐由盐爪爪+珍珠群丛演化为鬼箭锦鸡儿+高山柳群丛和苔草+嵩草群丛(图3);群落的物种丰富度指数(R)和Shannon-Wiener 多样性指数(H′)整体呈“双峰”变化趋势,Pielou 均匀度指数(J)表现出增减交替趋势,而Simpson 优势度指数(C)呈先降低后增加的趋势。表明海拔梯度一定程度上决定群落内物种组成,在海拔由低向高转变的过程中,植物群落的H′和R表现为荒漠区<荒漠草原区<草甸草原区<典型草原区;植物群落的C为荒漠区>荒漠草原区>草甸草原区>典型草原区。
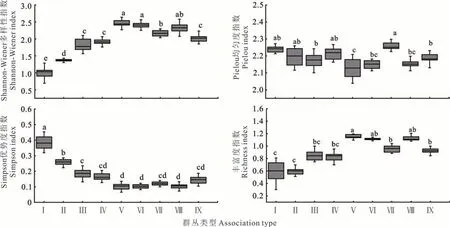
图3 祁连山北坡9 个植物群丛类型多样性指数Fig.3 Diversity index of 9 plant association types on the north slope of Qilian Mountain
3 讨论
物种多样性表征着生物群落和生态系统的结构复杂性,能够体现群落的结构类型、组织水平、发展阶段、稳定程度和生境差异,是揭示植被组织水平的生态基础[21]。海拔是一类特殊的环境因子,反映了水分、温度和光照等因素的综合情况[22-23]。本研究发现,祁连山北坡植物群落可以划分为9 个类群(图1 和表1),随着海拔的升高,群落的R和H′整体呈“双峰”变化趋势(图3),Pielou 均匀度指数(J)表现出增减交替趋势,而Simpson 优势度指数(C)呈先降低后增加的趋势。在海拔对光、热、降水等资源的再分配下,植被类型由荒漠逐渐演化成草甸草原(图2),植物群落的物种数逐渐增加,结构趋于复杂,分布格局发生相应改变。形成这种分布格局的主要原因有:1)群丛I 主要分布在海拔1700 m 以下的荒漠地带,该地区降水稀少,蒸发量大,土壤表层盐碱累积强烈,植被受水盐的双重胁迫,植被盖度最低,发育了以盐爪爪为优势种,茎叶肉质化的耐干旱、耐盐碱的丛生荒漠植被,植物种类稀少,群落结构单一,大多呈斑块分布。因此,群落R和H′较低,C较高。2)群丛Ⅱ、Ⅲ和Ⅳ为半灌木群落,主要分布在海拔1710~2480 m 的荒漠草原地带,植被盖度介于20%~50%,该地区热量充沛,降水稀少,土壤含水量较低,植物生长所需水分主要来自短暂降水、凝结水和地下水;加之高温增强了土壤微生物活性,不利于地表枯落物及根系的腐烂与分解,减慢了土壤有机质的积累速率[24],有机质含量低,土壤相对贫瘠,发育了地下根系发达,茎叶肉质化或表面发育有绒毛适应荒漠生境的灌木亚菊和合头草为优势种的灌木群落;此外,相较于群从I,该区植被生长环境得以改善,植物群落内部成员增加,伴生种开始出现,原来的优势种群生态位被压缩,逐渐被伴生种取代,各种群间生态位发生重叠[25],因此,R和H′逐渐增加,C开始降低。3)群丛Ⅴ、Ⅵ及Ⅶ主要分布在海拔2470~2900 m 处,植被盖度介于60%~100%,群丛Ⅴ的H′和R达到峰值,该地带为典型草原,处于荒漠草原和草甸草原的过渡区,温度适中,降水和光照相对充足,土壤含水量升高,促进了土壤有机质的积累,改善了土壤质地,土壤容重减小,孔隙度增加,增强了土壤涵养水源的能力[26]。因此,发育了以甘蒙锦鸡儿为优势种的喜光灌木群落和以针茅、扁穗冰草以及赖草为主的适应性强的禾本科植物群落,群落结构最复杂,种类丰富,生态位重叠加剧,种间资源竞争激烈,植物群落中伴生种占据了一定位置,使得原有优势种的功能逐渐被弱化,物种的H′,J和R均较高,C较低。4)群丛Ⅷ和Ⅸ主要分布在海拔2760~3440 m 的草甸草原,植被盖度介于90%~100%,该地带属于高海拔地区,降水和光照充足、气温低。严酷的环境过滤使大部分中低海拔植物群落被淘汰,适应耐寒生境的鬼箭锦鸡儿、苔草等成为优势种,群落结构趋于稳定,因此,群落C呈增加趋势(图3)。在山体阴坡发育了高度较大、叶片具绒毛,花具苞片等保温结构的鬼箭锦鸡儿+高山柳耐寒灌木群丛;在阳坡发育了以苔草、嵩草为主的草甸植被,植被盖度最大,植被呈镶嵌结构分布。海拔影响下,植物群落的分布格局发生了改变,由聚集的斑块状分布逐渐演化为成群片状分布,均匀度上升,最后演化为镶嵌分布的草甸群落,均匀度下降,所以群落的J则表现出增减交替的现象(图3)。祁连山高寒草地植物群落H′,随C的降低而增加,即H′越高,C越低,群落的结构越复杂,组成群落的植物种类和个体数越多,分布越不均匀,这与冯刚等[27]的研究结果一致,体现了海拔对山地植物群落多样性和群落分布格局的影响和山地植被对环境的生态适应机制。
4 结论
随着海拔的升高,生境类型由荒漠逐渐过渡为草甸草原,植物群丛类型由优势度指数高、群落结构单一的荒漠植被盐爪爪+珍珠群丛逐渐演化为多样性指数和丰富度指数高的草甸植被鬼箭锦鸡儿+高山柳群丛和苔草+嵩草群丛,体现了山地生境对植物群落构建的过滤作用。本研究仅从海拔的角度出发,通过分析海拔梯度对群落分布及多样性的影响,探讨了祁连山植物对海拔的适应性。植物群落的分布格局及多样性也受气候、水文、土壤环境及其他群落环境影响,需要进一步探讨和研究。
猜你喜欢
发展(2024年1期)2024-04-25 03:45:30
中国水土保持科学(2023年6期)2024-01-05 05:53:12
水土保持研究(2022年1期)2022-12-27 07:35:24
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
读者·原创版(2021年2期)2021-02-26 10:12:21
林业调查规划(2020年3期)2020-06-03 07:02:36
华人时刊(2016年17期)2016-04-05 05:50:36
黄河之声(2016年20期)2016-02-21 11:55:33
