
中国牧草育种中的若干科学问题
2021-12-17 01:27:04刘志鹏周强刘文献张吉宇谢文刚方龙发王彦荣南志标
草业学报 2021年12期
刘志鹏,周强,刘文献,张吉宇,谢文刚,方龙发,王彦荣,南志标
(兰州大学草地农业科技学院,兰州大学草地农业生态系统国家重点实验室,甘肃兰州730020)
牧草(grass 或herbage)是饲用植物的总称,以草本植物为主栽培的或野生的饲用植物,包含可供饲用的半灌木、灌木、小乔木。牧草具有改良土壤理化性状、维持土壤肥力、防风固沙、保持水土、绿化环境和调节气候等生态作用[1]。牧草富含各种营养物质和粗纤维,是粮食、青贮或其他饲料不能替代的。发展草业是我国农业供给侧结构性改革的重要内容之一,也是我国“生态优先、绿色发展国家战略”的重要保障,对于落实“绿水青山就是金山银山”和“山水林田湖草沙生命共同体”理念具有重大战略意义。草地占全球陆地面积的54.3%,中国的草地面积占国土面积的27.6%(草地面积是耕地面积的2.1 倍,与林地面积相当),不论是全球还是国内,草地都是最大的陆地生态系统。目前,我国80%以上的草地都出现不同程度的退化,亟须补播改良,以便恢复天然草地生态系统。因此,牧草不仅是我国草食畜牧业发展的基石,也是我国退化草地补播改良的物质基础,影响着国家的食品安全和生态安全,决定着生态与生产的协调发展。
1 我国牧草育种的现状与挑战
1.1 干草和草种依赖进口
据估计,我国未来每年牧草饲料需要量将达到1000 万t[2]。而国产干草存在数量少、品质低等问题,不能满足我国畜牧业发展的需要,因此,亟须通过国外进口的方式改善我国牧草产能不足的局面。目前,我国进口的干草主要包括苜蓿(Medicago sativa)和燕麦(Avena sativa)[3]。优质苜蓿指达到《苜蓿干草捆》(NY/T1170-2020)二级及以上的苜蓿干草及其草产品,在我国自给率只有64%,需要从美国(约90%)和西班牙进口。2008、2012、2015和2020 年我国优质苜蓿进口量分别是1.9、46.0、120.0、135.9 万t(5.6 亿美元)。至于燕麦草,则全部从澳大利亚进口[4-5],2008、2013、2015 和2020 年,燕麦草的进口量分别是0.2、4.3、15.2、33.5 万t(1.2 亿美元)。近5 年,我国每年草种子需求量在15 万t 左右,每年进口草种子5 万t 左右,1/3 依赖进口,其中草坪草种子进口量几乎达90%。2020 年1-11 月,我国草种子进口5.98 万t(1.0 亿美元)[5]。在新冠疫情持续发展和中外经贸摩擦等背景下,我国干草和草种国外依赖性过高,缺乏主动权,已成为农业发展的“卡脖子”问题。
1.2 牧草品种数量较少
从1987 年全国牧草品种审定委员会成立以来,共审定草品种636 个,其中苜蓿属(Medicago)牧草114 个,紫花苜蓿品种87 个,杂花苜蓿品种20 个,其他苜蓿7 个,包括黄花苜蓿(M. falcata)、天蓝苜蓿(M.lupulina)、花苜蓿(M. ruthenica)和金花菜(M.polymorpha),而同期美国和欧洲国家登记的牧草品种多达5000 多个。以紫花苜蓿为例,2016-2020 年我国共审定28 个品种(年均5.6 个),同期北美苜蓿改良协会(NAAIC)共审定登记354 个品种(年均70.8 个)[6]。目前,我国紫花苜蓿种子仍有80%以上依赖进口,亟须培育出一批具有自主知识产权且适合我国本土生长的优良牧草新品种。
1.3 牧草育种基础研究发展迅速
近年来,我国牧草育种基础研究发展迅速,我国科学家陆续解析了国际为数不多的牧草基因组,如鸭茅(Dactylis glomerata)[7]、紫 花 苜 蓿[8-10]、无 芒 隐 子 草(Cleistogenes songorica)[11]、长 穗 燕 麦 草(Thinopyrum elongatum)[12]、象草(Cenchrus purpureus)[13]、金花菜[14]和花苜蓿[15-16]等。以紫花苜蓿文章数量为例,在“Web of Science”搜索“题目”包含“Medicago sativa”“alfalfa”或“lucerne”的文章,比较了 1990-2020 年中国、美国和加拿大三国发表的文章数目,总数分别是1197、3622 和1016 篇。近30 年,美国发表文章最多,中国和加拿大接近。2005年之前,我国在紫花苜蓿研究领域文章稀少,接近空白,美国在该领域发表的文章最多;2016 年之后,年度发表文章数量我国跃居第一;2020 年,我国发表的紫花苜蓿文章数量遥遥领先。由此可见,我国牧草育种的基础研究虽然起步晚,但发展非常迅速(图1)。
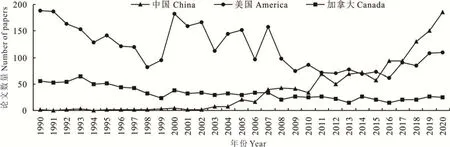
图1 1990-2020 年中国、美国和加拿大发表的紫花苜蓿论文数量比较Fig.1 Comparison of the number of alfalfa papers published in China,the United States and Canada from 1990 to 2020
1.4 前景严峻
预计到2030 年,我国人口将达到16 亿,口粮和饲料粮的产量都要增加,在耕地面积不变的前提下,粮食单产仍需提高1/3 以上,目前我国粮食产量的增长速度远低于人口增长的速度。在“谷物基本自给,口粮绝对安全”的方针指导下,大部分牧草只能种植在中低产田、边际土地或不适合农作物生长的土地。因此,我国培育的牧草新品种在抗逆、优质、高产等方面要更加突出,相关基础研究要全面开展。
然而,我国牧草种类繁多,天然草原上就有6000余种饲用植物[17],国家林业和草原局 2021 年 7 月公布的中华人民共和国主要草种目录也包含了120 个物种。据“Web of Science”统计,2001-2020 年“题目”包含“水稻(Oryza sativa)”“小麦(Triticum aestivum)”“玉米(Zea mays)”“大豆(Glycine max)”和“紫花苜蓿”分别检索到 82526、72480、69319、41327 和 7165 篇论文,我国科学家发表的文章数目分别为23242、14026、12499、6529 和 1178 篇。由此可见,我国科学家在水稻、小麦、玉米和大豆发表的文章数量分别是紫花苜蓿的 20、12、11 和 6 倍(图 2)。“牧草之王”尚且如此,其他主要牧草如羊草(Leymus chinensis)、黑麦草(Lolium perenne)、白三叶(Trifolium repens)等的研究就更加少了。

图2 2001-2020 年紫花苜蓿与其他农作物论文数量比较Fig. 2 Comparison of the number of papers on alfalfa and other crops from 2001 to 2020
我国科学家高彩霞研究员在Cell 发表综述论文,将农作物的育种方法总结为杂交育种、突变育种、转基因育种和基因编辑育种4 个发展阶段[18]。截至目前,绝大多数牧草尚处在杂交育种或者驯化育种阶段,远落后于主要农作物。因此,牧草育种意义重大,虽然前景严峻,但是发展空间广阔。
2 牧草育种的关键瓶颈
2.1 种质资源
20 年前,模式植物必须满足3 个条件:即突变体库、基因组并具备高效遗传转化体系,当时最符合条件的植物是拟南芥(Arabidopsis thaliana)和水稻。时至今日,满足这3 个条件的植物越来越多,如玉米、小麦和蒺藜苜蓿(M. truncatula)等,有了这3 个平台,就可以从正向遗传学或反向遗传学对植物的重要性状进行深入的解析。但是,对于牧草,绝大多数物种还不具备这3 个条件,严重地制约着牧草育种的快速发展。
我国的饲用植物涵盖5 门246 科1545 属,共计6704 个物种,近20 年来保存种质55811 份,但是如果平均到物种,数量上则远低于国内的水稻、小麦、大豆等种质资源,还应继续大规模收集,仔细地摸清家底。尤其是随着草地过度放牧、气候变化、城镇化、土壤污染、采矿、外来物种入侵等因素影响,很多濒危种质、天然突变种质面临丧失,进而优异基因资源面临灭绝。牧草育种者应该进一步加强野生种质资源的收集、保护、系统评价以及共享,尤其是对重点种质应加大保护和收集力度,如苜蓿属、野豌豆属(Vicia)、赖草属(Leymus)、披碱草属(Elymus)等,具体见陈志宏等[19]的研究,将来通过高通量测序方法厘清物种形成机制,解析属内系统进化特性。
对于牧草育种而言,种质资源不但包括野生种质,还包括人工构建的突变体库、各种遗传群体。可以利用EMS 诱变、快中子诱变、航空诱变、反转座子插入等多种方法构建主要牧草的突变体库,为通过反向遗传学的方法定位控制重要农艺性状的关键基因奠定基础。此外,还可以通过人工杂交和自交等方法,构建杂交群体、回交群体,将来用于高密度遗传图谱构建,也用于重测序定位目标基因。
2.2 组学数据
任何一个物种基因组的首次破译,对该物种的基础研究而言都具有划时代意义,会为人类研究该物种开辟出新的分子生物学工具,如物种特异性分子标记开发、基因克隆、代谢通路、基因定位、遗传图谱构建、基因育种芯片、基因组选择、基因家族鉴定等,可为该物种的品种选育、功能基因发掘、品种鉴定、多样性保护等提供有力保障。
目前,四倍体紫花苜蓿已组装了2 套基因组,分别使用了育成品种“中苜1 号”和地方品种“新疆大叶”,但是这还远远不够。首先,组装质量还有待于进一步提高。由于紫花苜蓿是同源四倍体、自交不亲和、重复序列多,没有两粒紫花苜蓿种子的基因组是一致的。再者,紫花苜蓿分类复杂,最新的研究结果表明紫花苜蓿复合体包括6 个类群[20]:M. sativasubsp.caerulea(2n=16)、M. sativasubsp.sativa(2n=32)、M. sativasubsp.×vari(2n=16,32)、M. sativasubsp.falcatavar.falcata(2n=16,32)、M.sativasubsp.falcatavar.viscosa(2n=16,32)、M.sativasubsp.glomerata(2n=16,32)。这些同倍性的亚种和变种之间杂交可育,存在基因流,“中苜1 号”和“新疆大叶”都属于M. sativasubsp.sativa,仅仅解析一个亚种的基因组对于研究紫花苜蓿复合体是远远不够的。
除基因组之外,尚缺乏详细的紫花苜蓿转录组图谱。2021 年之前,发表了大量紫花苜蓿转录组的文章,极大地推动了紫花苜蓿的基础研究。由于当时缺乏基因组,所有转录组文章都采用无参组装,存在以下不足,如转录组组装质量较差、测序方法不一致、测序品种不统一、取样不够精细、不同转录组之间没有可比性及缺乏一个权威的紫花苜蓿组学数据库等。因此,紫花苜蓿的转录组仍有很多工作需要做。同时,紫花苜蓿的泛基因组、单倍型基因组、单细胞转录组、小RNA 组、蛋白质组、代谢组、DNA 甲基化和修饰组学等仍需研究。与紫花苜蓿相比,其他主要牧草的组学数据更少,亟须拓荒式研究。
2.3 遗传转化
利用重组DNA 原理,将目的基因整合到植物基因组中,使之稳定遗传并获得抗逆、高产、优质等靶标性状,称为遗传转化。任何一个物种,高效遗传转化体系的建立将为其打开转基因育种、基因编辑育种和反向遗传学的大门。目前,主要农作物都已建立了稳定、高效的农杆菌介导的遗传转化技术。紫花苜蓿、蒺藜苜蓿、百脉根(Lotus corniculatus)、白三叶、二穗短柄草(Brachypodium distachyon)、多年生黑麦草和柳枝稷(Panicum virgatum)等牧草的遗传转化系统也比较成熟。而其他牧草的遗传转化报道较少,或有零星报道但重复性差,这些牧草未来可在根癌农杆菌介导、发根农杆菌介导、基因枪转化、电穿孔法、花粉管通道法、茎尖或芽尖分生组织转化和纳米载体介导等不同转化技术上进行探索。
在高效遗传转化体系建立的基础上,建议进一步建立高效基因编辑体系,为基因编辑育种奠定基础。目前,国际上牧草的基因编辑报道不多,我国科学家已在紫花苜蓿[21]和蒺藜苜蓿[22-23]获得初步成功。
3 若干重点科学问题
3.1 乡土草资源
乡土草(图3)是本地野生的、或在本地生存很多年的,对当地土壤、气候等自然环境具有良好适应性的草类植物,是相对引进植物而言的,如我国东北的羊草、碱茅(Puccinellia chinampoensis)等,西北的无芒隐子草、醉马草(Achnatherum inebrians)、霸 王(Zygophyllum xanthoxylum)、沙 打 旺(Astragalus adsurgens)、红 豆 草(Onobrychis viciifolia)和柠条(Caragana korshinsk)等,青藏高原的垂穗披碱草(Elymus nutans)、老芒麦(E.sibiricus)、歪头菜(Vicia unijuga)、草地早熟禾(Poa pratensis)和扁杆早熟禾(P. pratensis)等[17]。在退化草地补播恢复中,乡土草对当地的环境条件更为适应,具有得天独厚的优势,比外来引进牧草更适合于草原修复和生态治理。
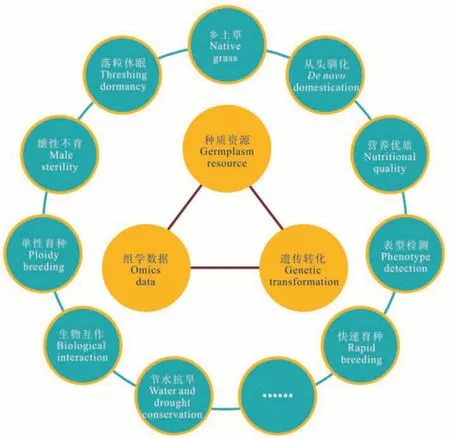
图3 我国牧草育种的关键瓶颈与主要科学问题Fig.3 Key bottlenecks and major scientific issues of forage breeding in China
此外,乡土草抗逆性强,蕴含着其他牧草没有的优异抗逆基因资源。但是由于乡土草种类多,研究人员少,导致以往对乡土草的关注不够,缺少积累。建议我国各个省区开展乡土草种质资源调查,摸清家底。在做好收集与保护工作的基础上,选择主要草种开展驯化选育,主要解决乡土草的种子高效采集与播种、种子休眠与破除、株型、地上生物量、栽培管理以及挖掘和利用乡土草特有的抗逆基因资源等。
3.2 落粒与休眠
任何优良牧草品种,如果种子产量低,就会限制其扩繁和推广。对野生植物而言落粒是一个有利性状,有助于种子的传播和种群的扩张;但对栽培作物而言,落粒是一个不利性状,因为种子成熟期不一致,大量种子在收获前从植株脱落掉落土壤,给种子收获造成严重损失,而且落地后的种子第2 年会成为其他后茬作物的杂草,危害后茬作物生长[24]。因此,任何牧草新品种,在选育过程中都必须尽可能地破除落粒这一劣性。落粒是一个数量性状,几乎所有的野生植物都存在落粒特性,禾本科牧草具有落粒特性,豆科野豌豆属、大豆属(Glycine)、菜豆属(Phaseolus)、苜蓿属、百脉根属(Lotus)、黄芪属(Astragalus)、锦鸡儿属(Caragana)、羽扇豆属(Lupinus)和扁豆属(Lablab)等牧草都具有裂荚特性。在对水稻的研究中发现,qSH1、Sh4和SHAT1等基因可影响小穗离层发育,是控制水稻落粒的主效基因,通过定点编辑qSH1基因可降低水稻的落粒性[25]。
牧草种子普遍存在休眠特性,休眠对种子安全度过非适宜生长期、保证种群延续具有重要作用,但也给人工草地建植、杂草防除、种子检验与加工等带来困难。禾本科牧草种子常具生理休眠,豆科牧草种子普遍具物理性休眠,偶见生理性休眠。从蒺藜苜蓿Tnt1 突变体库发现,KNOX4基因通过控制种皮栅栏状角质层的发育,影响种子的物理性休眠[26]。
建议首先大量收集种质资源或突变体、精确评价落粒/休眠特性,筛选易落粒/休眠、抗落粒/休眠种质,构建遗传群体,再通过GWAS、QTL 作图、转录组、解剖学、生理酶活等方法鉴定落粒/休眠基因,并与模式植物和农作物进行比较,揭示落粒/休眠调控机制、培育抗落粒/休眠高产牧草新品种。此外,应研发牧草种子大田栽培、水肥管理、授粉和收获清选等技术,提高优良牧草的种子产量。
3.3 雄性不育
牧草中含有多种特异的生殖方式,雄性不育不仅是牧草生殖发育研究的重要方向,更是牧草杂种优势利用的有效工具,具有非常重要的理论和应用价值。科学家已经在拟南芥、水稻和玉米中鉴定了上百个雄性不育基因。我国的水稻杂交育种经历了3 个世代,第1 代是细胞质雄性不育的三系法;第2 代是光温敏不育的两系法;第3 代也称智能不育杂交育种技术,是一种稳定的、能自我繁殖的、恢复性保持系一体的新型不育杂交育种体系。携带单合子的转基因水稻自交,按1∶1 产生转基因雄性不育和转基因可育两种种子,可根据红色荧光区分[27],玉米中也建立了类似体系[28]。为了满足第3 代杂交水稻育种的需要,已成功研发出繁殖系种子荧光检测分选仪。
EAN1基因在蒺藜苜蓿绒毡层中特异表达,属于bHLH 家族亚家族Ⅱ转录因子,EAN1 蛋白与bHLH 家族亚家族Ⅲ(a+c)1 成员形成异源二聚体调控绒毡层的降解,并发现该机制在被子植物中是保守的[29]。拟南芥和农作物雄性不育已经得到了详细的研究,这些研究成果为牧草雄性不育提供了重要的候选基因,为草类植物杂种优势利用和智能不育技术建立指明了研究方向。
3.4 倍性育种
倍性育种是培育优良牧草品种的重要手段。与同一物种的二倍体植株相比,多倍体植株具有高大、抗逆性强、茎叶繁茂、由一年生转变为越年生或多年生等特性,如二倍体和四倍体的紫花苜蓿、玉米。这些特性对农作物而言意义不大,但对以地上营养体为利用目标的牧草是非常有利的。加倍后,大部分植物的育性下降、种子产量降低,这些不利特性可以通过无性繁殖克服,禾本科牧草可以通过分蘖或根茎扩繁,豆科牧草可以通过扦插扩繁。倍性改变后的材料,可以为远缘杂交搭建新的桥梁。双单倍体的纯度高,可用于遗传图谱的构建,重要基因的QTL 定位。单倍体作为转基因受体可使外源基因迅速纯合,便于观察表型,加速选育进程。单倍体自然发生的概率极低,常采用孤雌生殖或孤雄生殖的办法人工诱导。
在水稻中,通过编辑OSD1、PAIR1和REC8三个基因,可以使水稻从二倍体变为同源四倍体、同源八倍体,编辑MTL基因可以使水稻从二倍体变为单倍体[30],其同源基因在拟南芥和玉米中获得了一致的结果。这些成果为牧草的倍性育种提供了新的研究思路。
3.5 牧草与生物互作
在草地上,牧草与生物间存在广泛的相互作用,研究领域包括禾草内生真菌互作、牧草与丛枝菌根真菌互作、豆科牧草与根瘤菌互作、牧草与病毒互作、牧草与害虫互作、牧草与杂草互作等。
大量研究表明,内生真菌与宿主植物形成共生体后,可以使宿主具有耐旱、耐寒、抗病虫等优良特性。近年来,接种禾草内生真菌,使其通过种子垂直传播,通过这种方法进行禾草育种已成为国际草业领域的发展趋势。将分离于野大麦(Hordeum brevisubulatum)的内生真菌(Epichloë bromicola)人工接种至非天然宿主栽培大麦(H. vulgare),可显著提高宿主的地上生物量和单株种子产量,并创制出了牧草新品系[31]。此外,牧草还蕴含着农作物不具备的优异抗病基因。20 世纪60 年代,李振声院士通过远缘杂交将长穗偃麦草[Elytrigia elongata(Host)Nevski]的抗“条锈病”基因转移给小麦,选育出持久性抗病小麦品种小偃6 号。2020 年,孔令让教授团队通过远缘杂交再次将长穗偃麦草的抗“赤霉病”主效基因Fhb7转移给小麦,揭示了抗病分子机制。菟丝子(Cuscutaspp.)是一种全寄生植物,严重威胁紫花苜蓿的生产,被我国海关总署列为紫花苜蓿检疫性有害生物。从栽培番茄(Solanum lycopersicum)中克隆Solyc08g016270基因并转入紫花苜蓿,转基因紫花苜蓿新材料通过抑制菟丝子吸器形成可以有效抵抗菟丝子寄生[32]。
今后,可利用生物技术和信息技术等手段,揭示牧草和生物的互作分子机理、挖掘主效基因,创制优异牧草新品种;将实验室研究和大田试验相结合,提出有害生物绿色高效防控策略。
3.6 节水耐旱
我国的粮食生产中,干旱造成的损失几乎是其他自然灾害造成损失的总和。天然草地方面,我国干旱半干旱地区约占国土面积的50%,北方草原地区干旱化正在进一步加剧,在过去的几十年中干旱半干旱草地的生产力持续下降,而在未来温度增高2~4 ℃和降水减少的情景下,草地生产力将会降低40%~90%[33]。人工草地方面,我国西北、东北和内蒙古商品苜蓿种植面积占全国的92.0%,上述地区绝大部分处于我国干旱和半干旱地区。因此,不论是天然草地还是人工草地,节水耐旱牧草新品种的培育意义重大。
植物的耐旱能力通过ABA、油菜素内酯、生长素等激素感知和调控,已经鉴定了大量基因参与植物对干旱的响应[34]。兰州大学科研人员将无芒隐子草和霸王的抗旱基因转入紫花苜蓿,创制出了紫花苜蓿转基因耐旱新材料[35-36]。在天然草地上,建议驯化选育当地优异的乡土草种,如无芒隐子草、霸王以及赖草属和苜蓿属野生牧草等。在人工草地上,建议加强紫花苜蓿耐旱性研究,可采用GWAS、QTLs、转录组、转基因、全基因组选择等研究方法。在培育耐旱品种时,一定要兼顾新材料的耐寒性(越冬)和耐热性(夏季高温)等。未来的研究中,可使用合成生物学合成一些小肽、激素类似物和受体激动剂来调控牧草的耐旱能力。
3.7 营养优质
人类仅食用粮食作物的籽粒,因此育种目标是种子产量、口感、香味、保健以及其他特殊需要等。但是,家畜的采食对象是牧草的全部地上营养体,因此育种目标是地上总生物量,以及整个地上部的粗纤维、粗蛋白、单宁、木质素等。紫花苜蓿的品质育种方面,应考虑粗纤维和粗蛋白含量,奶牛的过瘤胃蛋白、总可消化养分、泌乳净能、体外干物质消化率等。我国科学家已经在玉米中建立了大量遗传群体,通过GWAS、突变体库等方法鉴定了大量控制叶夹角、光周期、籽粒产量等驯化或育种关键功能基因。青贮玉米作为奶牛等反刍动物的主要粗饲料,建议利用已有材料和数据,进一步鉴定玉米地上总生物量(不单单是籽粒)以及粗纤维、总可消化养分、泌乳净能、氨基酸组成(如调高含硫氨基酸)等,加速青贮玉米的分子设计育种。另外,70%左右的玉米籽粒被加工为饲料,建议根据奶牛、猪、蛋鸡、肉鸡的个性化营养需要进行新品种选育。
此外,在亿万年的进化过程中,草类植物为了防止动物过度采食,体内积累了一些有毒成分。如禾本科的醉马草具有高产、抗逆、多年生的优良特性,但体内含有麦角酰胺等有毒次生代谢物,牛、羊采食后会中毒。豆科牧草白花草木樨(Melilotus albus)和黄花草木樨(M. officinalis)生物量大、耐旱能力突出,但含有香豆素。因此,研究致毒物质的代谢途径、致病机理,筛选无毒种质也是牧草品质育种的重要方向。
3.8 从头驯化
玉米、小麦、水稻和大豆等主要农作物都经过了人类5000~10000 年的驯化,与驯化前相比,驯化后的作物在形态、产量等方面发生了翻天覆地的变化。目前,科学家已鉴定了一批主要驯化基因。李家洋院士团队通过四步法实现了异源四倍体野生稻(O. alta)的“快速”从头驯化(de novodomestication)。第一,收集并筛选野生稻底盘种质资源;第二,建立从头驯化技术平台,即高质量参考基因组和高效基因组编辑体系;第三,多基因选择、编辑、聚合以及田间评估;第四,推广应用[37]。
几乎所有草类植物的驯化时间短或未经驯化,建议建立相关技术平台编辑牧草同源基因,即在农作物中鉴定的驯化基因和重要农艺性状基因的同源基因,实现草类植物“一步式”从头驯化。
3.9 表型检测
植物表型具有很强的可塑性,是自身基因表达和环境影响互作的结果,表型检测与植物育种息息相关。在作物育种进入分子时代的今天,人工方法测定作物表型,具有耗时、主观性强、不准确、效率低等缺陷。作物育种人员需要高精度、高通量、低成本的技术观察作物的生长发育、产量以及作物对生物、非生物胁迫的抗性等表型数据。作物表型自动化检测技术是新兴的研究热点,对作物重要农艺性状调控机制研究、品种选育具有重要意义。目前,以小麦、玉米、马铃薯(Solanum tuberosum)等为研究对象,已开发了多种成像技术,如可见光成像、多光谱遥感、热红外成像、荧光成像、3D 成像等,可以进行植物株型、杂草识别、病虫害和产量预测等检测[38]。
牧草与农作物不同,对表型自动化检测技术要求更高,如多数牧草具有多年生、匍匐、丛生、攀援、植株高大、叶片细长/复叶、花器官和种子小、同一物种表型变异大等特性。除了成像技术外,牧草表型自动化检测系统还应具有以下功能:土壤水分动态检测、自动灌溉/称重、花盆自动传输、自动控温控光、叶片温度感知、牧草营养价值活体无损伤检测、地上/地下生物学估算等。
3.10 快速育种
缩短牧草世代间隔,加速后代纯合速度可大幅提高育种效率。快速育种(speed breeding)不以种子产量为目的,通过延长光照时间提前开花、收获生理成熟种子人工干燥(减少种子后熟时间)等方法,缩短牧草生育期。南繁加代一年只能繁育两代,但人工控温、控光可进一步提高育种效率。通过多年探索,科研人员已经建立了多种长日照作物的快速育种技术,光照长度采用22 h,通过提前收获和增加种植密度,完成春小麦、大麦、鹰嘴豆(Cicer arietinum)及豌豆(Pisum sativum)1 年繁殖6 代[39]。对于短日照作物,适当缩短光照长度也可缩短生育期。快速育种可以克服牧草多年生、生育期长、多倍体的不足,在牧草杂交后代纯化、转基因植株纯化和基因编辑植株外源基因分离等育种过程中具有重要的科学价值。可加速牧草杂交后代纯化、转基因植株纯化、基因编辑植株外源基因分离以及作图群体、重组自交系和近等基因系等构建。
除上述10 个科学问题外,下列科学问题同样重要,由于篇幅所限,不在此一一赘述。如耐盐碱、耐低温、耐涝、抗紫外、耐瘠薄、耐放牧践踏、耐生存竞争等的机理与新品种选育,花期调控、自交不亲和的分子机理与应用,重要牧草的起源、系统进化与物种形成机制,全基因组选择评估育种群体重要性状的遗传潜力,牧草新品种的认证与保护等。
4 结语
牧草育种关系国家的食物安全和生态安全,决定生态、生产的协调发展。客观上,牧草育种面临众多难题,如物种多、多年生、多倍体、异花授粉、基因组复杂、遗传转化难等。主观上,与农作物相比,国家投入少、成果产出低、研究平台差,作为牧草之王的紫花苜蓿从未被纳入我国农业生产主体,得到应有的重视。例如,近20 年与牧草育种有关973 项目仅有2 个,2020 年底中国科学院启动了战略性先导科技专项(A 类)“创建生态草牧业科技体系”,其中“牧草新品种定向分子选育”是该专项包含的6 个项目之一。与水稻、小麦、玉米等农作物,或者与林木和蔬菜相比,国家在牧草育种上支持力度亟须提高。上述客观和主观上的原因,导致牧草育种难以支撑国家的食物安全和生态安全,成为我国农业发展的“卡脖子”问题。
笔者认为,我国牧草育种的3 个关键瓶颈是:种质资源、组学数据和遗传转化(图3)。依据面向育种科技前沿和面向国家重大需求,提出了10 个主要科学问题:乡土草资源挖掘与利用、落粒与休眠、雄性不育、倍性育种、生物互作、节水耐旱、从头驯化、营养优质、表型自动化检测、快速育种以及其他科学问题等。我国牧草种类繁多,可根据牧草种植面积、国家需求等因素选择少数牧草集中研究,同时根据先当前后长远、先易后难的原则选择合适的科学问题。力争到2030 年,培育出一批有自主知识产权的、市场竞争力强的牧草新品种,尤其是在紫花苜蓿、羊草、燕麦、青贮玉米等主要牧草上培育出“突破性”的新品种,保障草食畜牧业生产和生态治理用种,期望在短时间内缩短与美国等国家牧草育种水平上的差距,并引领全球牧草育种水平。
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13 06:38:52
今日农业(2021年10期)2021-07-28 06:28:00
现代畜牧科技(2021年4期)2021-07-21 06:13:04
儿童时代·幸福宝宝(2019年9期)2019-10-28 18:04:52
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
幼儿园(2018年15期)2018-10-15 19:40:36
莫愁·家教与成才(2017年7期)2017-07-11 21:31:47
中国三峡(2017年4期)2017-06-06 10:44:22
为了孩子(3~7岁)(2016年6期)2016-05-14 03:09:17
鹿鸣(2015年7期)2015-05-30 10:48:04
