
环境因子对云南扁穗雀麦种子萌发和出苗的影响
2021-12-17 01:27:00张敏NIPAPANKanjana李铷傅杨汤东生
草业学报 2021年12期
张敏,NIPAPAN Kanjana,李铷,傅杨,汤东生
(云南农业大学云南生物资源保护与利用国家重点实验室,云南昆明650201)
雀麦(Bromusspp.)属于禾本科(Poaceae)多年生或一年生草本植物。据中国植物志记载,全世界雀麦属植物有约250 种,我国约有70 多个种。雀麦分布极广,遍布于世界温带和热带高海拔地区,为天然草地和人工牧场中利用价值高的牧草资源[1]。由于雀麦具有抗旱、抗寒和抗病虫等优良特性,近年来随着我国畜牧业的快速发展,无芒雀麦(B. inermis)[2-5]、扁穗雀麦(B. catharticus)[3,6-8]和旱雀麦(B. tectorum)[4,9-10]等牧草种类在我国已广泛种植。牧草逃逸到农田就可能会变成杂草。雀麦入侵到农田成为危害性杂草的报道可追溯到100 多年前由欧洲传入到美洲的旱雀麦[11]。从20 世纪50 年代开始,全球有关杂草雀麦的报道越来越多,报道最多的仍是旱雀麦,双雄雀麦(B. diandrus)[12-13]、硬雀麦(B. rigidus)[14]、贫育雀麦(B. sterilis)[15]、无芒雀麦[16]等种类也有报道。对雀麦杂草的研究内容涉及生物学[16-18]、危害性[19-20]、竞争性[21-23]、防治[24-26]、抗药性[27-29]等。我国自 1986 年报道雀麦危害[30]以来,在我国北方农田传播扩展蔓延很快,成为麦田重要的恶性杂草。据维基百科报道,由于雀麦属植物与小麦族(Triticeae)小麦属(Triticum)和大麦属(Hordeum)植物有很相近的亲缘关系(linkage),在抽穗之前与大、小麦极其相似,在田间一般不易被发现,很难防治。扁穗雀麦曾作为一种优质牧草在云南广泛种植,由于云南大多数区域低纬度高海拔产生的温暖气候使扁穗雀麦表现周年生长繁殖的现象,近年来在绿化带、次生荒地、田间快速扩张繁殖,成为潜在田间恶性杂草。其外形与麦类作物高度相似,防治比较困难,因此有必要开展雀麦的生物生态学研究,旨在为扁穗雀麦的精准防治措施制定科学依据。
种子萌发是杂草入侵农田和危害作物的前提条件。气候、土壤等因子均会对杂草种子萌发产生影响。Beckstead 等[31]的研究发现,采自5 个不同区域的旱雀麦种子均能在5/15 ℃至20/30 ℃昼夜温差的温度环境中萌发。也有研究表明,15 ℃黑暗条件下培养的贫育雀麦种子发芽率高,而红光处理却抑制种子的萌发[32]。还有研究表明,提高空气中的CO2浓度也可提高直立雀麦(B. erectus)种子萌发率[33]。虽然也有少量文献报道了部分环境因子对扁穗雀麦种子萌发的作用[2-5],但这些研究均只是部分揭示环境因子对种子萌发的效应,缺乏系统性的研究。而云南这种高海拔低纬度生长的扁穗雀麦群体种子的萌发规律是否具备区域性、独特性有待明确。本研究拟从杂草生态学的角度,研究温度、湿度、pH、盐、渗透势等环境因子对扁穗雀麦种子萌发的影响规律,旨在为扁穗雀麦的综合防治措施的制定提供理论依据。
1 材料与方法
1.1 材料采集
扁穗雀麦种子于2015 年9 月采集于云南省昆明市云南农业大学后山荒地,分批次收集成熟发黄的穗子带回实验室,自然风干1 个月左右,而后去除颖壳,混合后装袋,室温保藏2 个月破除休眠后进行试验。
1.2 试验方法
1.2.1 温度试验 将购置的石英砂(粒径≤2 mm 用清水冲洗3 遍,在120 ℃烘箱中烘足3 h,冷凉后精确称取60 g 石英砂铺于直径为9 cm 的培养皿中,铺设厚度约5 mm,各处理精选30 粒籽粒饱满的种子均匀地置于砂上,每皿加灭菌纯水15 mL,使水面刚好没过砂面,盖上培养皿,并用保鲜膜封好置于人工智能培养箱中培养。培养条件为12 h 光照/12 h 黑暗,光照强度为2400 lx。种子萌发以胚根突破种皮1 mm 为统计标准,逐日统计发芽数直至第10 天,计算萌发率和萌发势。温度设置5 个恒温处理水平即30、25、20、15、10 ℃和10 个变温处理水平即30/25、30/20、30/15、30/10、25/20、25/15、25/10、20/15、20/10、15/10 ℃。在变温处理情况下,高温与光照处理相对应,低温与黑暗处理相对应。4 次重复。
1.2.2 pH 试验 培养条件基本同温度试验,只是将水换成不同pH 值的溶液,温度设置为恒温25 ℃。用于培养种子的 pH 值缓冲液配制参照 Chachalis 等[34]的方法。即用 1 mol·L-1HCl 调节 2 mmol·L-1邻苯二甲酸氢钾配制 pH 为 4 的缓冲液;用 1 mol·L-1NaOH 调节 2 mmol·L-1的 MES[2-(N-吗啉)乙磺酸]配制 pH 为 5 和 6 的缓冲液;用1 mol·L-1NaOH 调节2 mmol·L-1HEPES[N-(2-羟甲基)哌嗪-N′-(2-乙磺酸)]配制pH 为7 和8 的缓冲液。用 1 mol·L-1NaOH 调节 2 mmol·L-1三甲基甘氨酸配制 pH 为 9 和 10 的缓冲液。
1.2.3 耐盐性试验 基本培养条件同温度试验,只是将培养溶液换成不同浓度的NaCl 溶液。NaCl 的浓度设置为 0、10、20 、40、80、160、320、500 mmol·L-1[35]。恒温 25 ℃培养。
1.2.4 耐旱性试验 基本培养条件同温度试验,只是将培养溶液换成不同渗透势的PEG 8000 溶液。参照Chauhan 等[36]的方法配制渗透势为0、-0.1、-0.2、-0.4、-0.6、-0.8、-1.0 MPa 的溶液。恒温25 ℃培养。
1.2.5 土壤湿度试验 将田土风干磨碎,过2 mm 筛,在120 ℃烘箱烘干至恒重。将筛后的土壤混合均匀后倒入塑料杯,土面距杯口2 cm,分别配制100%、90%、80%、70%、60%、50%、40%相对含水量的土壤。将加足水的杯子用封口膜封好后静置12 h,均匀播入种子30 粒,播种深度为1 cm。用封口膜密封杯口,然后将放有种子的杯子置于人工智能培养箱中恒温25 ℃培养,每日记录出苗数。其他设置同温度处理试验。
1.2.6 播种深度试验 取上述烘干的土壤,将筛后的土壤混合均匀后倒入高12 cm 的塑料杯中。设置0、2、4、6、8、10 cm 共6 种播种深度。将底层土铺好后,均匀播入30 粒种子,盖上不同厚度的细土。土壤含水量控制在60%,然后用封口膜封住杯口,置于恒温25 ℃人工智能培养箱中培养,每日记录出苗数,连续3 d 不再出苗为止。其他设置同温度处理试验。
1.3 数据处理

采用Excel 2010 进行试验数据的初步处理,采用SAS 9.4 软件分析各处理之间的差异显著性(Duncan’s 方差分析法),种子最终萌发率和累积萌发率对各处理因素的响应规律采用SigmaPlot 14 软件绘图,并拟合盐度、渗透势、深度对种子最终萌发率的回归模型。
2 结果与分析
2.1 扁穗雀麦种子萌发对温度的响应
在恒温条件下,25 ℃处理的种子发芽率高达89%;温度增加到30 ℃,种子萌发率显著下降;而当温度下降至20 ℃,种子萌发率从90%左右下降到50%;而当给予10 ℃的恒温处理后,种子几乎不能萌发。在大多数情况下,变温处理促进种子萌发(图1)。在白天30 ℃的高温环境下,随着昼夜温差扩大,种子萌发率呈先增加再降低的趋势,30/20 ℃的种子萌发率最高达到92.1%,恒温30 ℃萌发率最低。在白天为25 ℃的条件下,种子萌发率随昼夜温差的扩大而降低。而白天温度在20 ℃情况下,20/15 ℃的变温条件种子萌发率最高。白天温度低于20 ℃以后,种子萌发率低于40%。
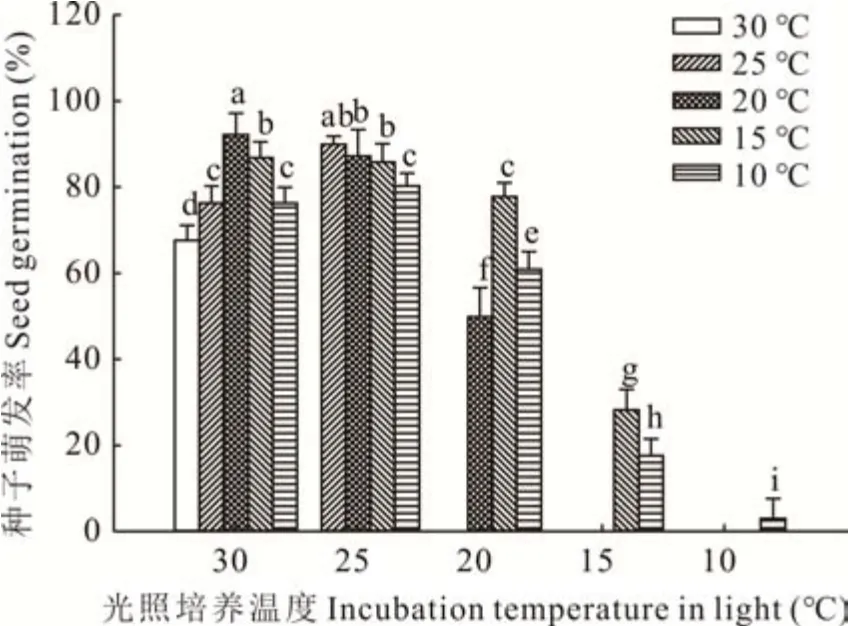
图1 不同培养温度条件对扁穗雀麦种子萌发的影响Fig. 1 Effect of incubation temperature conditions on seed germination of B. catharticus
2.2 扁穗雀麦种子萌发对pH 值的响应
扁穗雀麦在pH 4~10 的环境下,种子萌发率均高于90%。种子萌发率和萌发势在弱酸和弱碱环境下要显著高于中性环境(图2)。种子在pH 为6 时萌发率最高,达到98.3%,但与种子在pH 为5、9、10 的处理之间没有显著差异性。当pH 低至4 时,种子萌发率最低,为90.2%。在pH 为9 和10 的情况下,扁穗雀麦种子保持着高达96.0%的萌发率。同时,种子的萌发势差异更大,种子在pH 为7 时,萌发势不到80%,而pH 为9 的处理萌发势已经超过90%。由此可见,种子萌发率高的处理,其种子萌发势也高。可以推知,扁穗雀麦对酸碱度有广泛的适应性,最适条件为弱酸环境,但也对碱性环境表现出极高的适应性。
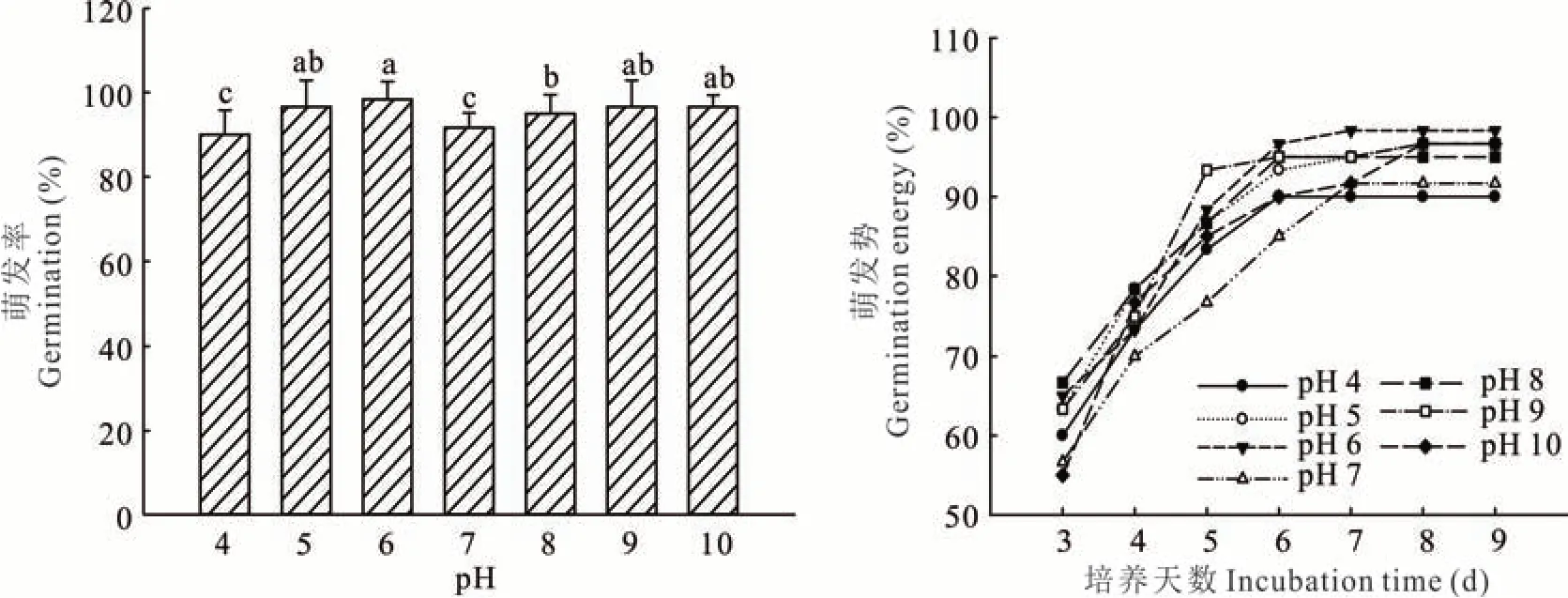
图2 pH 对扁穗雀麦种子萌发的影响Fig.2 Effect of incubation pH on seed germination of B. catharticus
2.3 扁穗雀麦种子萌发对NaCl 的响应
种子在NaCl 浓度≤20 mmol·L-1,种子萌发率高于95%;在浓度为80 mmol·L-1时,种子萌发率高于90%;在浓度为160 mmol·L-1时,种子萌发率高于85%;在浓度320 mmol·L-1时,种子萌发率高于50%;NaCl 在浓度500 mmol·L-1时方能完全抑制种子萌发(图3)。经回归分析,NaCl 对种子的萌发影响规律符合三参数S 型曲线模型{y=97.96/[1+exp(x+212.0)/83.25],R2=0.99}。种子萌发势的规律基本同于种子萌发率。NaCl 的浓度越低,种子的萌发势越高,尽管在低浓度下各处理的最终萌发率差异不显著。由此可见,扁穗雀麦种子萌发对盐胁迫表现出较高的适应性和耐受性。

图3 NaCl 对扁穗雀麦种子萌发的影响Fig.3 Effect of NaCl on seed germination of B. catharticus
2.4 扁穗雀麦种子萌发对渗透势的响应
随着培养液渗透势的降低,扁穗雀麦种子的萌发率和萌发势均呈下降的趋势(图4)。对照和-0.1 MPa 的处理,在培养6 d 后,种子萌发率基本接近最终萌发率;而渗透势低的处理,种子萌发率一直呈缓慢增长的过程。经回归分析,渗透势对种子萌发的影响规律符合三参数S 型曲线模型{y=96.02/[1+exp(-x-0.88)/0.15],R2=0.99}。根据这个模型,50%抑制种子萌发率的渗透势为-0.98 MPa。显然,扁穗雀麦在渗透势极低的土壤环境中具有萌发的能力。由此可见,扁穗雀麦有较高的抗胁迫能力。
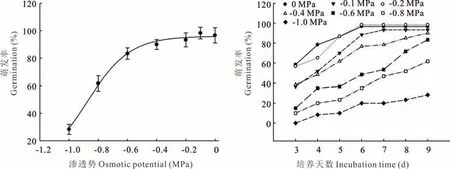
图4 渗透势对扁穗雀麦种子萌发的影响Fig.4 Effect of osmotic potential on seed germination of B. catharticus
2.5 扁穗雀麦种子萌发对土壤湿度的响应
随着土壤含水量的增加,种子萌发率和萌发势均呈先增大后减少的趋势(图5)。0.6 相对含水量的处理最终种子萌发率最高,达到100%。1.0 相对含水量处理的种子完全不能萌发。萌发率高的处理,在种子培养5 d 后,种子的萌发率基本接近最终萌发率。对数据进行回归分析发现,土壤相对湿度对种子萌发的影响规律符合二项式曲线回归模型(y=-198.09+948.17x-736.11x2,R2=0.94)。根据这个回归模式,种子萌发率最高时土壤相对湿度为0.63,与相对湿度0.6 的结果非常接近。0.4 的相对湿度对大多数植物种子几乎难以萌发的情况下,扁穗雀麦种子萌发率超过60%;而饱和含水量的处理种子不能萌发。以上结果表明,扁穗雀麦更适应干旱而不是湿润的土壤条件。

图5 土壤湿度对扁穗雀麦种子萌发的影响Fig.5 Effect of soil humidity(RH)on seed germination of B. catharticus
2.6 扁穗雀麦出苗对播种深度的响应
在土壤表面培育的种子萌发率最高达100%,随着播种深度的增加,扁穗雀麦的出苗率逐渐降低,但0~4 cm的处理之间差异不显著,播种10 cm 的处理种子几乎不出苗(图6)。对数据进行回归分析发现,土壤深度对种子萌发的影响规律符合三参数S 型曲线模型为y=97.15{1+exp[(x-7.81)/0.78]},R2=0.99。根据这个模型,抑制种子出苗率50%的土壤深度为8.58 cm。种子在土壤中培育4 d 后,播种深度为0 cm 的处理有48%的出苗率,播种2 cm 的处理有78%的出苗率,其他播种深度均未出苗。各处理首次出苗的时间随着播种深度的增加而延后。培养结束时,将播种深度为10 cm 的处理倒出,发现土壤中萌发的种苗已腐烂。可见10 cm 的播种深度不能满足扁穗雀麦的出苗条件。

图6 土壤深度对扁穗雀麦种子出苗的影响Fig.6 Effect of soil depth on seedling emergence of B. catharticus
3 讨论与结论
扁穗雀麦是云南近年来发展较快的入侵杂草。适应性强是入侵杂草的显著特征之一。而对各因子响应的生态幅度是量化其生态适应性的重要指标。种子萌发是杂草入侵的第一步,萌发率和萌发势的强弱对其建立种群和危害作物起决定作用。种子萌发对土壤温度、湿度、埋藏深度、pH、盐浓度和渗透势的响应是常用描述杂草种子适应性的主要指标。本研究表明,日均温度在15~30 ℃的昼夜温差环境中,种子有超过50%的萌发率,恒温条件下25 ℃种子萌发率最高,30℃种子萌发率下降到不足70%。本研究表明,在土壤相对含水量40%~80%、土壤深度低于6 cm、pH 介于4~10 时扁穗雀麦种子均可保持60%以上的萌发率。土壤盐浓度低于160 mmol·L-1和渗透势低于-0.4 MPa 的处理时,雀麦种子萌发均可达到80%以上。
植物生长对环境因子的响应规律是分阶段的,种子萌发所需的最适温度往往不同于苗期所需。田宏等[3]的研究表明,扁穗雀麦种子在25~35 ℃恒温条件下发芽率均超过96%,远高于本研究的萌发率,但最适萌发温度为25 ℃,本研究与之相同。本研究表明变温处理种子萌发率更高,与Ahshallash[37]的研究趋势基本一致。周志彬等[10]研究了温度对旱雀麦、扁穗雀麦、无芒雀麦种子萌发的影响,但仅设定了5、15 和25 ℃三种温度处理条件,得出高温处理有利于种子萌发的结论;本研究结果也支持在10~25 ℃,高温更有利于扁穗雀麦种子的萌发。有研究表明,旱雀麦种子在恒温条件下比变温条件下种子萌发率更高,这与许多植物种子萌发规律相背[38]。在大多数情况下,光照处理通过刺激生长物质的合成促进种子的萌发。本研究表明光照处理对扁穗雀麦种子萌发无抑制作用,但有研究表明光照处理却抑制贫育雀麦和旱雀麦种子萌发。本研究表明扁穗雀麦种子在土壤表面的种子萌发率最高,而随着种子深度的增加而降低。Andersson 等[39]的研究却是土壤表面的种子比埋藏在土壤的种子更受抑制,这说明贫育雀麦和旱雀麦是厌光型种子,与扁穗雀麦差异巨大。进一步研究表明光照对贫育雀麦种子的抑制程度与温度有关[40]。光照和温度是影响种子萌发最基本的环境要素,以上研究结果表明雀麦不同种间甚至同种不同群体之间对温度和光照响应差异大。事实上有证据支持采自不同区域不同群体的旱雀麦种子萌发对环境的响应差异较大,并推断这种较大的差异可能与居群和母株植物的生长环境有关[41-42]。Li 等[43]的研究发现,普通雀麦(B. japonicus)在pH 5~10 有超过80%的萌发率,土壤渗透势在-0.6 MPa 和盐浓度达到140 mmol·L-1左右,种子萌发率也能达到80%,本研究与其基本一致。Tavili 等[44]的研究表明,Bromus tomentellus和贫育雀麦对盐和渗透势均有较高的耐受性,申忠宝等[45]的研究表明,80 mmol·L-1混合盐处理,无芒雀麦的种子萌发率仍达80%左右,胡生荣等[46]的研究表明,1.5%的NaCl 可完全抑制本地和引进无芒雀麦的群体种子的萌发。王莹等[47]的研究表明,无芒雀麦在5 种禾本牧草中抗旱性是最强的,可耐受-0.6 MPa 的渗透势。尽管不同群体的雀麦对光照和温度的响应差异巨大,但前人和本研究均表明雀麦种子萌发具有较高的抗胁迫能力,种间和居群之间的差异不大。雀麦是世界范围内重要的杂草,尽管上述研究得到种子萌发的部分规律,但从目前公开报道的文献来看,对各种雀麦种子萌发生态学的研究报道模棱两可,仍缺乏系统性的工作。
雀麦属于冷季草系,与小麦(Triticum aestivum)、大麦(Hordeum vulgare)和燕麦(Avena sativa)同属于小麦簇。雀麦属植物对人类具有重要经济价值,可用作牧草、也有少数种被用作草坪草、观赏植物、防止水土流失的植物等。当前我国外来恶性杂草中,大多数是被作为有用植物引入后逃逸到野外而成为杂草。人们在种植雀麦的过程中必须防治其逃逸,因为大多数雀麦具有快速扩张、演变为入侵杂草的潜力,如旱雀麦成为美国西北部严重的恶性杂草[48]。在欧洲,黑麦状雀麦(B. cecalinus)和粗雀麦(B. grossus)成为黑麦(Secale cereale)田、B. bromoideus成为小麦田的重要田间杂草[49]。目前国内外很少有关于扁穗雀麦作为入侵杂草的报道。扁穗雀麦起源于地中海,在我国引种后作为北亚热带和暖温带地区的牧草种植,本研究结果证明这种植物对生态环境有广阔的适应性,具备入侵杂草的典型特征。当前我国城市建设速度很快,城市绿化需要从野外荒地取土壤种植景观植物,保留在土壤中的雀麦种子也被带入到人类的生活区,这也许是扁穗雀麦由牧草变成农田和环境杂草的重要原因。雀麦属包含几十个种,在国外多个种已经入侵农田成为田间恶性杂草;而我国目前还缺乏有关全国麦区不同雀麦作为潜在杂草的风险预警研究。由于雀麦与麦类作物高度的形态相似性,植保工作者很难在苗期区分,人工防治和化学防治都较为困难。因此,今后国内同行要积极行动起来,细致地开展我国农田杂草雀麦不同种的分类学调查、分布情况、危害状况等生物生态学研究,为我国雀麦的防控和风险预警做出贡献。
猜你喜欢
科教新报(2022年22期)2022-07-02 12:34:28
Rice Science(2021年4期)2021-07-13 10:00:54
河北农机(2020年10期)2020-12-14 03:13:42
河北省科学院学报(2020年4期)2020-03-19 02:02:40
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
第二课堂(小学版)(2018年6期)2018-07-09 01:06:46
大陆桥视野·下(2017年11期)2017-10-31 11:02:24
电子制作(2017年24期)2017-02-02 07:14:16
快乐语文(2016年10期)2016-11-07 09:44:51
现代农业(2015年5期)2015-02-28 18:40:49
