
纤维素酶和木聚糖酶对象草青贮发酵品质及体外消化率的影响
2021-12-09 09:05:52吴永杰代童童尹雪敬贾玉山李君风
草地学报 2021年11期
丁 浩, 吴永杰, 邵 涛, 赵 杰, 代童童, 董 东, 尹雪敬, 贾玉山,李君风*
(1.南京农业大学草业学院饲草调制加工与贮藏研究所, 江苏 南京 210095; 2.内蒙古农业大学农业农村部饲草栽培加工与 高效利用重点实验室, 内蒙古 呼和浩特 010018; 3.内蒙古农业大学教育部草地资源重点实验室, 内蒙古 呼和浩特 010018)
象草(PennisetumpurpureumSchumach)属于禾本科狼尾草属多年生草本植物,具有生物产量高、再生性强、生长速度快等优点,广泛种植于热带、亚热带地区,是反刍动物重要的粗饲料来源[1]。热带、亚热带地区雨热同期、高温高湿,牧草很难调制成干草进行全年储藏利用。而青贮不受气候条件的限制,青贮饲料可全年长期有效地保存,调节饲草季节性供给不平衡[2-3]。
象草水溶性碳水化合物含量相对较低、粗纤维含量高,表现出多茎、多空、节间粗硬的物理特性。在青贮调制过程中难以压实,残余的空气较多,造成植物呼吸作用及好氧性微生物活动时间延长[4]。好氧腐败菌代谢易使水溶性碳水化合物大量损失,乳酸菌发酵底物不足,乳酸生成量少,pH很难快速降到4.2以下,往往导致青贮发酵品质不佳[5]。
酶制剂常用于改善青贮发酵品质[6]。添加纤维素酶能使纤维水解为葡萄糖,增加乳酸菌发酵底物;木聚糖酶能将木聚糖类半纤维素降解为低聚糖和木糖[7]。同时纤维素酶与木聚糖酶能打破青贮原料细胞壁表面结构,使纤维素网状结构变得松散,比表面积增大。因此反刍动物采食酶制剂处理后的青贮饲料,更利于瘤胃微生物的附着与分解,提高饲料的消化率[8]。Desta等人[9]在象草青贮中添加木聚糖酶,发酵30 d后,青贮过程中水溶性碳水化合物和乳酸含量增加,半纤维素含量降低。李茂[10]报道王草青贮时添加纤维素酶,显著提高了乳酸含量、降低了pH值、酸性洗涤纤维和中性洗涤纤维含量,提高了发酵品质。酶制剂添加剂在暖季型牧草青贮中研究较多,但在生产运用时仍面临作用不稳定、普适性低等问题。本试验以象草为材料,旨在探讨纤维素酶、木聚糖酶单独以及组合添加对象草青贮发酵品质和体外消化率的影响,为优质象草青贮饲料的生产和利用提供理论依据与技术支持。
1 材料与方法
1.1 试验材料
供试象草种植于南京农业大学试验地(32°01′19″ N,118°51′08″ E,海拔25 m),于2020年8月27日刈割,此时象草处于营养生长期,其化学成分如表1所示。试验所用酶为青贮专用酶制剂,购于中国南京奥德美生物技术有限公司。纤维素酶、木聚糖酶的酶活为5 400 U·g-1,添加量参考使用说明。实验室青贮窖采用容积为2 L有内外盖的聚乙烯容器。
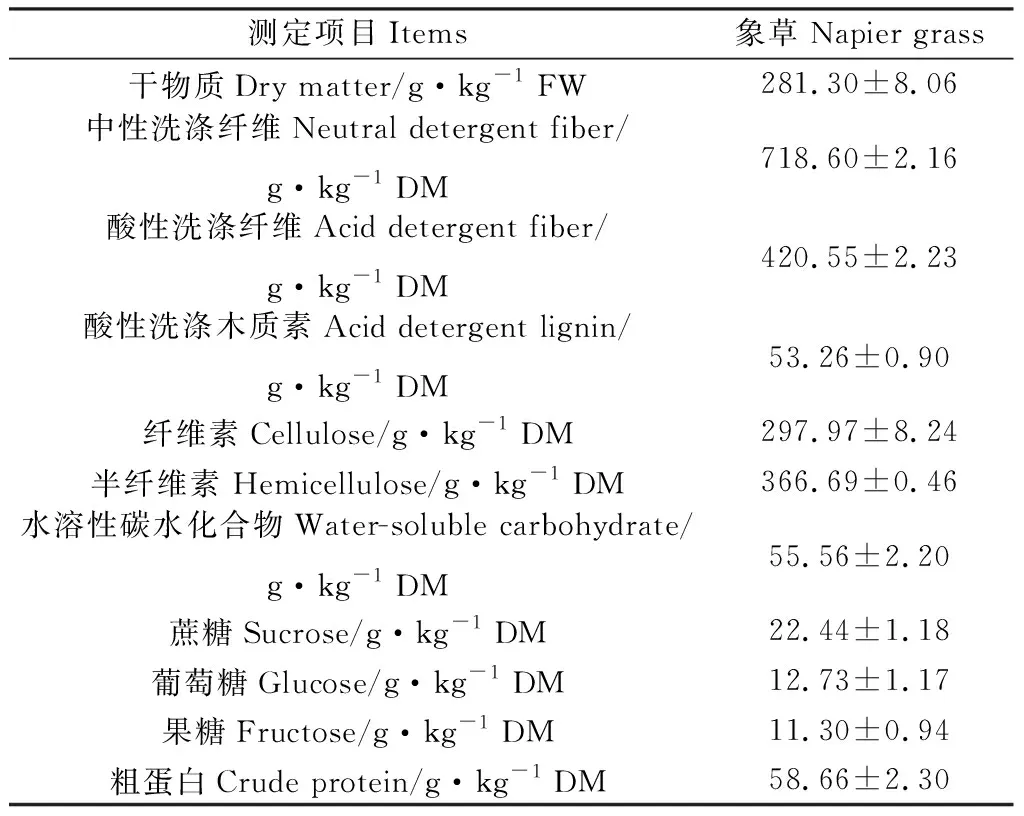
表1 象草的化学成分Table 1 The chemical compositions of napier grass
1.2 试验设计
试验采用完全随机区组设计,设对照组(CK),添加纤维素酶组(CE),添加木聚糖酶组(XE),组合添加酶组(CX,纤维素酶+木聚糖酶)。添加量均以青贮原料鲜重(Fresh weight,FW)为基础,其中纤维素酶、木聚糖酶添加量为0.3%(酶活为16.8 U·g-1FW),组合添加酶组添加量为0.15%纤维素酶+0.15%木聚糖酶。分别在青贮7,30,60,90 d后打开实验室青贮窖取样,测定相关发酵指标。每个处理每个青贮天数设3个重复,总共48个实验室青贮窖。
1.3 试验方法
1.3.1青贮饲料调制 象草切碎至2~3 cm,茎叶充分混合均匀后,按试验设计添加量将溶解于灭菌水中的纤维素酶、木聚糖酶溶液喷洒在切碎的象草上,对照组加等量灭菌水,再次充分混匀,装入2 L实验室青贮窖中,每个青贮窖中装1.36 kg,压实后盖上内外盖,用胶带密封,置于室温(25℃±5℃)下保存。
1.3.2样品处理 青贮窖打开后,将全部青贮饲料取出混匀,采用四分法精准称量样品20 g放入100 mL广口锥形瓶中,加入60 g蒸馏水,密封后于4℃冰箱静置24 h,然后通过4层纱布和定性滤纸过滤,得到青贮饲料的浸提液,立即测定pH后,于—20℃冰箱冷冻保存待测。浸提液用于测定氨态氮、乳酸、挥发性脂肪酸含量。将剩余青贮饲料收集,于65℃烘箱中烘60 h以上至恒重,粉碎过0.5 mm筛保存于自封袋中,用于水溶性碳水化合物、中性洗涤纤维、酸性洗涤纤维和体外消化率的测定。
1.3.3测定指标及分析方法 采用烘干恒重法测定青贮饲料干物质含量[11];pH值使用211型pH计(意大利哈纳科技有限公司)测定[12];氨态氮含量采用苯酚-次氯酸钠比色法测定[13];缓冲能采用盐酸、氢氧化钠滴定法测定[14];水溶性碳水化合物采用硫酸-蒽酮比色法测定[15];采用范氏纤维分析法测定青贮饲料中中性洗涤纤维(Neutral detergent fiber,NDF)、酸性洗涤纤维(Acid detergent fiber,ADF)和酸性洗涤木质素(Acid detergent lignin,ADL)含量,NDF测定时加入α淀粉酶、亚硫酸钠[16]。
采用高效液相色谱仪(1260 HPLC;德国安捷伦有限公司)测定乳酸及挥发性脂肪酸含量,色谱柱为Carbomix®H-NP5,流动相为2.5 mmol·L-1H2SO4,流速设定0.5 mL·min-1,温度设定55℃[14]。
采用高效液相色谱仪(1260 HPLC;德国安捷伦有限公司)测定青贮饲料中葡萄糖、果糖、蔗糖含量,色谱柱为Agilent InfinityLab Poroshell 120 HILIC-Z,流动相0.3%的乙腈氨水溶液,流速设定0.4 mL·min-1,温度设定35℃[12]。
1.3.4体外消化率测定方法 瘤胃液取自4只体况相近,装有永久性瘘管的去势波尔杂交山羊(平均体重为35 kg)。饲喂水平及日粮组成为:苜蓿干草45 g,羊草干草350 g,混合精料240 g(10%麸皮+25%豆粕+65%玉米粉),维生素和矿物质。于晨饲前采集瘤胃液,四层纱布过滤,置于39℃保温瓶中,并立即带回试验室,通入CO2维持厌氧条件。
称取1 g青贮饲料样品(65℃烘干后粉碎,过0.5 mm筛)于滤袋中,每个处理设3个重复,密封后置于血清瓶中,再向血清瓶中加入20 mL瘤胃液和40 mL McDougal缓冲液,置于摇床中培养(39℃,150 r·min-1),进行空白校正后,使用压力传感器技术记录4,8,12,24,48,72 h的产气量。试验结束后,从血清瓶中取出滤袋,洗涤后在65℃烘箱中干燥48 h至恒重,测定DM,NDF和ADF残留量,计算体外干物质消化率(InvitroDM digestibility,IVDMD),体外中性洗涤纤维消化率(InvitroNDF digestibility,IVNDFD)和体外酸性洗涤纤维消化率(InvitroADF digestibility,IVNDFD)[17]。利用GraphPad 6.01软件,根据France产气模型计算产气动力学参数,模型:GPt=B×(1—exp(-c×(t—Lag)))[18],其中GPt是在t时间的累计产气量,B表示理论最大产气量(mL);c表示产气速率常数(mL·h-1),Lag表示体外发酵产气延滞时间(h),t表示产气时间点(h)。
动力有限元模型的离散参数需要根据相应地层的剪切波波速及激励的频率范围来确定[16]。为了简化模型,同时又能反映地层的动力特性,根据地层剪切波速的变化范围,将复杂的真实地层简化为5层水平构造,每层代表一个主要的剪切波速域[17]。剪切波速cs的计算公式[18]为:
1.4 数据处理与分析
采用Excel 2019进行基础数据收集归纳。采用统计分析系统(SAS 8.0)GLM程序对数据进行双因素方差分析(ANVOA),采用Tukey’s HSD方法对处理间和青贮天数间平均数进行多重比较(P<0.05)。
2 结果与分析
2.1 添加酶制剂对象草青贮过程中pH、干物质损失、干物质含量和氨态氮的影响
如表2所示,酶制剂处理、青贮天数及其互作对青贮过程中pH、干物质损失、干物质含量和氨态氮有显著影响(P<0.05)。整个青贮过程中,各组pH逐渐降低,30 d降至最低,随后略有提高。CE,XE,CX组pH低于CK组;青贮30 d后CX组pH显著低于其它三组(P<0.05),CE与XE组无显著差异。
青贮过程中,干物质含量逐渐降低,青贮前7 d各组差异不显著,之后CE,XE,CX组干物质含量显著高于CK组(P<0.05);CE,XE组青贮60 d,90 d后干物质含量显著高于CX组(P<0.05),CE和XE组无显著差异。氨态氮含量随着青贮的进行显著提高(P<0.05),CE,XE,CX低于CK组;青贮30 d后CX组氨态氮含量显著低于CE和XE组(P<0.05),CE和XE组无显著差异。
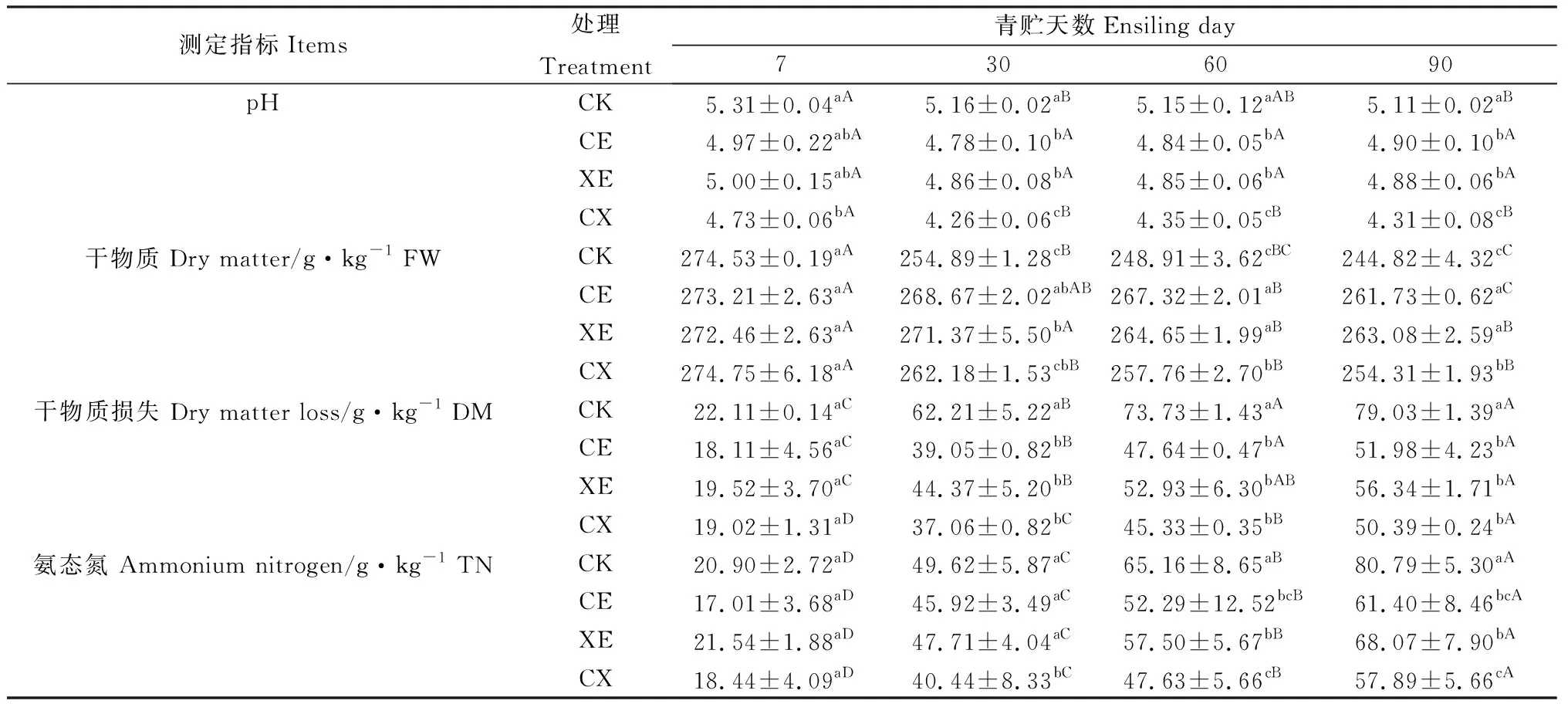
表2 添加酶制剂对象草青贮过程中pH、干物质损失、干物质和氨态氮含量的影响Table 2 Effects of enzymes addition on pH,DM loss,AN and DM content of napier grass during ensiling
2.2 添加酶制剂对象草青贮过程中有机酸含量的影响
如表3所示:酶制剂处理、青贮天数及交互作用对青贮过程中乳酸、乙酸、丙酸和丁酸含量有显著影响(P<0.05)。青贮过程中乳酸含量逐渐提高,青贮第30 d达到峰值,之后逐渐降低。CE,XE,CX组乳酸含量在21.28~48.44 g·kg-1DM之间,显著高于对照组(P<0.05);CX组乳酸含量始终高于CE,XE组。CE与XE组间青贮前期乳酸含量无显著差异,但青贮60 d,90 d后CE组乳酸含量显著高于XE组(P<0.05)。
随着青贮天数的延长,乙酸含量逐渐提高。CE,XE,CX组乙酸含量始终低于CK组。青贮过程中乳酸/乙酸下降,但CE,XE,CX组均高于CK组(P<0.05);除青贮第7 d以外,CX组乳酸/乙酸均显著高于CE,XE组(P<0.05)。青贮过程中各组丙酸含量逐渐提高,除对照组青贮30 d后检测到少量丙酸(3.49~4.85 g·kg-1DM)外,其余处理组均仅检测出微量丙酸(0.38~1.86 g·kg-1DM)。
整个青贮过程中,各组丁酸含量有所提高,但含量均较低。青贮30 d后CE,XE,CX组检测到少量的丁酸(2.95~6.17 g·kg-1DM),显著低于CK组(7.11~13.02 g·kg-1DM,P<0.05)。
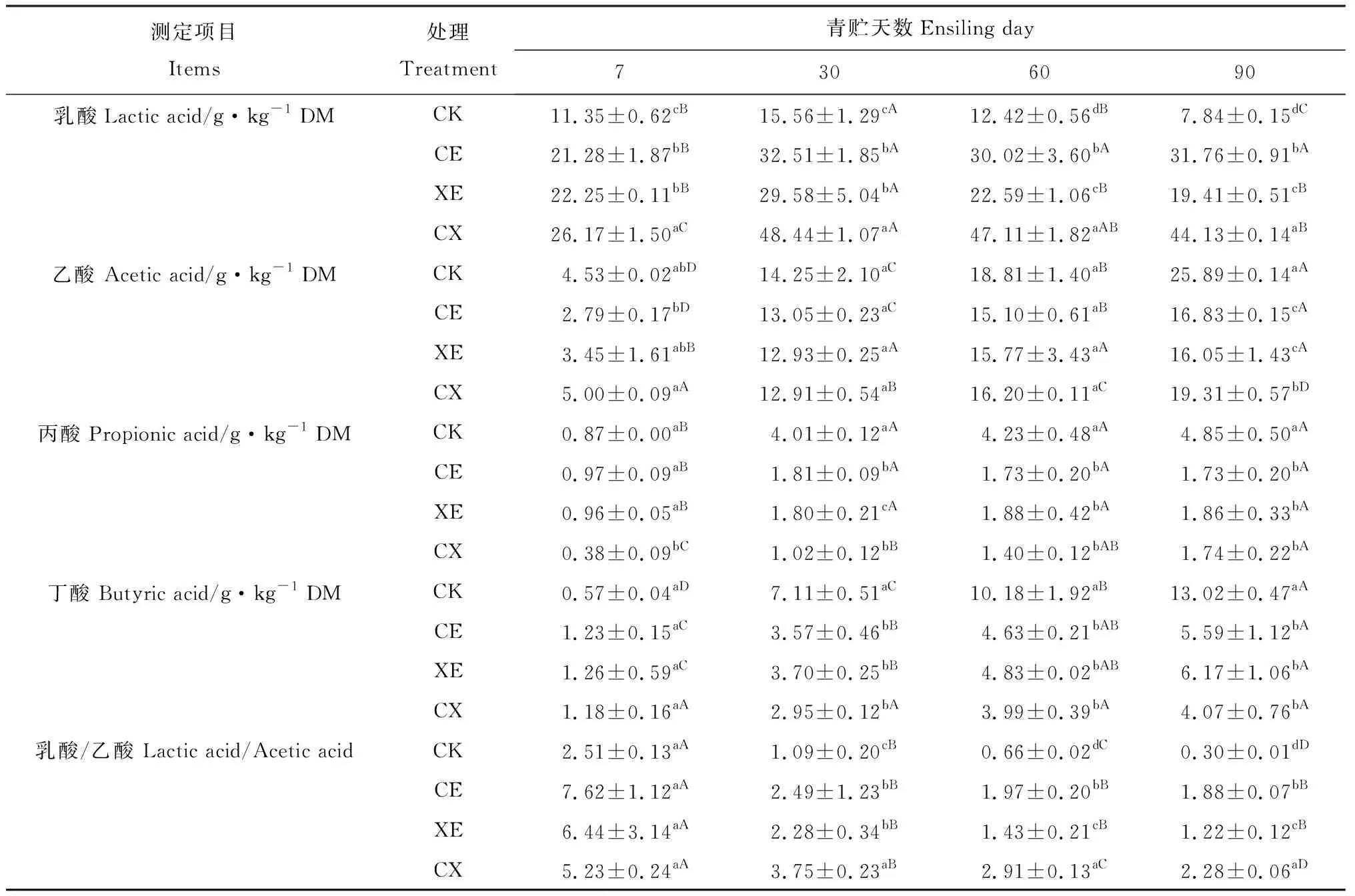
表3 添加酶制剂对象草青贮过程中有机酸含量的影响Table 3 Effects of enzymes addition on organic acid content of napier grass during ensiling
2.3 添加酶制剂对象草青贮过程中结构性碳水化合物组分含量的影响
如表4所示,酶制剂处理、青贮天数及交互作用显著影响青贮过程中除ADL以外所有结构性碳水化合物组分含量(P<0.05)。青贮过程中各组NDF含量逐渐降低,CE,XE,CX显著低于CK组(P<0.05);CX组NDF含量最低,在青贮第30 d与90 d显著低于CE,XE组(P<0.05);CE与XE组除青贮第30 d外无显著差异。
青贮过程中添加酶处理组ADF含量逐渐降低,且均低于CK组。从青贮第30 d起,CE与CX组ADF含量显著低于XE组(P<0.05),CE与CX组无显著差异。整个青贮过程中,各组ADL含量呈略有提高的趋势,CE,XE,CX组ADL含量始终高于CK组,但各酶添加组差异均不显著。
随着青贮天数的延长,纤维素、半纤维素含量逐渐降低。CE,XE,CX组纤维素含量低于CK组,青贮30 d后CE,CX组显著低于XE组(P<0.05),CE,CX组无显著差异。CE,XE,CX组半纤维素含量低于CK组,CX组半纤维素含量最低,在青贮30 d与60 d时显著低于CE,XE组(P<0.05),XE组半纤维素含量低于CE组。
2.4 添加酶制剂对象草青贮过程中水溶性碳水化合物、葡萄糖、果糖和蔗糖含量的影响
如图1所示,整个青贮过程中,水溶性碳水化合物含量持续降低,CE,XE,CX水溶性碳水化合物含量高于CK组,其中CX组最高,CE与XE无显著差异。青贮过程中,各组葡萄糖含量逐渐降低,CE,XE,CX组葡萄糖含量高于CK组,CX组含量最高,且在青贮第7 d,30 d显著高于CE,XE组(P<0.05)。随着青贮时间的延长,蔗糖、果糖含量逐渐降低,添加酶处理组始终显著高于CK组(P<0.05),CX组蔗糖、果糖含量最高,CE与XE组无显著差异。
2.5 添加酶制剂对象草青贮饲料体外消化率、体外产气量的影响
如表5所示,整个体外发酵过程中,CE,XE,CX组48,72 h累计产气量均高于CK组,CX组48,72 h累计产气量显著高于CE,XE组(P<0.05),CE,XE组无显著差异。添加酶制剂处理后的象草青贮饲料IVDMD在57.09%~61.30%之间,显著高于对照组(48.03%,P<0.05),CX组IVDMD显著高于CE,XE组(P<0.05),CE和XE组无显著差异。添加酶制剂象草青贮IVNDFD提高了4.33%~8.98%,IVADFD提高了7.83%~12.46%,均显著高于对照组(P<0.05)。
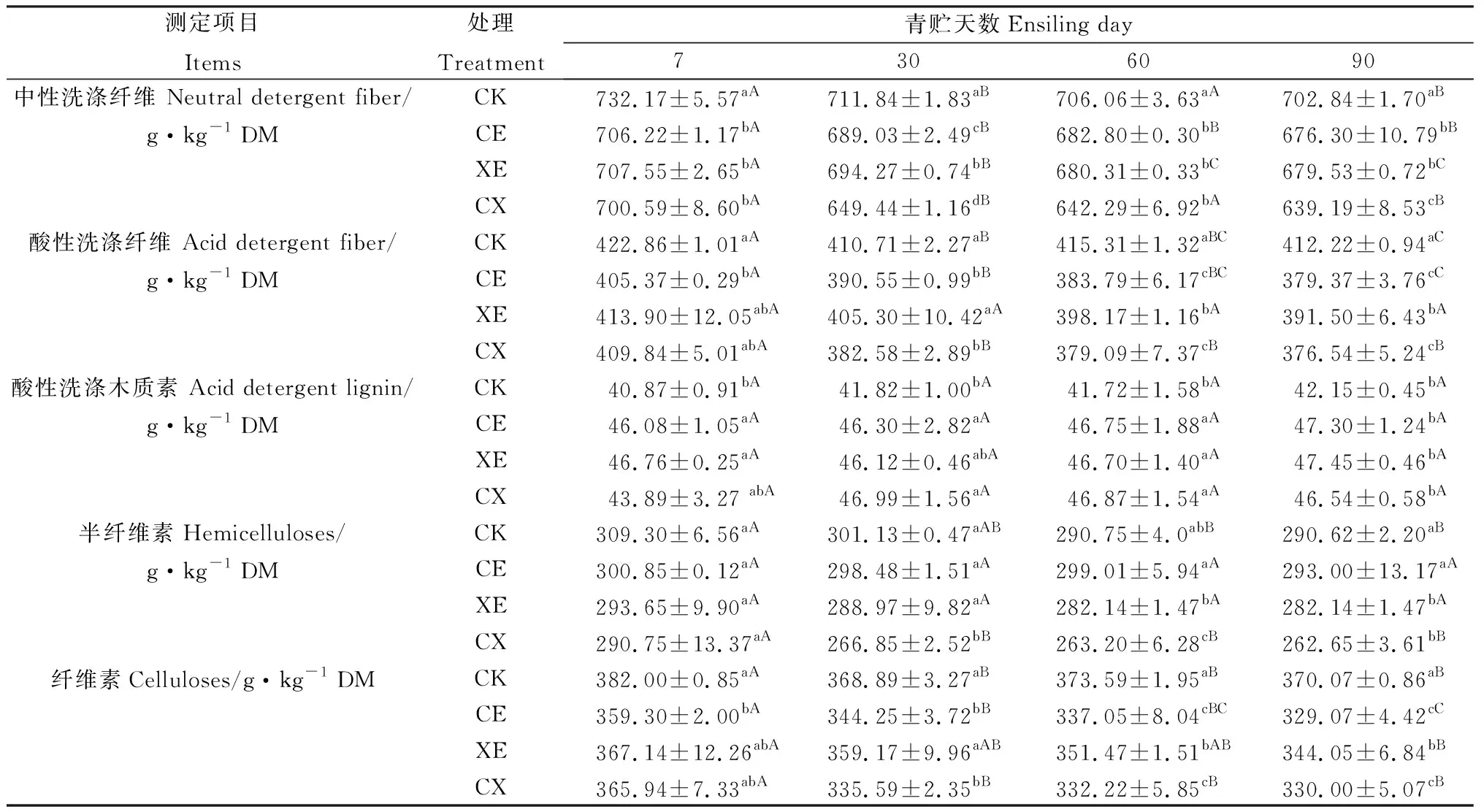
表4 添加酶制剂对象草青贮过程中结构性碳水化合物组分含量的影响Table 4 Effects of enzymes addition on structural carbohydrate components of napier grass during ensiling
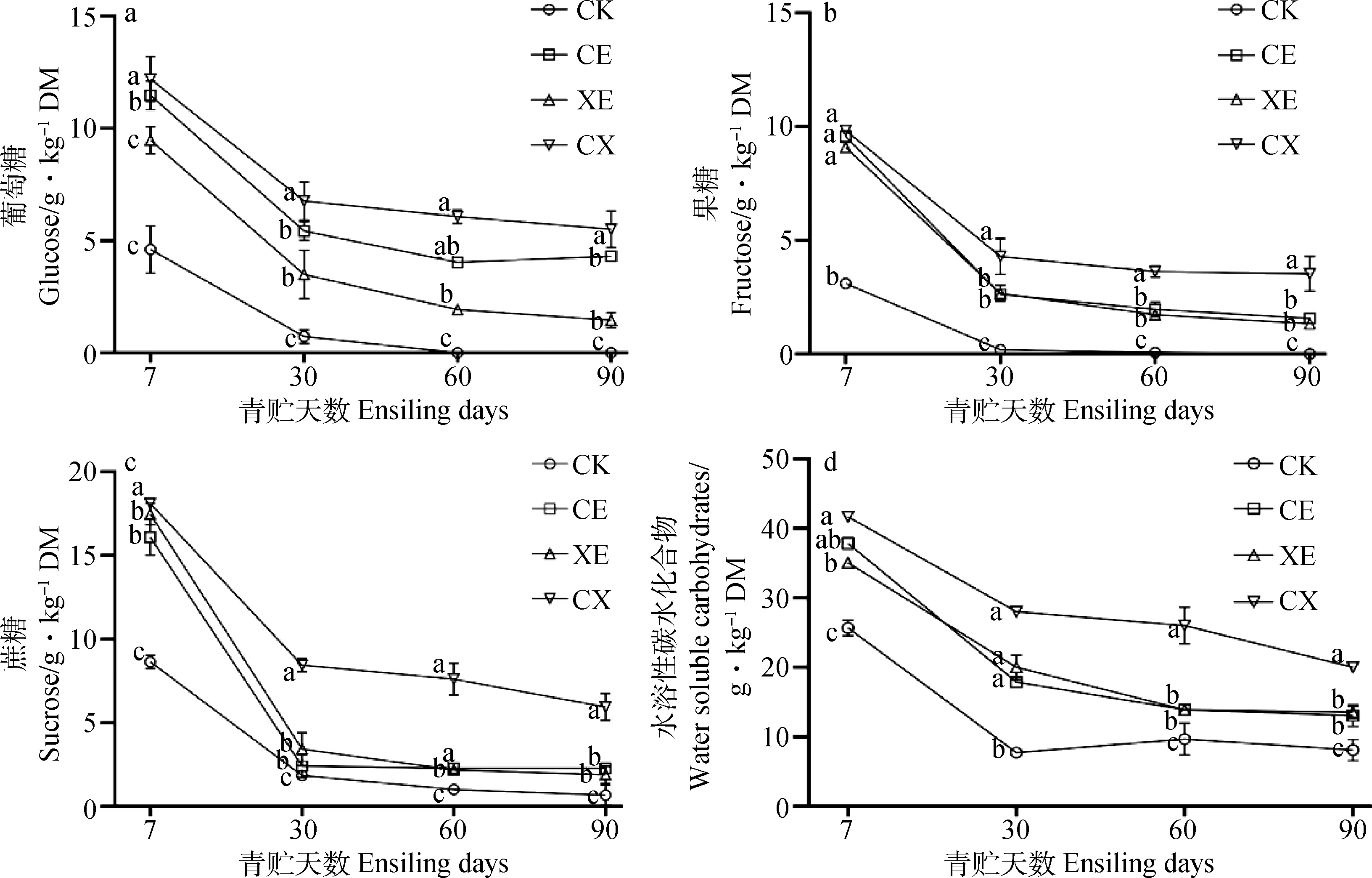
图1 酶制剂对象草青贮过程中水溶性碳水化合物、葡萄糖、果糖、蔗糖含量的影响Fig.1 Effects of enzymes addition on water soluble carbohydrate,glucose,fructose,sucrose contents of napier grass during ensiling注:不同小写字母表示同一时间内不同处理差异显著(P<0.05)Note:Different lowcase letter indiacte significant differences at the 0.05 level
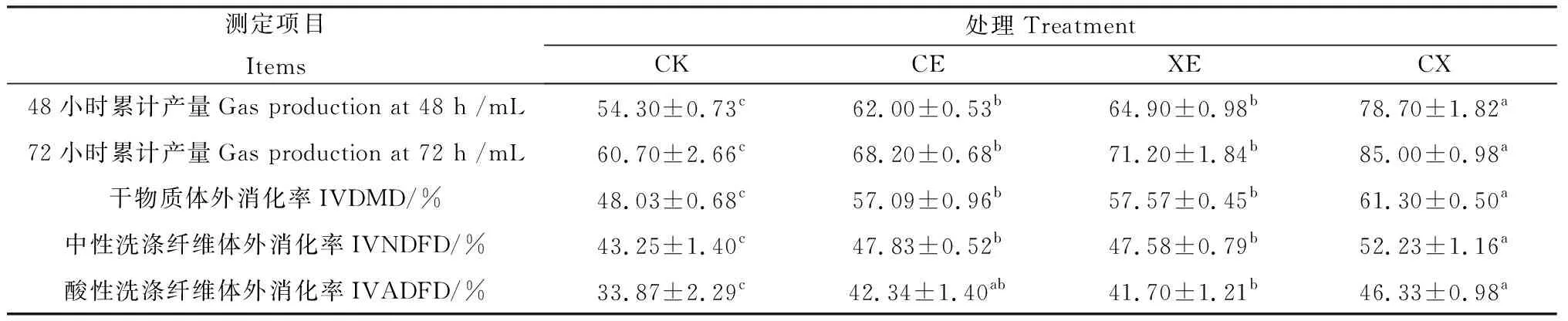
表5 酶制剂对青贮90 d象草青贮饲料体外消化率和体外产气量的影响Table 5 In vitro digestibility coefficient and gas production of napier grass silage treated with enzymes after 90 days of ensiling
3 讨论
3.1 添加酶制剂对象草青贮过程中发酵品质的影响
青贮过程中CE,XE及CX组干物质损失在18.11~56.34 g·kg-1DM之间,低于对照组22.11~79.03 g·kg-1DM,表明添加酶制剂组减少了干物质的损失。干物质损失主要是青贮早期的植物呼吸作用及青贮中、后期的微生物代谢所致[19]。青贮7 d后,酶制剂组乳酸含量显著高于对照组,表明在象草青贮中添加酶制剂促进了粗纤维的降解,释放水溶性碳水化合物,为乳酸菌提供了额外的发酵底物,快速产生乳酸,降低pH,抑制了不良微生物的活性,这与酶制剂组WSC含量显著高于对照组相呼应[20]。CE,XE及CX组在青贮30 d后pH值显著低于对照,CX组pH降至4.21,乳酸含量达48.84 g·kg-1DM,高于CE,XE组,说明在青贮中组合添加酶发酵品质优于单一添加。本研究与Smerjai等[21]研究结果一致。
通常青贮饲料中乳酸/乙酸大于2时,以同型乳酸发酵为主[22]。本试验中青贮前期乳酸含量提高,pH下降,以同型乳酸发酵为主,随着青贮的进行耐酸的异型发酵乳酸菌逐渐替代同型发酵乳酸菌成为优势菌群,既能产生乙酸,也能利用乳酸异化产生乙酸,导致乙酸含量提高,乳酸/乙酸下降[23-25]。
Tao等人[26]报道优质青贮饲料氨态氮含量应小于100 g·kg-1TN,本试验氨态氮含量最高为80 g·kg-1TN(< 100 g·kg-1TN),整个青贮过程中酶制剂组氨态氮含量低于对照组,青贮60 d显著低于对照组,这是由于酶添加组乳酸含量高、pH低,抑制了不良微生物的活性,减少了对蛋白质的降解,使氨态氮含量降低。Li等[27]在狼尾草(Pennisetumsinese)青贮中添加纤维素酶检测出微量的丙酸、丁酸,与本试验结果一致,表明在象草青贮中添加酶制剂能有效抑制有害微生物活性,减少了丁酸和丙酸的产生[3,14]。
3.2 添加酶制剂对象草青贮过程中碳水化合物的影响
青贮过程中添加酶制剂,有助于植物细胞壁成分的降解,使纤维素、半纤维素含量降低,水溶性碳水化合物含量提高[18]。本试验添加酶制剂、青贮天数及两者的交互作用显著降低了象草青贮过程中除ADL以外所有结构性碳水化合物组分的含量。青贮过程中CE,XE及CX组的纤维素、半纤维素含量均低于对照组,葡萄糖、果糖、蔗糖及水溶性碳水化合物含量高于对照组,认为酶解作用直接引起了纤维素、半纤维素的降解,导致水溶性碳水化合物含量的提高[9]。寇江涛等[28]报道在皇竹草中添加木聚糖酶,显著降低了ADF含量。不同种类的酶制剂之间具有协同降解粗纤维的作用,组合添加更有利于青贮早期将结构性碳水化合物降解为水溶性碳水化合物,本试验组合添加酶制剂组中纤维素酶与木聚糖酶协同作用导致纤维素、半纤维素含量显著低于单独添加纤维素酶、木聚糖酶[12],这与组合添加酶组具有较高的WSC含量相呼应。
Shao等[29]研究表明葡萄糖和果糖是乳酸菌优先利用的底物,青贮开始后乳酸菌增殖,利用葡萄糖和果糖,产生乳酸,乳酸含量逐渐提高,葡萄糖和果糖含量逐渐降低,青贮后期趋于稳定。本试验整个青贮过程中,CE,XE及CX组水溶性碳水化合物、葡萄糖、果糖和蔗糖含量均高于对照组,表明纤维素酶和木聚糖酶降解了部分纤维素和半纤维素,释放了葡萄糖、果糖和蔗糖,提高了水溶性碳水化物含量;其次添加纤维素酶和木聚糖酶加速了青贮过程中乳酸的生成,抑制了有害微生物对水溶性碳水化合物的消耗,使更多的WSC得以保留。黄秋连[30]和赵金鹏等[6]在羊草和水稻秸秆青贮中也得到相似结果。CX组WSC含量始终高于CE,XE组,纤维素和半纤维素含量显著低于CE,XE组,说明纤维素酶和木聚糖酶组合添加更有利于提高象草青贮发酵品质。
3.3 酶制剂对象草青贮饲料体外消化率的影响
体外模拟瘤胃发酵在一定程度上可以反映反刍动物对青贮饲料的消化率,IVDMD,IVNDFD,IVADFD和体外产气量是重要的衡量指标[31]。研究表明青贮饲料IVDMD与粗纤维含量负相关[20]。与对照组相比,酶制剂组青贮90 d后的青贮饲料IVDMD显著高于对照组,这与Chen等[32]研究结果一致。Li等[27]报道酶降解作用可以改变青贮饲料表面结构,增加纤维间的孔隙度、表面积和可及性,增加反刍动物瘤胃微生物酶与青贮饲料的接触面积,提高IVNDFD和IVADFD。本试验添加酶制剂组与对照相比IVNDFD提高了4.33%~8.98%,IVADFD提高了7.83%~12.46%,说明添加酶制剂组较低的ADF,NDF有利于瘤胃微生物对粗纤维的降解。CX组IVNDFD,IVADFD高于CE,XE组,这是由于纤维素酶与木聚糖酶之间的协同作用,不仅提高了青贮饲料的WSC、乳酸含量为瘤胃微生物提供了发酵底物,也裂解了象草中纤维素与半纤维素之间的连接键,使纤维素结晶度降低,比表面积增大,利于瘤胃微生物附着及降解[9]。
青贮饲料的发酵产物、碳水化合物和含氮物质是体外产气的主要底物[33-34]。体外产气速率越快,累计产气量越多,饲料消化率越高。本试验添加酶制剂组72 h累计产气量、产气速率常数和理论最大产气量均显著高于对照组,表明添加酶制剂可提高青贮饲料的消化率。IVNDFD,IVADFD的提高增加了饲料比表面积,利于瘤胃微生物的附着与增殖[35]。CX组72 h累计产气量显著高于CE,XE组,也证明了组合添加酶组青贮饲料消化率更高。
4 结论
综上所述,添加纤维素酶、木聚糖酶降低了纤维素、半纤维素含量,提高了葡萄糖、果糖、蔗糖及总水溶性碳水化合物含量,促进了青贮过程中乳酸发酵,快速产生乳酸、降低pH、氨态氮含量,提高了发酵品质、体外消化率和体外产气量。但纤维素酶和木聚糖酶组合添加效果最好。
猜你喜欢
林产化学与工业(2022年4期)2022-09-13 09:47:12
现代畜牧科技(2021年8期)2021-10-13 07:21:44
湖南饲料(2019年5期)2019-10-15 08:59:12
湖南饲料(2019年5期)2019-10-15 08:59:10
猪业科学(2018年5期)2018-07-17 05:55:56
猪业科学(2018年4期)2018-05-19 02:04:36
广东饲料(2016年2期)2016-12-01 03:43:05
中国酿造(2016年12期)2016-03-01 03:08:21
化工进展(2015年6期)2015-11-13 00:27:42
中国医科大学学报(2015年10期)2015-03-01 02:09:50
